土壤纤毛虫群落对退牧还草生态恢复的响应
——以玛曲县为例
2020-06-03宁应之周小燕烟郑杰张世虎陈凌云万贯红
宁应之,周小燕,烟郑杰,张世虎,陈凌云,万贯红
西北师范大学生命科学学院,兰州 730070
20世纪中后期,我国大多数草原草场特别是西部草原由于生态环境的脆弱,自然气候因素的影响,外加人类活动的干扰(如开垦草原、超载放牧、破坏草原植被等),草地退化问题严峻[1]。特别是针对西部地区草原生态环境持续恶化的严峻现状,全国草原荒漠化趋势不断扩大,为了遏制和扭转这种日益严重的草场退化,国务院于2002年12月正式批准在西部11个省份实施退牧还草政策,明确退牧还草的总体思路、规划总体目标和政策要点,使实现草地生态环境良性循环[2]。退牧还草工程涉及草地生态修复和牧区建设,坚持保护为先,以建设和合理利用相结合,实行以草定畜,严格控制载畜量;实行草场围栏封育,开展禁牧、休牧和划区轮牧措施;适当建设人工草地和饲草基地,旨在保护草原生态环境、遏制天然草地的持续恶化,实现畜牧业可持续发展。
纤毛虫原生动物作为一种复杂而高等的单细胞真核生物,在土壤生态系统和食物网中占有十分重要的地位。因其种类繁多、繁殖率高、个体微小、现存量和生产量巨大、群落演替迅速、分布广泛、比表面积大,从而对生态环境的变化十分敏感。在不同土地管理方式导致的土壤微环境变化中,其纤毛虫群落结构及多样性发生明显改变,因此在环境监测领域中可作为理想的指示生物[3- 4]。目前,土壤纤毛虫已被广泛应用于农田[5]、森林生态系统[6]的评价与重金属毒性[7]监测中并成为当前环境质量评价必不可少的重要组成部分,针对农田或退牧还草后土壤纤毛虫群落结构变化的研究则相对较少,目前仅见宁应之等对甘肃省定西市、天水市、庆阳市等地关于土壤纤毛虫对退耕还林生态恢复响应[8- 10]的相关报道。甘肃作为首批天然草地退牧还草工程实施省份,甘南玛曲草地是亚洲植被最好的草地之一,是甘肃天然草场载畜量最高、耐牧性较高的牧区。近年来在人为的干预下草地质量退化较为突出,相应出现了“私地悲剧”等现象。玛曲县退牧还草工程于2004年全面实施,包括尼玛、曼日玛、阿万仓、齐哈玛和欧拉5乡[11],其中欧拉乡比国家提出退牧还草政策要早,在当地政府引导下牧民于1995年对退化严重的地区实行禁牧,中度和轻度退化区实行休牧,植被较好的草原实行划区轮牧。本文以甘肃省甘南藏族自治州玛曲县境内欧拉乡的退牧还草地为样点,探讨不同恢复年限的退牧还草区域中土壤纤毛虫群落结构变化,分析研究表层土壤纤毛虫群落物种数和密度与土壤环境因子间的相关关系,并利用土壤纤毛虫物种数和密度对退牧还草生态恢复的效果进行评价,旨在为该地区退牧还草效果的生物学评价体系建立及退牧还草工作的实践提供基础资料和理论依据。
1 材料与方法
1.1 研究区概况
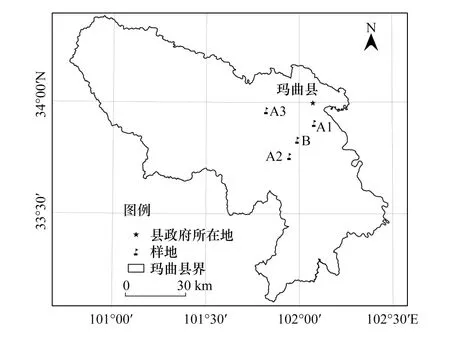
图1 研究区地理位置Fig.1 Location of study area A1:退牧还草10年;A2:退牧还草15年;A3:退牧还草20年;B:未进行退牧还草的对照样点
