铅胁迫下干旱对侧柏光合特性与水分利用效率的影响
2020-06-01黄优王进鑫刘俊峰冯树林吕国利
黄优 王进鑫 刘俊峰 冯树林 吕国利
摘要:研究铅胁迫下干旱胁迫对侧柏幼苗光合作用的影响规律,探讨侧柏对双重胁迫的适应与自我调节机制,为西北矿区植被恢复与重建提供理论依据。通过盆栽试验,以土壤相对含水率为100%且无铅的处理为对照,其余处理外源铅浓度统一设为2 000 mg/kg,分别测定侧柏(Platycladus orientalis)幼苗在3个生长阶段水分胁迫条件下的光合速率、蒸腾速率和水分利用效率。结果表明:(1)单一铅胁迫下,侧柏净光合速率较对照降低,铅和干旱双重胁迫下,净光合速率随着土壤相对含水率的降低呈现先增加后降低的趋势。(2)在2 000 mg/kg铅浓度胁迫下,侧柏3个生长阶段净光合速率对水分胁迫的响应不同,当净光合速率达到最大值时,生长初期、中期和后期土壤相对含水率分别为68.64%、67.54%和98.99%。侧柏在生长初期受到干旱胁迫时能迅速调节蒸腾速率以维持体内水分平衡;对模型进行拟合分析发现,当生长初期、中期和后期土壤相对含水量分别为67.52%、78.26%、45.68%时,侧柏蒸腾速率降到最小值。(3)随着土壤相对含水率的降低,侧柏水分利用效率整体呈现上升的趋势,土壤相对含水率为40.00%时,生长初期和中期的水分利用效率最大。因此,在铅胁迫下适度的干旱胁迫可降低侧柏的蒸腾速率,從而提高其水分利用效率。
关键词:侧柏;水分胁迫;铅胁迫;光合速率;蒸腾速率;水分利用效率
中图分类号: S791.380.1;Q945.11文献标志码: A
文章编号:1002-1302(2020)08-0168-06
收稿日期:2019-03-04
基金项目:国家自然科学基金(编号:31670713、31170579);陕西省科技统筹计划(编号:2016KTCL03-18)。
作者简介:黄 优(1994—),女,山西运城人,硕士研究生,研究方向为林草生态工程。E-mail:youhuang572799@163.com。
通信作者:王进鑫,博士,教授,博士生导师,主要从事旱区人工植被生态恢复与环境生态学理论研究。E-mail:jwang118@126.com。
我国西北地区铅锌矿产资源丰富[1],随着工矿业的迅速发展,生态破坏、土壤污染等问题日益严重。由于西北矿区气候干旱,降水稀少,水资源严重短缺,土壤保水能力差,且铅锌矿废弃地重金属含量较高,土壤水分与重金属含量成为限制矿区植被恢复的主要因子[2]。木本植物具有生物量大、生长周期长、便于管理等特点,且以木材形式被利用,不会进入食物链对人体造成危害,因此在矿区植被恢复方面具有很大应用潜力,已成为废弃地植被恢复重点研究的植被类型[3]。
