盐胁迫对酸柚苗光合作用和荧光特性的影响
2020-06-01金星宇马佳冯美
金星宇 马佳 冯美
摘要:以酸柚二年生实生苗为试验材料,进行0%、0.3%、0.6%、0.9%浓度的NaCl处理,研究在NaCl胁迫下酸柚苗光合作用和叶绿素荧光参数的变化。结果表明,盐胁迫使酸柚叶片叶绿素含量、蒸腾速率(Tr)、最大荧光(Fm)、PSⅡ 最大光化学效率(Fv/Fm)、PSⅡ实际光化学效率(ΦPSⅡ)、PSⅡ有效光化学量子产量(Fv′/Fm′)、光化学反应猝灭系数(qP)下降,初始荧光(Fo)和非光化学猝灭系数(NPQ)逐步上升,净光合速率(Pn)、气孔导度(Gs)、潜在光化学效率(Fv/Fo)先升高后降低,胞间CO2浓度(Ci)先降低后升高。0.3%、0.6% NaCl处理对酸柚苗造成了光抑制,其净光合速率下降的原因主要是气孔限制;0.9% NaCl处理对酸柚苗的影响较大,其净光合速率下降的原因主要是非气孔限制,酸柚苗将能量转变为热量耗散掉,以缓解盐胁迫对光合作用的影响。由结果可以看出,在盐胁迫下,酸柚能保护内部的光合机构功能状态,从而提高对NaCl胁迫的适应能力。
关键词:酸柚苗;NaCl胁迫;光合作用;荧光特性;叶绿素荧光参数
中图分类号: Q945.78文献标志码: A
文章编号:1002-1302(2020)08-0159-05
收稿日期:2019-03-22
基金项目:宁夏回族自治区科技支撑计划(编号:00315-0426);园艺作物优质高效安全生产产学研合作基地(编号:201606)。
作者简介:金星宇(1994—),男,宁夏吴忠人,主要从事园艺植物栽培研究。E-mail:1219367768@qq.com。
通信作者:冯 美,硕士,教授,主要从事果树栽培及栽培生理研究。E-mail:13895186355@126.com。
土壤盐渍化是一个全球性的生态问题,世界上约有20%的可耕地受到了盐渍的影响,盐胁迫使农业生产受到了严重损失。宁夏地处我国西北地区,盐渍化面积约为14.8万hm2,而且由于土壤次生盐渍化,盐渍化面积还在不断增加[1]。盐胁迫会通过抑制植物的光合作用而影响植物的生长发育。目前,许多学者关于盐胁迫对不同植物光合作用和荧光特性的影响进行了大量研究,已有大量成果发表[2-4]。许多研究表明,盐胁迫使植物的光合速率下降,影响植物组织的光系统Ⅱ(PSⅡ)光化学效率,但是由于受胁迫的植物、胁迫浓度及时间不同,关于抑制光合作用的原因仍然结论不一[5-7]。不同植物对盐胁迫的适应机制不尽相同,研究和掌握植物的耐盐机制,筛选能适应高盐环境的植物,对于提高盐渍化土地利用率具有重要意义。