研究区位于甘肃省甘南藏族自治州玛曲县境内(图1),海拔3300—4806 m,整个地势呈东高西低的走向。气候属高原大陆性气候—高寒湿润区,高寒多风雨雪。年平均温度1.7℃,冬季最低温度达-20.9℃,夏季最高温度20.7℃,年均降水量599.7 mm。海拔高、辐射量大、温度低、牧草生长期短,年均日照时数为2594 h。草地类型以高寒草甸和沼泽类草甸为主。土壤类型以高寒草甸土和沼泽化草甸土为主,植被优势种为禾本科(垂穗披碱草Elymusnutans)、毛茛科(毛茛Ranunculusjaponicus,丝叶毛茛Ranunculustanguticus,云生毛茛Ranunculuslongicaulis,丝叶唐松草Thalictrumfoeniculaceum),蔷薇科(莓叶委陵菜Potentillafragarioides,蕨麻Potentillaanserina)和菊科(细叶亚菊Ajaniatenuifolia,冷蒿Artemisiafrigida)等为主,境内分布着黄河上游支流330余条,补给黄河径流水量的45%,是黄河源区主要水量补给区。
1.2 样品采集和处理
根据退牧还草年限的不同(图1),在玛曲县欧拉乡共设置4个采样点,分春夏秋冬四季采集土样。其中,A1、A2和A3样点在分别为2005、2000和1995年实施退牧还草工程(主要为当地政府指导下牧民自发开展的划区轮牧)。截至取样时,分别持续10、15、20年;B样点为对照样点未进行退牧还草工程(正常放牧、未采取休牧或轮牧措施)。
采样分别于2015年5月(夏)、8月(秋)、11月(冬)、2016年3月(春)进行。在各样点内随机选取5个25 m2(5 m×5 m)样区,每个样区用土壤采集器以“梅花五点式”采样法采集0—5 cm土层土样,拣去覆盖在土层表面的枯枝落叶凋落物后混合为1个样品,每次每个样点采集土壤样品5份;同时用土壤环刀在每个样点同样以“梅花五点式”采样法取0—5 cm土层土壤样品5个,封闭装好并做好标记,同时记录样地经纬度、地温。随后,将采集好的土样进行密封后带回实验室,一部分测定其含水量(24 h完成)与孔隙度;另一部分土样平整的在牛皮纸上铺开,上面覆盖两层草纸(防止空气中的包囊进入土样影响实验结果)于室内自然风干,用于土壤纤毛虫的定性和定量实验。在植物生长旺季即8月份对各样点土壤采样区以0.25 m2(0.5 m×0.5 m)的样方对地上植物种类、数量、盖度进行调查统计,每个样点5个样方。
1.3 项目测定与方法
1.3.1土壤理化因子的测定
用曲管地温计测定土壤温度,利用复合式电极进行土壤pH测定,烘干法测定土壤含水量,凯氏定氮法测定总氮,氢氧化钠碱熔-钼锑抗比色法测定总磷和总钾,碳酸氢钠浸提-钼锑抗比色法测定速效磷,重铬酸钾容量法测定有机碳,火焰光度计测定速效钾。
1.3.2纤毛虫的研究方法
定性研究采用“非淹没培养皿法”[12]即在直径为15 cm的培养皿中加入50 g的土壤,25℃泰斯特(GZX- 250E)下培养,第三天开始镜检(Nikon光学显微镜)鉴定物种,每份土样重复培养若干次、多次镜检,直到不再有新物种的出现为止。鉴定技术包括活体观察(Nikon光学显微镜)和Wilbert(1975)蛋白银染色[13]。采用Lynn(2008)分类系统[14]进行物种鉴定。