侧柏(Platycladus orientalis)因有抗旱、耐铅、适应性强等特性,成为矿区植被恢复研究的重要植物[4]。低浓度铅对植物生长有促进作用,而浓度过高的铅将抑制植物的生长,有研究表明,侧柏具有一定的抗铅性,并且在土壤铅浓度为2 000 mg/kg时对侧柏生长有一定的促进作用[5]。在一定的干旱胁迫下,植物自身的光合特性和水分生理特性等都会发生相应的变化,不同植物对水分胁迫的适应力不同,同一植物在不同生长阶段对水分的需求也不同[6],只有掌握了植物的水分利用规律,才能更好地促进干旱区植物的健康生长,改善矿区生态环境。目前多数研究是针对单一短期胁迫的,而对植物生长期不同阶段、不同强度双重胁迫光合生理特性的研究鲜有报道。因此,本研究探讨在一定的铅胁迫下不同土壤水分条件对侧柏幼苗的光合特性和水分利用效率的影响,揭示侧柏在西北铅锌矿业废弃地不同土壤水分条件下光合特性与水分利用效率的变化规律,以便充分发挥侧柏的生产潜力,为干旱半干旱铅锌矿区的植被恢复与重建、生态环境的改善提供理论依据。
1 材料与方法
1.1 试验材料与处理
供试植物为1年生侧柏幼苗,来源于陕西省杨凌农业高新技术产业示范区(以下简称杨凌区)附近苗圃,起苗前对圃地苗木进行调查并逐一标记,起苗后精心挑选,确保苗高、地径与长势相对一致,以减少苗木的基础生长引起的随机误差。栽培土壤质地为黏壤,田间持水量为22.3%,土壤铅的背景值为(16.3±2.96) mg/kg[7],土壤有机质含量为13.70 g/kg,全氮含量为0.73 g/kg,速效磷含量为35.91 mg/kg,速效钾含量为96.52 mg/kg;将土壤风干后过筛,除去石砾、结核及林草根系。向土壤中加入99%的分析纯醋酸铅[Pb(CH3COO)2·3H2O],充分混匀后钝化2个月备用;试验期间栽培基质保持自然肥力,不施肥。
1.2 试验设计
试验采用三因素二次通用旋转组合设计,即生长初期(3月22日至6月17日)、生长中期(6月18日至8月15日)、生长后期(8月15日至11月1日)的土壤供水量分别以Z1、Z2、Z3表示,每个因素分别设5个水平,拟定每个因素的上下水平[8-9],上水平以Z2j(田间持水量的100%)表示,下水平以Z1j(田间持水量的40%)表示,则零水平(Z0j)=(Z1j+Z2j)/2=70%,变化间距(Δj)=(Z2j-Z0j)/j=17.84%(j=1.682),因此土壤相对含水率5个水平的编码值和实际值分别为1.682(100.00%)、1(87.84%)、0(70.00%)、-1(52.16%)、-1.682(40.00%),每个处理重复4次,共20个处理。另设1个对照处理(CK),土壤相对含水率为100%且不含外源铅。
