基于生长与结实评价红松种子园亲本
2020-06-03周雪燕高海燕李召珉赵银琨葛丽丽侯庆文丁文雅赵曦阳
周雪燕 高海燕 李召珉 赵银琨 葛丽丽 侯庆文 丁文雅 赵曦阳*
(1.东北林业大学林木遗传育种国家重点实验室,哈尔滨 150040; 2.吉林省临江林业局,临江 134600)
林木种子园是为获得优质种子,经过人为设计的特种人工林,在实现林木良种化过程中发挥着重要的作用[1],具有提早开花结实,实现树体矮化,便于采收,方便管理等优点。林木种子园的营建历史可以追溯到1880年,荷兰人为了提高金鸡纳树(Cinchonaledgeriana)的奎宁含量,在爪哇营建了金鸡纳无性系种子园[2]。20世纪30年代,许多欧美国家进行了多种针叶树种种子园的营建[3]。20世纪60年代种子园风靡世界,各国以种子园为主要环节的针叶树遗传改良工作取得较大进展。我国于1964年开始建立杉木无性系种子园,20世纪70年代林木种子园有了较大发展,特别是从20世纪80年代初开始,种子园营建技术列为国家重点科技攻关项目,受到林业生产部门的重视,取得了一定的研究成果。但是与欧美国家相比,我国种子园的发展相对落后,目前我国大部分树种种子园还停留在初级种子园阶段,造成我国良种化水平与林业发达国家相差甚远。
红松(Pinuskoraiensis)是松科(Pinaceae)松属(Pinus)的常绿高大乔木[4],属北半球中高纬度地区分布面积较大且闻名世界的三大五针松之一,其主要分布在亚洲东北部日本海西岸从朝鲜半岛经中国东北到俄罗斯远东南部的三角地带[5],在我国主要分布于长白山及其北部的张广才岭、老爷岭、完达山和小兴安岭[6]。红松不仅树冠高大,树干通直,材性优良,易于加工[7],而且果实美味,富含不饱和脂肪酸和酚类等活性物质,具有较高的营养价值和医疗保健价值[8~12],是我国东北地区重要的果材兼用树种。我国于20世纪80年代开始营建红松种子园[13~14],由于红松生长缓慢,最初建立的种子园目前仍处于初级种子园阶段,只有部分种子园营建了子代测定林,并开展了生长性状的测定研究[15],但研究结果未能有效指导种子园的升级换代,且红松存在无性系间花期不遇、周期性结实、产籽量低等问题,极大影响红松遗传改良进程[16]。
本研究以吉林省临江林业局闹枝林场红松子代测定林的69个半同胞家系为材料,测定其26和28年生的生长性状和结实性状,通过遗传变异参数的计算,初步筛选生长与结实性状优良的亲本无性系与家系,选出的家系可以进行区域化试验,为生产应用提供物质基础,初选的优良亲本无性系可为改良种子园和1.5代种子园的营、改建提供材料,本研究可为其他树种遗传改良提供参考。
1 试验材料与方法
1.1 试验地点
试验林营建于吉林省临江种苗示范中心国家红松、水曲柳良种基地(47°37′N,126°54′E),该地属温带季风气候,海拔510 m,年均气温5℃,年均降水744 mm,无霜期128 d,土壤类型为暗棕土壤。
1.2 试验材料
本研究中试验材料包括67个红松半同胞家系与2个对照家系(见表1),于1987年采种,次年播种育苗,1992年利用4年生实生苗营建子代测定林。试验设计采用完全随机区组设计,6株小区,5次重复,株行距2 m×2 m,外加两层保护行。

表1 家系来源表
1.3 试验方法
2013年11月左右测定69个家系的树高、胸径和单株球果数,2015年11月左右测定树高、胸径、单株球果数、鲜塔重、千粒重及单塔出籽量。树高、胸径和单株球果数进行全林调查;各家系随机采集30个鲜塔测定其鲜塔重和单塔出籽量;各家系随机选取4 000粒种子分为4份,测定千粒重。
1.4 数据分析方法
利用Excel 2010和SPSS 19.0软件对所测得数据进行分析[17]。
树高、胸径和单株球果数方差分析线性模型采用公式[18]:
Xijk=μ+Fi+Bj+FBij+eijk
(1)
式中:μ为总体平均值,Fi为家系效应,Bj为区组效应,FBij为家系和区组的交互作用,eijk为随机误差。
鲜塔重、千粒重和单塔出籽量方差分析线性模型采用公式[19]:
Xij=μ+Fi+eij
(2)
式中:μ为总体平均值,Fi为家系效应,eij为随机误差。
家系遗传力(h2)采用公式[20]:
(3)
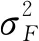
表型变异系数(PCV)采用公式[21]:

(4)
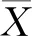
表型相关系数(r)采用公式[22]:
(5)

一般配合力(GCA)采用公式[23~24]:
GCA=Xi-X..