定量采用直接计数法,即在直径为5 cm的培养皿中加入5 g土壤,添加蒸馏水至刚刚没过土壤并记录添水量,培养皿壁的水平面划线标记,进行培养,每天观察蒸发水量并重新添加水量至刻度线,在培养的第8、9、10、11四天内对其中的纤毛虫数量进行统计,再根据添水量和土壤质量对土壤中的纤毛虫进行丰度换算。
1.4 数据统计处理与方法分析
1.4.1类群的划分
对鉴定到的物种进行统计分析,将物种数最多的目定义为优势类群,次多的目定义为次优势类群,只有单一种的目规定为罕见类群,其余目均为常见类群[15- 16]。
1.4.2物种多样性指数
多样性指数是用简单的数值表示生境中动植物以及微生物种类的丰富程度,用来判断群落或者生态系统是否稳定,是衡量一定地区生物资源丰富程度的一个客观指标。本文采用Shannon-Wiener多样性指数[17]来计算物种多样性:
H=-∑Ni/N×lnNi/N
式中,H为多样性指数,N为所有属的个体数,Ni是第i属的个体数。H的范围在0到任何整数之间,当H=0时表示全部个体均属于同一属生物,H值越大,表示当前物种多样性指数越高。一般来说,当H值在0—1之间时污染程度较重,H>3时当前污染程度较轻[18]。
1.4.3丰富度指数
丰富度指在某一特定区域内所有物种的数量,用来整体衡量当前区域内物种资源的丰富程度,表征生物群集中种类的丰富程度。本文采用Margalef丰富度指数[19]来计算物种丰富度:
d=(S-1)/lnN
式中,d为丰富度指数,S为种类数,N为个体数,d值越高则当前污染程度较轻,反之则较重。一般来说,当d>6时表明当前环境下生物丰富度较高[20]。
1.4.4均匀度指数
均匀度指一个群落或者生境中不同群落的分布情况,其反应出物种个体数目之间分配的均匀程度。在衡量一个区域内的群落多样性时,必须同时考虑群落丰富度及均匀度,这样计算出来的结果才具有统计学意义。本文采用Pielou均匀度指数[21]计算群落均匀度:
E=H/lnS
式中,E为均匀度指数,H为Shannon-Wiener多样性指数,S为种类数。一般情况下E的值在0—1之间,E值越高,则表明当前生态环境不同群落分布越均匀,种类越丰富。
使用SPSS 19.0和Primer 6.0对数据统计、聚类分析(Cluster Analysis)和多维排序分析(Multidimensional Scaling,MDS);单因素方差分析用于各样点间土壤纤毛虫群落组成随季节变化(P<0.05)、用Origin 8.0做图。
2 结果
2.1 土壤理化因子对退牧还草的响应
土壤含水量和孔隙度是表征土壤结构以及其水源涵养能力的物理环境指标(表1)。土壤含水量和孔隙度A1、A2、A3样点均高于B样点,且A3>A2>A1;即表明随着退牧还草工程年限的增长,其土壤物理状况得到显著改善;4个样点之间的土壤pH值与退牧年限成反比,随着退牧年限的增长土壤由弱碱性向中性过渡差异显著;土壤速效钾和速效氮含量在恢复样地A1、A2、A3样点均显著高于对照样点B且A3样点与B样点均差异显著(P<0.05),即随着退牧还草年限增加,土壤速效养分相应增加;土壤全钾、有机质含量在样点各样点间差异不显著,可见随退牧还草工程的实施和年限的增加,对土壤理化性质及养分的改变较为明显。
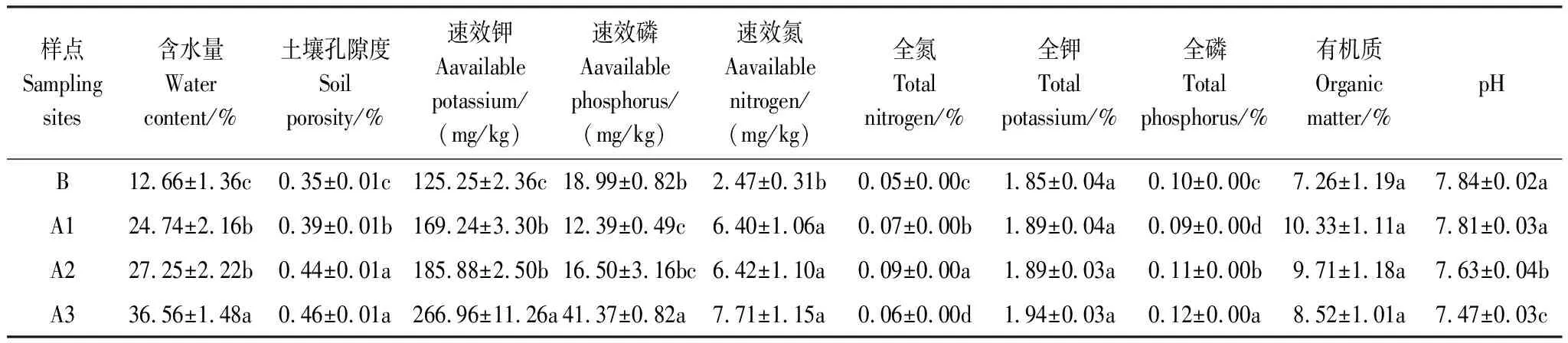