盆栽试验在陕西省杨陵区西北农林科技大学南校区内进行,采用旱棚人工控水的方法进行布设。3月份将钝化好的土壤装入内径为27 cm、高为30 cm 的塑料桶中,使每桶中风干土的量一致,土壤铅浓度依据野外调查和前期盆栽试验确定,统一为 2 000 mg/kg。将苗木移栽至塑料桶中,每桶1株,移栽后在土壤表面覆盖1层约2 cm厚的碎石粒,以减少土壤表面水分蒸发而引起的试验误差。之后1个月缓苗期间,充分供水以保证苗木成活,植物生长期间采用人工称质量法来控制土壤相对含水率,每隔2 d称1次质量,及时调整补充水量,以确保土壤水分维持试验设计的土壤相对含水率。
1.3 指标测定
凈光合速率(net photosynthesis rate,Pn)和蒸腾速率(transpiration rate,Tr)采用Li-6400便携式光合测定仪分别于6月11日、8月15日、10月4日08:00—11:00,选取中部成熟叶片进行测定,每个处理测定3个重复;水分利用效率(water use efficiency,WUE)为净光合速率与蒸腾速率的比值。
1.4 数据处理与分析
采用SPSS 20.0进行数据处理,采用Excel 2010作图,采用One-way ANOVE和最小显著差异(LSD)法对数据进行方差分析和多重比较,利用降维法对各因素的单因子效应进行分析。
2 结果与分析
2.1 铅胁迫下不同水分条件对侧柏净光合速率的影响
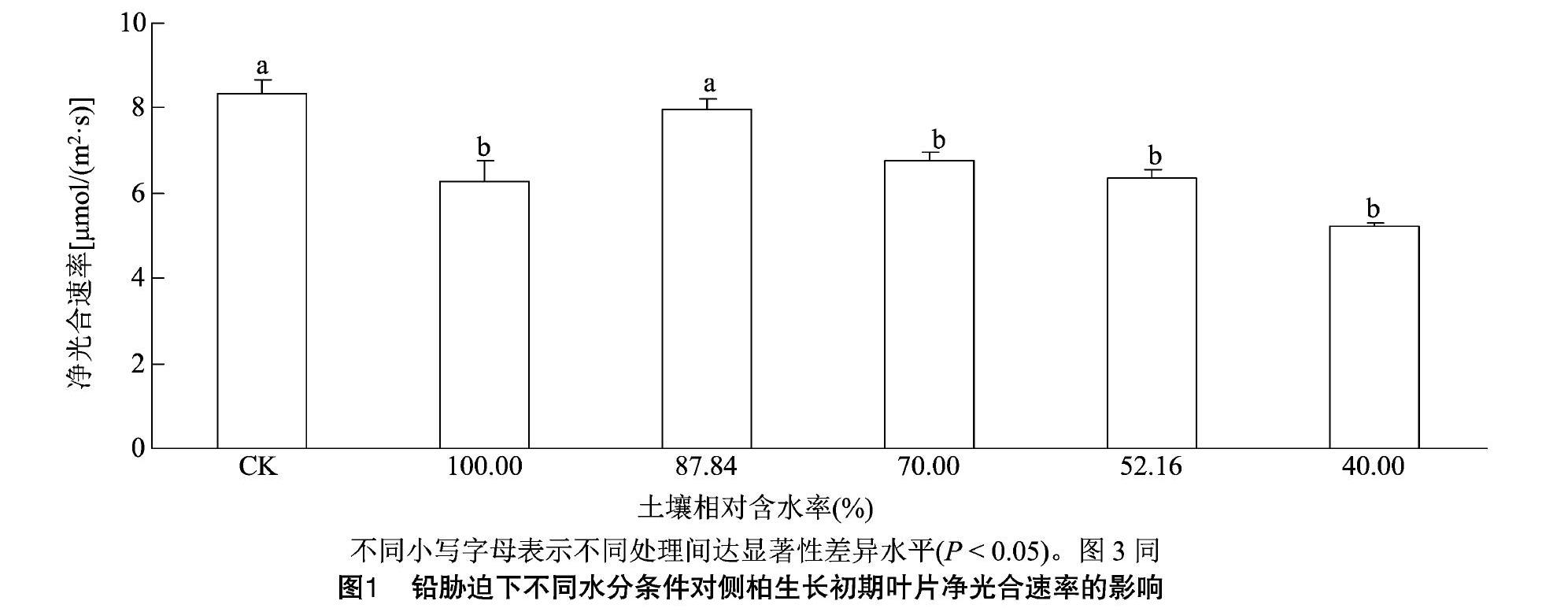
2.1.1 铅胁迫下不同水分条件对侧柏生长初期净光合速率的影响 由图1可知,侧柏在单一铅胁迫下净光合速率低于CK,且差异显著(P<0.05),较CK降低24.66%。在铅浓度均为2 000 mg/kg处理下,随着土壤相对含水率的降低,净光合速率呈现先升高后降低的趋势。在土壤相对含水率为87.84%时,净光合速率达到最大值,比土壤相对含水率为100%时显著增加27.46%(P<0.05)。当土壤相对含水率为40.00%~87.84%时, 净光合速率随土壤相对含水率的降低而降低,其中土壤相对含水率为87.84%的净光合速率显著高于其他3个处理(P<0.05),说明在侧柏生长初期,适宜的水分供应有利于植物进行光合作用。
2.1.2 铅胁迫下不同水分条件对侧柏整个生长期净光合速率的影响 按三因素二次通用旋转组合设计原理,建立侧柏净光合速率(Y)与3个生长时期土壤相对含水率编码值(X)之间的回归方程。