(6)
式中:Xi为某一性状某个家系平均值,X..为该性状总体平均值。
利用多性状综合评价法对家系进行综合评定,采用公式[25]:
(7)
式中:其中ai=Xij/Xjmax,Qi为各家系的综合评价值,Xij为某一性状的平均值,Xjmax为该性状的最大值,n是该性状的个数。
遗传增益估算采用公式[18]:
(8)
2 结果分析
2.1 红松半同胞家系生长与结实性状的方差分析
69个红松半同胞家系各指标方差分析见表2,结果表明除单株球果数在家系与区组交互作用时差异不显著外,其他各测定指标在不同树龄时各变异来源间均达极显著差异(P<0.01)。
2.2 家系间各指标遗传变异参数
69个红松半同胞家系各指标遗传变异参数见表3,所有家系除千粒重外,其他指标最大值均在最小值1.50倍以上,其中树高、鲜塔重和单塔出籽量在1.50~2.00倍,胸径和单株球果数在3.00倍以上,而单株球果数的变化幅度较大,最大达到32.67,最小为0.33。各指标表型变异系数变化范围为6.30%~63.00%,其中26和28 a单株球果数的表型变异系数均超过50%,树高、胸径和鲜塔重的表型变异系数在10%~40%,千粒重和单塔出籽量的表型变异系数均小于10%。各指标的遗传力均较高,大部分在0.90以上,除了单塔出籽量的遗传力为0.87。
2.3 红松半同胞家系各指标的一般配合力
69个红松半同胞家系各指标一般配合力见表4。从26 a分析结果来看,单株球果数的一般配合力范围为-12.29~11.71;从28 a分析结果看,树高、胸径、单株球果数、鲜塔重、千粒重和单塔出籽量的一般配合力范围分别为-1.77~1.69、9.10~8.65、-15.97~16.03、85.29~85.95、-89.25~80.75和-11.61~14.26。对于树高来说,P029、A101、P119、P081、P076、P113和P045家系的一般配合力较高(1.45~1.69),对于胸径来说,A096、P047、A113、P041、P113、P039和A030家系的一般配合力较高(7.01~8.65),其中P113家系的树高和胸径的一般配合力均较高;对于单株球果数来说,P004、P096、P039、P053、A124、A100、A109等家系的一般配合力较高(15.03~16.03),对于单塔出籽量来说,A124、A108、A048、P095、P078、P044和P045家系的一般配合力较高(9.16~14.26),综合来说A124家系的单株球果数和单塔出籽量的一般配合力均较高。
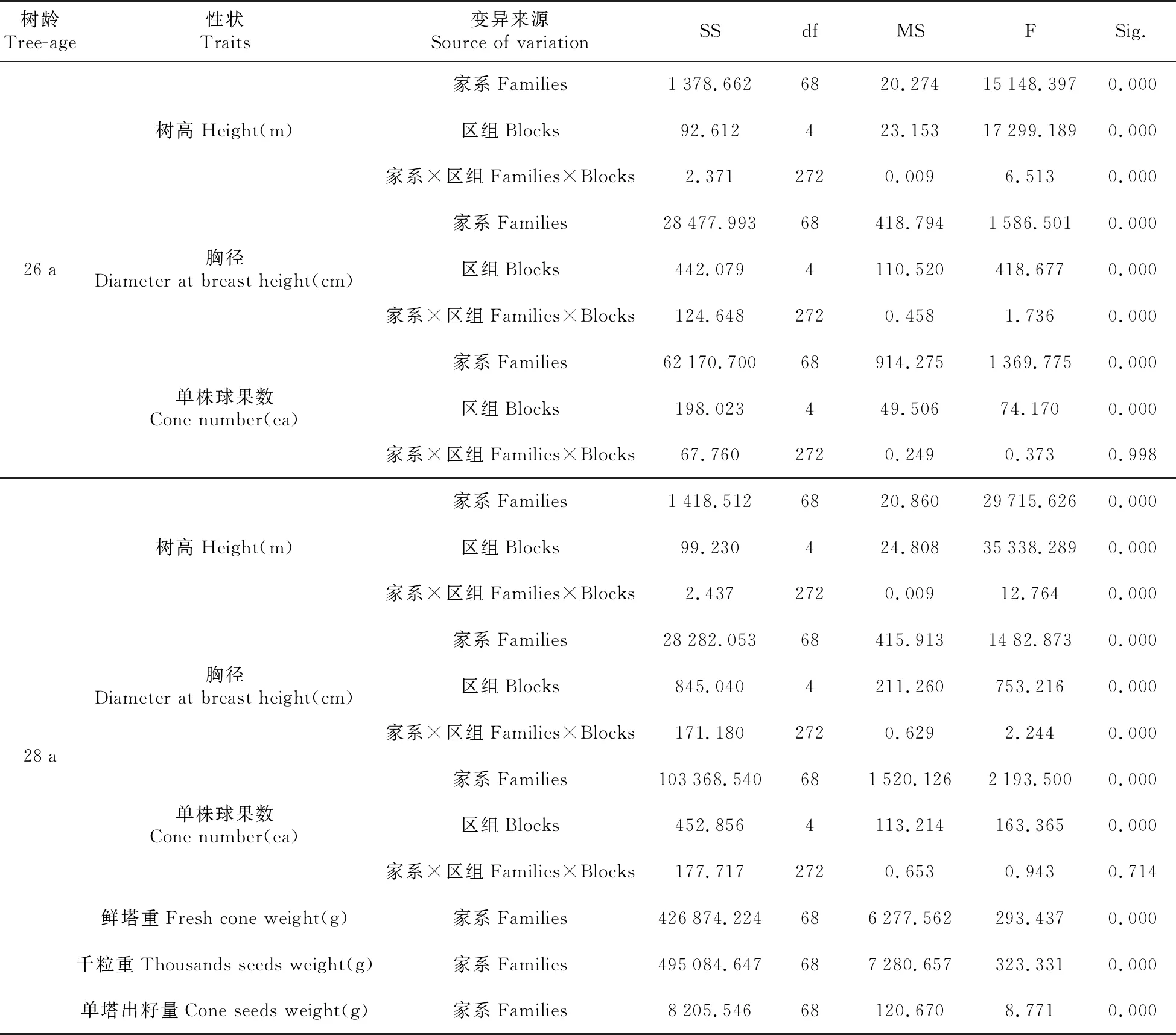
表2 69个红松半同胞家系各指标方差分析表

表5 69个红松半同胞家系各指标相关系数
注:*表示相关性达显著水平(P<0.05);**表示相关性达极显著水平(P<0.01)
Note:*correlation is significant at the 0.05 level;**correlation is significant at the 0.01 level