表1 各样点土壤理化因子
A1:退牧还草10年;A2:退牧还草15年;A3:退牧还草20年;B:未进行退牧还草的对照样点; 表中所列数据为平均值±标准误(SE),同一列不同字母代表经LSD多重比较在P<0.05水平样点间差异显著(n=20)
对所研究样点的土壤理化指标(含水量、土壤孔隙度、pH值、速效氮、速效磷、速效钾、有机质、全氮、全磷、全钾)进行聚类分析(图2)可得,未进行退牧还草工程的对照样点B首先被单独分出来,而经过10年和15年退牧还草工程的A1样点和A2样点聚为一类,接着与退牧还草20年的A3样点聚为一类,随后3个样点与B聚为一类,这表明退牧还草后的土壤质量与对照样点B产生了明显差异,退牧还草20年的A3样点的草原生境得到了明显的改善。
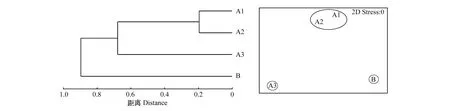
图2 各样点环境因子聚类分析树状图和多维度排序图Fig.2 Dendrogram of cluster analysis and Multi dimensional scaling on soil environment factorsat various sites
2.2 纤毛虫的群落结构
2.2.1纤毛虫的群落组成
本研究中共计鉴定到纤毛虫95种,隶属9纲15目21科28属(表2)。在群落组成上,对照样点B的群落结构较为简单,A1—A3样点与对照样点B有较大的差别,A1—A3样点的目、科、属和种的数目呈明显的增加趋势在A3样点高达85种,表明随着退牧还草工程的实施和退牧还草年限的增加,土壤逐渐改善,越来越适合土壤纤毛虫的存在,纤毛虫群落结构逐渐复杂化,其中散毛目(Sporadotrichida)居多,为优势类群,肾形目(Colpodida)、尾柱目(Urostylida)、刺钩目(Haptorida)次之,为次优势类群。
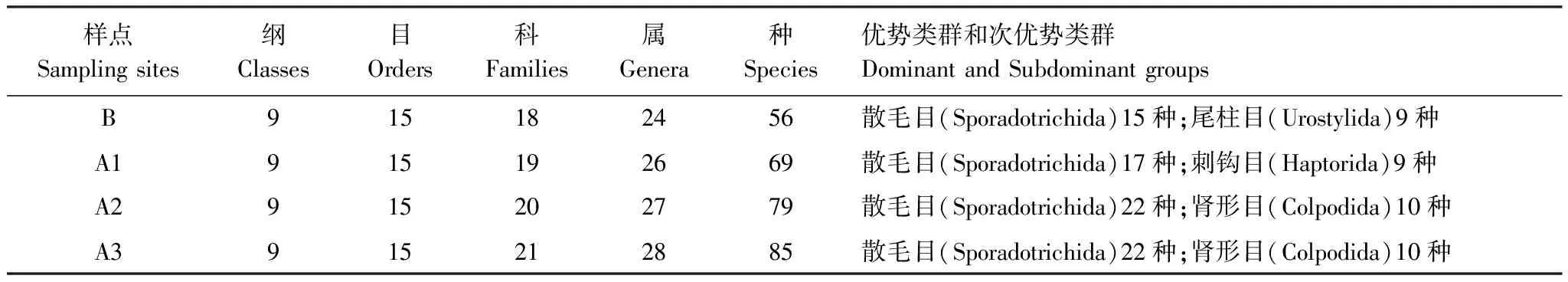
表2 各样点土壤纤毛虫的群落组成
2.2.2优势种
本研究将各样地不同季节中出现频次相对较高的纤毛虫物种作为优势种。其优势种的响应情况如下:尖毛虫属(Oxytricha)中的Oxytrichasp1.、颗粒尖毛虫(Oxytrichagranulifera)、Oxytrichasp2.,肾形虫属(Colpoda)中的土壤肾形虫(Colpodaedaphoni)、小肾形虫(Colpodaminima),弹跳虫属(Halteria)中的大弹跳虫(Halteriagrandinella)在大部分的处理中成为优势种。
从各样点的优势种来看,对照样点B中的优势种为大弹跳虫(H.grandinella)、土壤肾形虫(C.edaphoni)、膨大肾形虫(Colpodainflata),A1样点中的优势种尖毛虫属一种(Oxytrichasp1.)、大弹跳虫(H.grandinella)、土壤肾形虫(C.edaphoni)、腔裸口虫(Holophryaatra),A2样点中的优势种为尖毛虫属一种(Oxytrichasp2.)、大弹跳虫(H.grandinella)、土壤肾形虫(C.edaphoni)、小肾形虫(C.minima),A3样点中的优势种为苔藓斯道克虫(Sterkiellahistriomuscorum)、颗粒尖毛虫(O.granulifera)、契氏片尾虫(Urosomacienkowskii)、大弹跳虫(H.grandinella)。