Y=4.989-0.398X1-0.314X2+2.010X3+0.528X21-0.092X22-0.200X23-0.251X1X2+0.089X1X3+0.054X2X3。(1)
式中:X1、X2、X3分别为生长初期、中期、后期净光合速率编码值。
失拟性检验F1=0.928小于F0.05(5,5)=5.05,说明失拟项不显著,回归模型F2=10.146大于F0.01(9,10)=4.94,达到显著水平,说明模型的预测值和实际值吻合程度较高。
根据编码回归模型中的一次项系数可以看出,侧柏苗木生长后期的土壤相对含水率对其叶片的净光合速率影响最大,初期和中期的土壤相对含水率影响较小。通过降维法分析单因素效应,得到单因素影响回归模型。
生长初期:Y=4.989-0.398X1+0.528X21;(2)
生长中期:Y=4.989-0.314X2-0.092X22;(3)
生长后期:Y=4.989+2.010X3-0.200X23。(4)
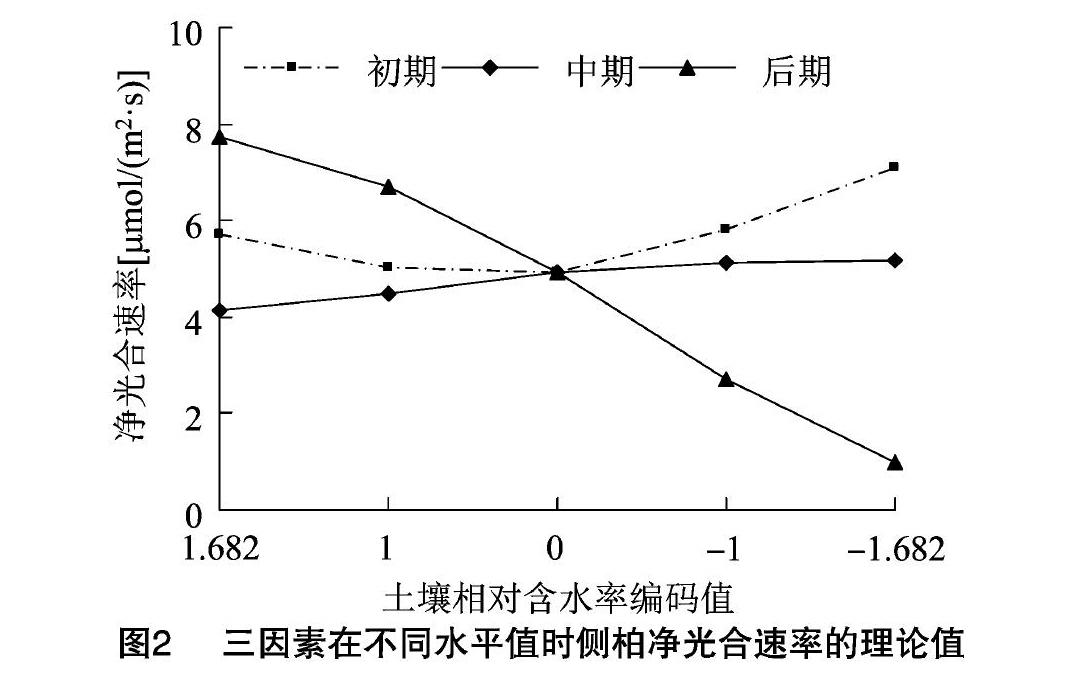
对方程进行显著性检验分析发现,侧柏回归模型中的X1、X21、X2、X3均达到显著水平,说明不同生长时期土壤相对含水率的变化对侧柏的光合速率有显著影响。单因素效应曲线如图2所示。
由互作项偏回归系数可知,初期和中期土壤相对含水率的互作效应(X1X2)大于初期和后期土壤相对含水率的互作效应(X1X3),且它们均大于中期和后期土壤相对含水率的互作效应(X2X3),说明在侧柏苗木生长初期和中期土壤相对含水率的互作效应对苗木叶片的净光合速率影响最大。
当净光合速率达到最大值时,得到X1=-0.076 4、X2=-0.138 0、X3=1.625 3,即对应的土壤相对含水率为初期68.64%、中期67.54%、后期98.99%,此时净光合速率为7.77 μmol/(m·s)。
由图2可以看出,铅胁迫下,在生长初期侧柏净光合速率随土壤相对含水率的减少呈先减少后增加的趋势,在土壤相对含水率为70%时减至最小值;在生长中期侧柏净光合速率随土壤相对含水率的减少而平稳上升;在生长后期则随着土壤相对含水率的减少而大幅度下降。
2.2 铅胁迫下不同水分条件对侧柏蒸腾速率的影响
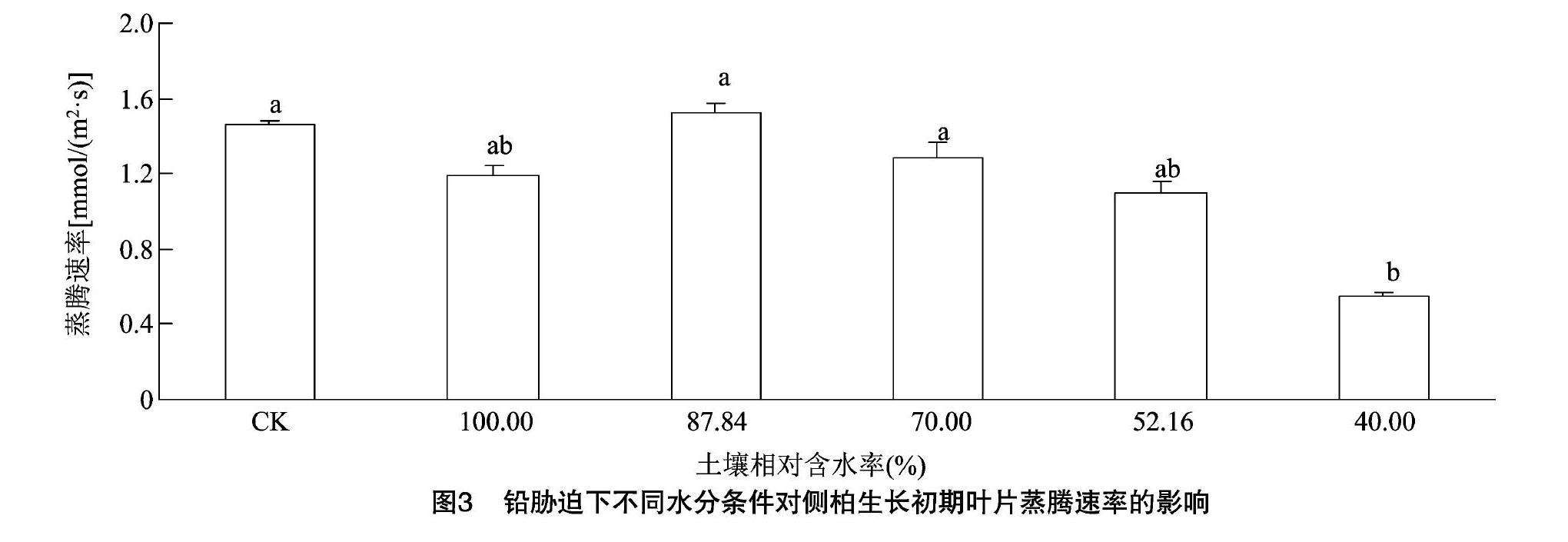
2.2.1 铅胁迫下不同水分条件对侧柏生长初期蒸腾速率的影响 蒸腾作用是植物生命活动中重要的生理活动,植物通过调节蒸腾速率的大小来维持体内水分平衡,水分胁迫能显著影响植物的蒸腾速率[10]。由图3可以看出,侧柏在单一铅胁迫下蒸腾速率较对照有所降低,下降17.93%,但二者差异不显著。