表6 多性状综合评价表
2.4 红松半同胞家系各指标相关性分析
69个红松半同胞家系各指标相关性系数见表5,从生长性状来看,26和28 a胸径与树高均达极显著正相关水平(0.823,0.720);28与26 a树高和胸径均达极显著正相关水平(0.995,0.998)。从结实性状来看,28与26 a单株球果数之间呈正相关,但相关性未达显著水平(0.045),说明红松的周期性结实问题;28 a鲜塔重与26和28 a单株球果数之间分别呈负相关和正相关,均未达显著水平(-0.131,0.035);28 a的单株球果数和鲜塔重与26和28 a树高和胸径均呈负相关,与树高之间相关性达极显著水平(-0.204,-0.202,-0.214,-0.189),与胸径之间相关性未达显著水平(-0.022,-0.051,-0.014,-0.038),说明树高越高胸径越大,单株球果数越少,鲜塔重越轻,但主要取决于树体的高低;千粒重与单株球果数之间呈极显著负相关(-0.230),与其他性状之间的相关性很弱;28 a单塔出籽量与26和28 a树高之间分别呈显著正相关(0.157,0.151),与26和28 a的单株球果数之间分别呈极显著负相关(-0.259)和未达显著水平的正相关关系(0.112),进一步说明了红松的周期性结实问题,与鲜塔重之间呈显著正相关(0.138),与胸径和千粒重之间存在很弱的正相关关系。
2.5 红松半同胞家系多性状综合评价
相关性分析表明生长性状和结实性状之间的相关性很弱,因此分别利用生长性状(树高和胸径)和结实性状(单株球果数、鲜塔重和单塔出籽量)为评价指标,以28 a红松数据为依据,对其半同胞家系进行综合评价(见表6所示)。以10%的入选率进行筛选,生长性状方面入选的家系有A096、P113、P047、A101、P041、P127和P107,入选家系树高和胸径的平均值分别为8.64 m和22.30 cm,遗传增益分别为15.66%和35.40%;结实性状方面入选的家系有A124、P096、A100、A123、P088、P105和P039,入选家系单株球果数、鲜塔重和单塔出籽量的平均值分别为30个、386.88和81.93 g,遗传增益分别为87.30%、1.77%和3.39%。
3 讨论
遗传、变异和选择是生物进化和新品种选育的三大因素,研究林木的遗传和变异是林木遗传改良的重要基础[26]。本研究通过对69个红松半同胞家系的26和28 a的生长性状和结实性状进行测定分析,方差分析结果表明除单株球果数在家系与区组交互作用时差异不显著外,其他各测定指标在不同树龄时各变异来源间均达极显著差异(P<0.01),说明不同树龄、不同家系间变异丰富,69个半同胞家系的生长性状和结实性状具有非常大的选择潜力,对优良家系的评价选择具有重要意义。