表明经过退牧还草生态恢复后的草场土壤纤毛虫优势种存在显著差异,随着退牧还草工程时间的推进,土壤纤毛虫优势种增加并且趋于复杂化,并且物种由对照样点B的肾形目(r-对策者)逐渐演替为以散毛目(k-对策者)等为主的土壤纤毛虫过渡,这也证明随着退牧还草工程的不断实施,尤其是随着退牧还草年限的增长,土壤环境条件跟质量逐步得到改善和提高。
2.2.3广布种和特有种
本研究中有32种纤毛虫在4个样点均出现,为广布种,占纤毛虫物种总数的33.68%,分别为透明赭虫(Blepharismahyalinum)、苔藓圆纤虫(Strongylidiummuscorum)、尖毛虫属一种(Oxytrichasp1.)、颗粒尖毛虫(O.granulifera)、椭圆尖毛虫(Oxytrichaelliptica)、尖毛虫属一种(Oxytrichasp2.)、似片尾虫属一种(Urosomoidasp1.)、契氏片尾虫(U.cienkowskii)、阔柱片尾虫(Urosomamacrostyla)、相似片尾虫(Urosomasimillis)、近缘殖口虫(Gonostomumaffine)、钢强殖口虫(Gonostomumstrenuum)、纺锤全列虫(Holostichakessleri)、大尾柱虫(Urostylagrandis)、尾瘦尾虫(Uroleptuscaudatus)、瘦尾虫属一种(Uroleptussp.)、大弹跳虫(H.grandinella)、驼扭头虫(Metopusgibbus)、斜口虫属一种(Enchelyssp.)、卑怯管叶虫(Trachelophyllumpusillum)、俏篮口虫(Nassulagracilis)、小肾形虫(C.minima)、僧帽肾形虫(Colpodacucullus)、土壤肾形虫(C.edaphoni)、膨大肾形虫(C.inflata)、迅捷肾形虫(Colpodafastigata)、背沟肾形虫(Colpodahenneguyi)、肾形虫属一种(Colpodasp1.)、长篮环虫(Cyrtolophosiselongate)、腔裸口虫(H.atra)、沟裸口虫(Holophryasulcata)、肾形瞬目虫(Glaucomareniformis)。
仅在某一样点出现的种为特有种,A1样点中的特有种有2种,分别为殖口虫属一种(Gonostomumsp.)、漫游虫属一种(Litonotussp.),A3样点中的特有种有1种,为角毛虫属一种(Keronopsissp.)。
2.2.4纤毛虫的物种分布
本研究中所鉴定到的纤毛虫95种的分布如图3所示,退牧还草生态恢复20年的A3样点物种最为丰富,有85种,约占物种总数的89%,A2样点和A1样点分别有79种和69种,分别占物种总数的83%和72%;对照样点B物种数为56种,占物种总数的59%,可见随着退牧还草工程的开展和年限的延长,土壤纤毛虫物种数呈明显增多趋势。
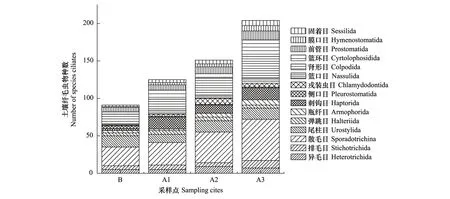