在铅浓度均为 2 000 mg/kg 处理下,随着土壤相对含水率的降低,蒸腾速率呈现先升高后降低的趋势,并在土壤相对含水率为87.84%处理下达到最大值。蒸腾速率的下降幅度在一定程度上反映了植物受水分胁迫的强度、植物调节体内水分散失的能力及植物自身适应逆境的方式。与土壤相对含水率为87.84%的处理相比,土壤相对含水率为40.00%、52.16%、70.00%和100.00%处理下的蒸腾速率分别降低了63.78%、27.89%、15.55%和21.59%,只有土壤相对含水率为40.00%的处理差异达显著水平(P<0.05)。说明侧柏在受到干旱胁迫时,能通过迅速降低蒸腾速率,维持体内水分平衡来适应干旱胁迫。
2.2.2 铅胁迫下不同水分条件对侧柏整个生长期蒸腾速率的影响 按三因素二次通用旋转组合设计原理,建立侧柏蒸腾速率(Y′)与3个生长时期土壤相对含水率编码值(X)之间的回归方程。
Y′=2.021-0.128X1-0.245X2+1.024X3+0.141X21-0.021X22-0.001X23-0.113X1X2+0.110X1X3-0.040X2X3。(5)
失拟性检验F1=0.897小于F0.05(5,5)=5.05,表明失拟项不显著,回归模型F2=9.723大于F0.05(9,10)=3.02,达到显著水平,说明模型的预测值和实际值吻合程度较高。
根据编码回归模型中的一次项系数可以看出,侧柏苗木生长后期的土壤相对含水率对其叶片的蒸腾速率影响最大,初期和中期的土壤相对含水率影响较小。通过降维法分析单因素效应,得到单因素影响回归模型。
生长初期:Y′=2.021-0.128X1+0.141X21;(6)
生长中期:Y′=2.021-0.245X2-0.021X22;(7)
生长后期:Y′=2.021+1.024X3-0.001X23。(8)
对方程进行显著性检验分析发现,侧柏回归模型中的X1、X2、X23均达到显著水平,说明不同生长时期土壤相对含水率的变化对侧柏的蒸腾速率有显著影响。单因素效应曲线如图4所示。
由互作項偏回归系数可知,初期和中期土壤相对含水率的互作效应(X1X2)大于初期和后期土壤相对含水率的互作效应(X1X3),且它们均大于中期和后期土壤相对含水率的互作效应(X2X3),说明在侧柏苗木生长初期和中期土壤相对含水率的互作效应对苗木叶片的蒸腾速率影响最大。
当蒸腾速率达到最小值时,得到X1=-0.139 3、X2=0.462 8、X3=-1.363 0,即对应的土壤相对含水率为初期67.52%、中期78.26%、后期45.68%,此时蒸腾速率为0.579 mmol/(m2·s)。