变异系数反映单位均值上的离散程度,变异系数越大,则性状离散程度越大[27],可以用表型变异系数的大小直接反映不同家系群体各性状的变异程度。本研究中,树高和单塔出籽量的表型变异系数略高于姜国云等[28]对红松家系的研究,说明树高和单塔出籽量存在更丰富的变异。遗传力是亲本将某一性状遗传给子代的能力,是选择育种中确定选种方法和估算遗传增益的重要参数[29],某一性状的遗传力越高,说明该性状可以稳定地遗传给子代,受环境因素的影响越小。本研究中,各指标的遗传力均较高,且除了28a的单塔出籽量的遗传力较低,其余均在0.90以上,明显高于宋云平等[30]的研究。高变异系数,高遗传力,更有利于家系的评价选择[31]。
在交配群体中,加性基因可以遗传给子代,而一般配合力恰好反映亲本加性基因的效应,一般配合力效应值的大小表明子代受亲本的影响程度,这是亲本选择的依据之一[32~33],因此种子园中优良亲本的选择主要是依据一般配合力的大小来确定。根据一般配合力的计算结果,本研究中分别选出7个树高、胸径、单株球果数和单塔出籽量一般配合力较高的亲本无性系。本研究中利用树高的一般配合力所选家系的平均值为1.58,利用胸径的一般配合力所选家系的平均值为7.89,都明显高于刘凤平等[31]对于树高和胸径一般配合力的研究结果,这主要由于树龄不同,环境不同所导致。针对不同树龄的树高和胸径选出的优良亲本基本相同,而利用不同树龄的结实量筛选的优良亲本差异较大,说明不同树龄结实不稳定,表明对于结实性状的评价选择应基于多年的观测结果[34~36]。
相关性反映了两个性状之间的相关程度,可以利用相关性确定与育种目标联系比较紧密的直观易测的性状用来挑选优良家系[37~38]。本研究中树高或胸径的不同测定年份之间具有极高的相关性,相关系数达0.99以上,与姜国云等[28]的研究结果相似,相同测定年份,树高与胸径的相关系数在0.720~0.823,高于王璧莹等[39]的研究结果。单株球果数与树高和胸径之间的相关性不一定,26 a单株球果数与树高和胸径之间相关性均不高;28 a单株球果数与26和28 a树高呈极显著负相关,与胸径之间相关性未达显著水平,这与王有菊等[40]、尚福强[41]关于红松生长与结实之间相关关系的研究不同,可能是由于本研究中试验林密度过大,且红松本身周期性结实所导致。焦宏等[42]的研究表明,红松的结实量与胸径相关性大于树高,但本研究中的结果正好相反,可能是由于本研究中的子代测定林密度较大,已经影响了红松的树高与胸径的生长。单塔出籽量与鲜塔重之间呈显著正相关,在一定程度上说明鲜塔越重,单塔出籽量越多。
育种方式取决于育种目标,红松是东北长白山等林区重要的乡土树种,长期以来都是以获取木材为目的,但是随着社会的发展,不少研究表明红松松籽富含很高的营养价值和保健价值[43~44],近些年红松更多地被当作经济树种来对待,育种目标也更偏向于用材—坚果兼用型。本研究中,以26和28年生红松的生长性状和结实性状对其半同胞家系进行综合评价,分别筛选出7个生长优良家系和7个结实优良家系,其中仅有家系P113(树高与胸径一般配合力较高)与A124(单塔出籽量与单株球果数一般配合力较高)与一般配合力选出的优良亲本相同,表明单性状评价与多性状评价结果可能不同,具体评价方法需要根据育种目标来选择。