图3 各样点土壤纤毛虫群落组成Fig.3 The community composition of the soil ciliates at various sampling sites
2.2.5土壤纤毛虫的物种密度、多样性、丰富度、均匀度指数
不同的生态恢复年限纤毛虫密度和各物种多样性指数有所不同(图4),纤毛虫密度在各处理间均随退牧还草年限增加而升高。春夏秋冬四季纤毛虫密度在退牧还草样点A均高于对照样点B,即A3(春:220 个/g;夏:201 个/g;秋:204 个/g;冬:158 个/g)显著高于A2(春:138 个/g;夏:132 个/g;秋:168 个/g;冬:106 个/g)、A1(春:145 个/g;夏:115 个/g;秋:143 个/g;冬:75 个/g)和B(春:56 个/g;夏:63 个/g;秋:83 个/g;冬:42 个/g),季节间为秋季>春季>夏季>冬季,表明退牧工程实施有助于纤毛虫物种数增加,从而使得土壤中纤毛虫密度的提高。
Shannon-Wiener多样性指数的各处理变化与Pielou均匀度指数的变化规律基本一致(图4),随着退牧还草年限的延续,在季节上A3物种多样性指数(春:3.14;夏:3.58;秋:3.52;冬:2.34)显著高于A2(春:2.75;夏:2.90;秋:2.78;冬:1.98)、A1(春:2.42;夏:2.69;秋:2.53;冬:1.82)和B(春:1.90;夏:2.10;秋:2.12;冬:1.48);A3均匀度指数(春:1.05;夏:1.20;秋:1.17;冬:0.83)显著高于A2(春:0.95;夏:0.98;秋:0.93;冬:0.70)、A1(春:0.81;夏:0.90;秋:0.85;冬:0.64)和B(春:0.74;夏:0.71;秋:0.72;冬:0.54),两者都呈明显的夏季>秋季>春季>冬季;Margalef丰富度指数各处理样点的变化除A2与A1差异不显著外(P>0.05)其他样点间随季节均表现差异显著,丰富度指数各样点间也是随着退牧年限增加逐渐升高,A3(春:4.89;夏:4.93;秋:4.91;冬:4.96)显著高于A2(春:4.54;夏:4.58;秋:4.48;冬:4.30)、A1(春:4.21;夏:4.15;秋:4.20;冬:3.91)和B(春:3.37;夏:3.50;秋:3.60;冬:3.00),整体呈秋季>夏季>春季>冬季。
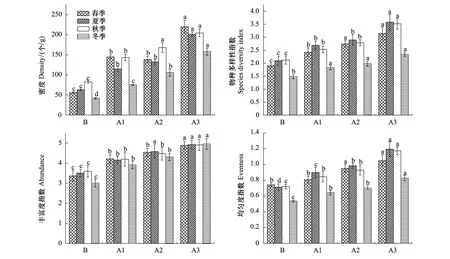
图4 各采样点的纤毛虫密度、物种多样性指数、丰富度指数和均匀度指数Fig.4 Density, diversity index, abundance and evenness of soil ciliates at various sampling sites*不同字母为随各季节在不同样点间在0.05水平上的差异显著性
对土壤纤毛虫密度、多样性指数、丰富度指数及均匀度指数季节间采用无重复双因子方差分析显示:土壤纤毛虫密度、丰富度指数、多样性指数和均匀度指数各样点处理之间差异显著,季节间差异不显著;可看出经过退牧还草工程后的样点随年限的增加土壤纤毛虫密度、多样性指数、丰富度指数与均匀度指数均得到了不同程度的提高,当退牧年限不断增加时,土壤纤毛虫各群落结构参数持续增加,表明土壤纤毛虫群落结构向稳定过度,并且物种趋于复杂化。