由图4可以看出,铅胁迫下,在生长初期侧柏蒸腾速率随土壤相对含水率的减少呈先减少后增加的趋势,在土壤相对含水率70%时减至最小值;在生长中期侧柏蒸腾速率随土壤相对含水率的减少而平稳上升;在生长后期则随着土壤相对含水率的减少而大幅度下降。
2.3 铅胁迫下不同水分条件对侧柏水分利用效率的影响
水分利用效率反映植物耗水与干物质生产间的关系,是水分生理和光合作用研究中的重要参数之一[11]。试验结果(表1)表明,不同生长时期单一铅胁迫侧柏水分利用效率均显著低于对照。相同铅浓度下,在生长初期侧柏水分利用效率随着土壤相对含水率的降低整体升高,在土壤相对含水率为40.00%处理下达到最大值,且与其他水分处理差异显著(P<0.05);在生长中期水分利用效率普遍低于初期,变化趋势与初期基本一致,在土壤相对含水率为100.00%时达到最小值,且与土壤相对含水率为40.00%和52.16%处理差异显著(P<0.05),但与土壤相对含水率为70.00%和87.84%处理差异不显著;在生长后期最大水分利用效率出现在土壤相对含水率为52.16%处理下,且与其他水分处理差异显著(P<0.05)。说明侧柏在受到重度干旱胁迫时,水分利用效率会有所提高。
3 讨论与结论
3.1 讨论
本研究结果表明,单一铅胁迫(浓度为2 000 mg/kg)处理下,侧柏净光合速率和水分利用效率均低于对照,说明铅胁迫抑制了植物生长,可能是由于该浓度铅胁迫已经破坏了叶绿体和叶绿素的合成,对植物造成了伤害,从而降低了植物净光合速率与水分利用效率[12]。在同一铅浓度下,随着干旱胁迫的加剧,净光合速率呈现先上升后下降的趋势,可能是由于当侧柏受到铅和干旱双重胁迫时,保卫细胞膨压降低,气孔开度减小或关闭[13],阻碍CO2的吸收,从而抑制光合作用的进行[14-15]。本研究结果表明,在铅胁迫下,当侧柏受到严重的水分亏缺(土壤相对含水率40.00%)时,净光合速率降到最小值,与土壤相对含水率为87.84%处理差异显著(P<0.05),此时侧柏叶片的叶绿体结构特别是光合膜系统受到损害,Rubisco活性下降,光系统Ⅱ失活或损伤,电子传递和光合磷酸化活力降低,进而造成了净光合速率的降低[16],可见,非气孔因素对侧柏光合作用的限制起决定作用[17-18]。而在土壤相对含水率大于87.84%时,侧柏的净光合速率有所下降,原因可能是当土壤相对含水率为100.00%时,土壤的通气性较差,影响了根系发育,造成根系活力下降,从而间接造成净光合速率降低,也可能是在土壤相对含水率为100.00%时,侧柏苗木吸收了更多的铅,叶绿体结构被毒害,从而导致净光合速率降低[19]。