2.3 土壤纤毛虫群落特征参数与土壤理化性质的相关性
对各样点土壤理化因子(速效氮、速效磷、速效钾、总氮、总磷、总钾、有机质、pH值、含水量、孔隙度)与纤毛虫群落特征参数(土壤纤毛虫密度、物种多样性指数、物种丰富度指数、物种均匀度指数)进行相关性分析(表3)。土壤纤毛虫密度与土壤速效氮、速效磷、速效钾、总氮、总磷、总钾、有机质和含水量均呈极显著的正相关关系(P<0.01),与土壤孔隙度呈显著的正相关关系(P<0.05),与土壤pH值呈极显著的负相关关系(P<0.01);土壤纤毛虫多样性指数和均匀度指数与土壤总氮、总磷、总钾、有机质、含水量、土壤孔隙度呈极显著的正相关关系(P<0.01);土壤纤毛虫丰富度指数与土壤速效磷、速效钾、总磷、总钾、有机质含量、土壤含水量和土壤孔隙度呈极显著的正相关关系(P<0.01),与土壤总氮呈显著的正相关关系(P<0.05),与土壤pH值呈显著的负相关关系(P<0.05),进一步表明随着退牧年限的增加土壤质量得到恢复为更多土壤纤毛虫的生长创造有利条件。
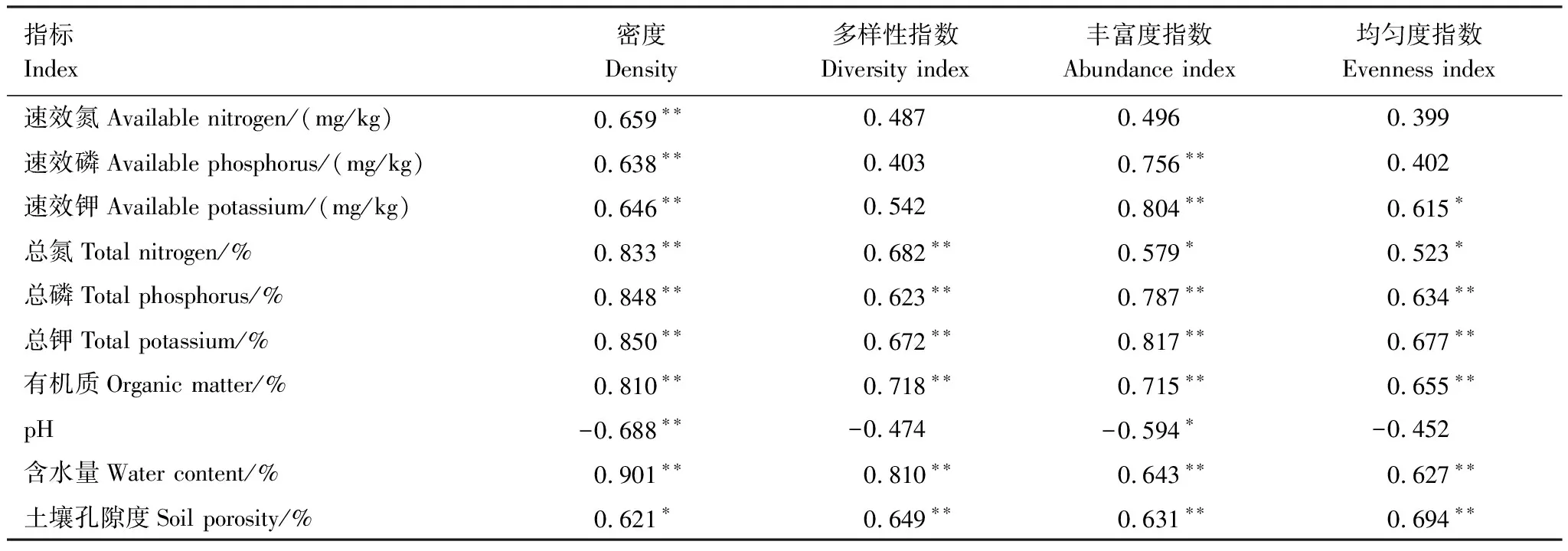
表3 土壤纤毛虫群落特征参数与土壤环境因子的相关性分析
*在0.05水平(双侧)显著相关,**在0.01水平显著相关
3 讨论
退牧还草政策旨在给予农牧民一定经济补偿的前提下,通过围栏建设、补播改良以及禁牧、休牧、划区轮牧等措施,恢复草原植被,提高草原生产力,促进草原生态与畜牧业协调发展。Peng等[22]在研究中发现食草性动物对草场的任意践踏可导致土壤容重、土壤pH等的增加,本研究中主要是采用休牧和划区轮牧两种方式,因为草地质量退化不是非常严重,在退牧还草工程实施后,草场植被在萌发期和结实期采取休牧其物种数量有了显著提高[11,23];章家恩等[24]认为在退化草地植被恢复和重建的过程中,物种多样性也会随即增加,植物多样性和盖度的提高会直接影响土壤养分的分布和提高[23,25]。本研究表明,经过退牧还草20年的A3样点地上生物量和盖度均最大,其土壤孔隙度、含水量、pH、钾、磷、氮以及有机质的含量较对照样点均有显著提高,这与曾辉等[25]和仝小林等[26]结果相对应,说明退牧还草生态恢复后,草地得到休养生息,地下土壤养分状况得到相应改善,而且退牧还草年限越长,地上植被得到恢复和再生,当年返回土壤中的枯落物使得土壤有机碳、氮、磷和钾得到补充和积累,使得土壤通气性得到改善,便于水汽交换和养分的保持,良好土壤通气性和肥沃的有机质将为土壤微生物的繁殖和活动提供良好的条件[27]。
至于退牧还草年限最终持续多久,土壤质量状况和土壤原生动物—纤毛虫群落组成、结构、数量和功能才能趋于稳定,仍需后续研究。
随着退牧还草生态恢复的持续,植被得到了恢复和重建,植被凋落物促进土壤动物活力和微生物的活性加强[28-29],土壤原生动物吸取排泄等改善土壤质地刺激植物生长[30],纤毛虫作为最复杂和最高等的单细胞原生动物,是土壤生态系统中十分重要的动物类群之一,因其独特的生命特征,对外界环境的变化十分敏感,因而可通过考察其群落的各项特征参数及其动态来反映生态环境的变化[31]。