有研究表明,植物蒸腾速率随土壤水分胁迫的加剧而降低[3,20]。本研究结果表明,铅胁迫下,当土壤相对含水率小于等87.84%时,随着土壤相对含水率的降低,侧柏的蒸腾速率明显减小;但当土壤相对含水率从100.00%下降至87.84%时,侧柏的蒸腾速率却有所升高。蒸腾速率的变化可能是由于土壤相对含水率为100.00%时,土壤的通气状况较差,而侧柏对土壤通气性要求较高,根系活动受到影响,同时植物受到铅胁迫时,体内活性氧积累增多,光合色素降解,阻碍光合作用正常进行,使得植物蒸腾作用也受到相应抑制[21-24]。当土壤相对含水率降至40.00%水平时,侧柏的蒸腾速率低于其他水平。表明侧柏在受到铅和干旱双重胁迫时,能迅速降低蒸腾速率,且维持较低的蒸腾速率,具有较好的控制失水和维持体内水分平衡的能力[25]。
水分利用效率是净光合速率与蒸腾速率的比值,用来表示植物体本身对蒸腾耗水的利用能力。李文华等对仁用杏在受到单一水分胁迫时的水分和生长特征进行了研究,结果表明,水分利用效率随土壤干旱胁迫的加剧而有所提高,各处理间的水分利用效率差异显著[26]。本研究结果表明,侧柏受到铅和干旱双重胁迫时,叶片的气孔导度由于水分亏缺而降低,光合作用和蒸腾作用同时减弱,但蒸腾作用对气孔导度的依赖高于光合作用,所以侧柏的水分利用效率总体随土壤相对含水率的降低而升高。
3.2 结论
铅和干旱双重胁迫对侧柏的光合作用与水分利用产生多方面的影响。同一铅胁迫下,生长初期进行水分胁迫,侧柏的净光合速率和蒸腾速率均随着干旱程度的加剧呈现先上升后下降的趋势,当土壤相对含水率为40.00%时,净光合速率和蒸腾速率降到最低值,水分利用效率达到最大值。不同生长阶段的双重胁迫对侧柏的影响有所不同,通过对回归方程的拟合分析可知,当生长初期、中期和后期土壤相对含水率分别为68.64%、67.54%、98.99%时,侧柏净光合速率达最大值;土壤相对含水量分别为67.52%、78.26%、45.68%时,蒸腾速率降到最小值,说明合适的水分胁迫可促进侧柏净光合速率的增加,降低蒸腾速率,从而使侧柏达到最高的水分利用效率。
参考文献:
[1]苏 帅,杨永刚,黄 磊. 矿区生态修复过程中不同立地类型土壤水动力学特性[J]. 水土保持通报,2018,38(1):18-23.
[2]徐玉霞,汪庆华,王华斌,等. 矿区周边土壤重金属污染治理的优势植物筛选——以关中西部某铅锌冶炼区为例[J]. 环境保护科学,2016,42(6):61-67.
[3]陈科皓,王进鑫. 紫穗槐生长前期对铅胁迫下不同土壤水分的生长和生理响应[J]. 草地学报,2015,23(4):883-886.
[4]李春燕,王进鑫,王 敏,等. 铅胁迫下干旱对国槐和紫穗槐幼苗抗氧化酶活性的影响[J]. 干旱地区农业研究,2016,34(3):174-178.
[5]周芙蓉,王进鑫,杨 楠,等. 干旱和铅交互作用对侧柏幼苗生长及抗氧化酶活性的影响[J]. 林业科学,2013,49(6):172-177.
[6]张永娥,余新晓,陈丽华,等. 不同土壤含水量下侧柏幼树叶片水分利用效率[J]. 应用生态学报,2017,28(7):2149-2154.
[7]夏增禄. 土壤元素背景值及其研究方法[M]. 北京:气象出版社,1987.
[8]陈科皓. 二次通用旋转组合设计下耐旱树种的生长特性研究[J]. 林业科技通讯,2018(1):3-7.
[9]余新晓,张建军,朱金兆. 黄土地区防护林生态系统土壤水分条件的分析与评价[J]. 林业科学,1996,32(4):289-297.
[10]于金凤,刘文兆,甘卓亭,等. 黄土塬区苹果树蒸腾速率变化特征及其影响因子[J]. 干旱地区农业研究,2010,28(4):59-63.
[11]张岁岐,山 仑. 植物水分利用效率及其研究进展[J]. 干旱地区农业研究,2002,20(4):1-5.
[12]初江涛,王进鑫,邹 朋,等. 干旱和铅胁迫对生长初期的国槐和侧柏叶绿素的影响[J]. 西北林学院学报,2012,27(4):19-23.
[13]贾瑞丰,徐大平,杨曾奖,等. 干旱胁迫对降香黄檀幼苗光合生理特性的影响[J]. 西北植物学报,2013,33(6):1197-1202.
[14]Lagergren F,Lindroth A. Transpiration response to soil moisture in pine and spruce trees in Sweden[J]. Agricultural and Forest Meteorology,2002,112(2):67-85.
[15]杨建伟,赵 丹,孙桂芳,等. 干旱胁迫对小紫珠光合生理的影响[J]. 西北植物学报,2018,38(4):733-740.
[16]杨 锐,郎 莹,张光灿,等. 野生酸枣光合及叶绿素荧光参数对土壤干旱胁迫的响应[J]. 西北植物学报,2018,38(5):922-931.
[17]Zlatev Z,Lidon F C. An overview on drought induced changes in plant growth,water relations and photosynthesis[J]. Emirates Journal of Food and Agriculture,2012,24(1):520-524.
[18]柴胜丰,唐健民,王满莲,等. 干旱胁迫对金花茶幼苗光合生理特性的影响[J]. 西北植物学报,2015,35(2):322-328.
[19]王榆鑫,王进鑫,初江涛,等. 侧柏和国槐幼苗生长对铅胁迫的阈值[J]. 江苏农业科学,2017,45(1):123-127.
[20]沙依然·外力,李 晨,葛道阔,等. 干旱胁迫下水稻光合特性、冠层-空气温差及水分利用效率的变化[J]. 江苏农业科学,2016,44(1):86-89.
[21]接玉玲,杨洪强,崔明刚,等. 土壤含水量與苹果叶片水分利用效率的关系[J]. 应用生态学报,2001,12(3):387-390.
[22]徐 飞,郭卫华,徐伟红,等. 刺槐幼苗形态、生物量分配和光合特性对水分胁迫的响应[J]. 北京林业大学学报,2010,32(1):24-30.
[23]李文华,刘广权,马松涛,等. 干旱胁迫对苗木蒸腾耗水和生长的影响[J]. 西北农林科技大学学报(自然科学版),2004,32(1):61-65.
[24]颜淑云,周志宇,邹丽娜,等. 干旱胁迫对紫穗槐幼苗生理生化特性的影响[J]. 干旱区研究,2011,28(1):139-145.
[25]张慕黎,王进鑫,李继文,等. 充分供水下刺槐和侧柏苗木光合特性及水分利用效率的研究[J]. 西北林学院学报,2009,24(3):27-32.
[26]李文华,吴万兴,张忠良,等. 土壤水分对仁用杏水分和生长特征的影响[J]. 西北农林科技大学学报(自然科学版),2003,31(4):139-144.