本研究中共鉴定到纤毛虫95种,隶属9纲15目21科28属,这与孙辉荣等[30]和刘灿等[31]对亚高寒草甸冷暖季土壤纤毛虫群落对坡向的响应结果较低,本研究中纤毛虫物种数与亚高寒草甸冷季纤毛虫物种数量为97种(9纲、17目、32科、50属)基本相等,但明显低于亚高寒草甸暖季纤毛虫物种数142(9纲、18目、32科、55属)和宁应之等[32- 33]玛曲高原湿地土壤纤毛虫类群春季(32科50属114种)和夏季(40科68属204种),主要归因于本研究区的地形海拔较高导致气温在本研究中更低所致以及沼泽湿地水热等优越的土壤环境有助于微生物的生存[30- 33]。且随着草地恢复年限的增加其土壤纤毛虫物种数增加变幅相应减小,且优势类群跟次优势类群在A2和A3间基本相似。随着退牧还草工程的不断推进,土壤纤毛虫群落结构趋于复杂化、物种数逐渐增加,以r-对策者肾形目为优势种逐渐向k-对策者散毛目过渡,Foissner[28]在研究中发现,散毛目(Sporadotrichida)和肾形目(Colpodida)的种类常占优势,其中又以散毛目(Sporadotrichida)的种类最为丰富[34- 35],这与本研究的结果相一致。
土壤纤毛虫密度、多样性指数、丰富度指数和均匀度指数都随着退牧还草进行均对生态恢复方面做出了积极响应。主要体现在,随着退牧还草年限的延长,纤毛虫群落组成逐渐复杂化,群落结构的稳定性持续提高,如纤毛虫物种数和特有种呈增多趋势、丰富度波动式上升、多样性指数逐渐升高及优势类群和优势种发生良性演替。方差分析显示尽管土壤纤毛虫多样性、丰富度和均匀度指数等群落特征在季节上差异不显著,但群落特征指数变化在夏季跟秋季间表现活跃,冬春季表现更为稳定,这主要受土壤环境的变化随高原气候低温高湿的限制更为明显,因为海拔限制主要是温度跟水分的权衡,研究表明土壤湿度对土壤纤毛虫物种数影响最大[32- 35],本研究中土壤含水量与纤毛虫密度、多样性指数、丰富度指数呈极显著正相关且相关性系数均较高相一致。
研究区属于典型的高寒草甸气候、温度、降雨等环境条件变化强烈,土壤植被等空间异质性很大。退牧还草生态恢复后,一般为植物盖度、地上生物量相应增加[23,35],伴随土壤孔隙度、含水量、总磷、总氮、总钾、速效氮、速效磷、速效钾随生态恢复年限的增长而显著增加,土壤pH显著降低,土壤理化性质的改变导致了土壤纤毛虫群落各项特征的相应变化。相关性分析表明,土壤纤毛虫群落主要特征参数(密度、均匀度指数、丰富度指数和多样性指数)随恢复年限及土壤主要环境理化因子(速效氮、速效磷、速效钾、总氮、总磷、总钾、有机质、含水量、土壤孔隙度)之间存在显著或极显著的正相关关系与pH呈极显著负相关关系,各环境因子中对纤毛虫群落影响较大的是氮、钾、磷和有机质含量,这与宁应之等[8-10]的研究结论相符。研究发现[36- 37],土壤纤毛虫是生态恢复效果的良好指示生物,说明土壤基质即土壤腐殖质等养分含量高低是决定土壤微生物包括原生动物—纤毛虫定居的前提,而土壤基质和植被等的优劣又会受到地形气候等水热条件的反馈和调节。
由此可见,土壤纤毛虫群落结构显著地响应了退牧还草生态恢复所导致的土壤生态环境效应,土壤纤毛虫群落的各项特征参数可作为退牧还草生态恢复效果的评价指标之一,这将为未来退化高寒草甸的植被恢复和土壤碳汇保持具有重要的指导意义。