酸柚(Citrus grandis)为芸香科(Rutaceae)乔木,一般作为柚的砧木,与柚栽培品种间的嫁接亲和性好,适宜土层深厚、肥沃、排水良好的土壤,也较耐碱性土壤,但抗寒性较枳砧差。宁夏光热资源丰富,秋季凉爽,发展设施果树具有良好的自然资源基础。为了丰富宁夏设施果树的树种结构,2015年3月宁夏大学从四川省遂宁市永红矮晚柚有限公司引进了用酸柚作砧木的遂宁矮晚柚品种。针对宁夏土壤盐碱化严重的问题,本研究以2年生盆栽酸柚实生苗作为试验材料,探讨NaCl胁迫对其光合作用和荧光特性的影响,分析其对盐胁迫的适应性,以期为酸柚砧木苗在宁夏设施果树中的栽培推广提供一定的理论基础。
1 材料与方法
1.1 试验材料
本试验以2年生盆栽酸柚实生苗作为材料,该酸柚苗种植在宁夏大学农科实训基地温室内,按常规管理。
1.2 试验方法
在实验室通过枯黄率预试验,初步确定酸柚NaCl胁迫的临界浓度为1.1%。于2017年5月选择生长均匀一致的2年生酸柚苗定植于花盆中,每盆称取田土11 kg,为了防止溶液渗漏,在每个盆下垫有托盘,按照常规管理。于2017年7月17日进行盐胁迫处理,设置0%(CK)、0.3%、0.6%、0.9%4个NaCl胁迫浓度,每个处理设3盆(3株),重复3次,所有植株都挂牌编号,第1次处理(浇灌)后,每隔 7 d 用1 000 mL相应处理浓度的溶液灌溉,CK浇施等量清水,直至胁迫结束。于处理后49d测定光合和荧光参数。
1.3 测定方法
1.3.1 叶绿素含量的测定 参照高俊凤的方法[8],取新鲜叶片,用80%丙酮提取叶绿素,分别测定D645 nm、D633 nm。
1.3.2 光合作用气体交换参数和荧光参数的测定 采用美国LI-COR公司生产的LI-6400光合测定系统,于盐胁迫49 d的09:00—11:30测定酸柚苗上数第2张叶的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)。
采用IMAGING-PAM全叶叶绿素荧光仪于处理49 d的09:00—11:30测定酸柚苗上数第2张叶的光反应下初始荧光(Fo)、最大荧光(Fm)、稳态荧光(Fs)、PSⅡ实际光化学效率(ΦPSⅡ),并测定暗适应30 min后的初始荧光(Fo′)、最大荧光(Fm′)、PSⅡ 最大光化学效率(Fv/Fm),此外,计算潜在光化学效率(Fv/Fo)[Fv/Fo =(Fm-Fo)/Fo]、PSⅡ有效光化学量子产量(Fv/′Fm′)[Fv/′Fm′=(Fm′-Fo′)/Fm′]、非光化学猝灭系数(NPQ)[NPQ =Fm/Fm′-1]、光化学反应猝灭系数(qP)[qP=(Fm′-Fs)/(Fm′-Fo′)][8]。
1.4 数据分析
用Excel、SPSS軟件进行数据处理与分析。
2 结果与分析
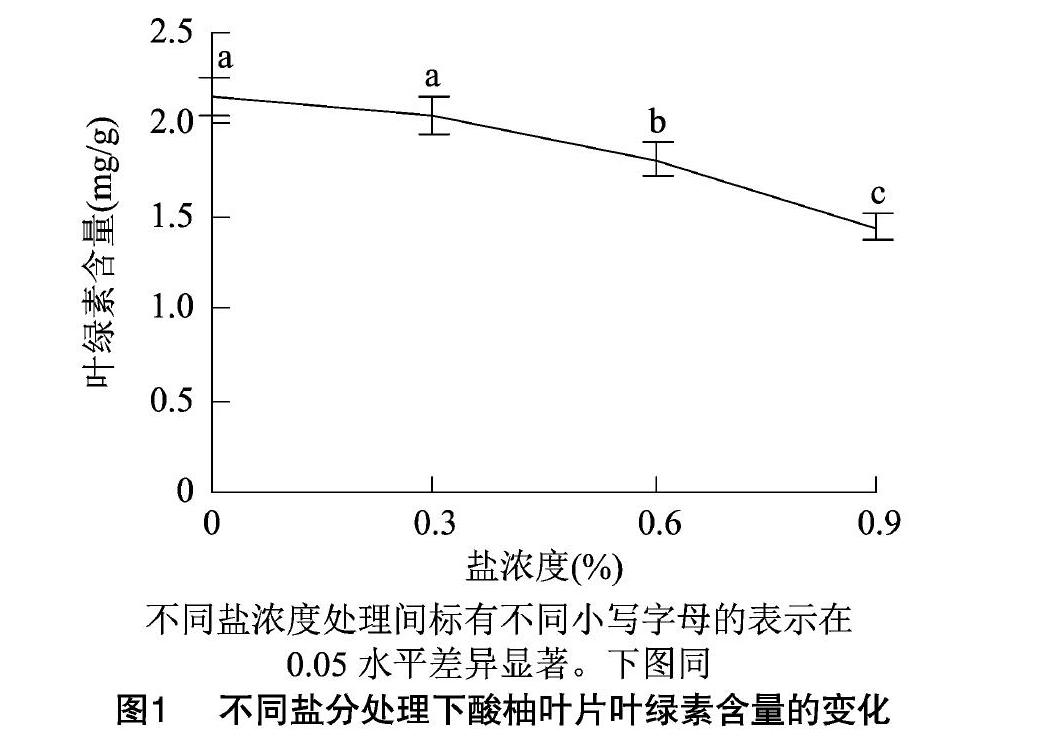
2.1 盐胁迫对酸柚苗叶绿素含量的影响
由图1可见,随着盐胁迫增强,叶绿素含量呈逐渐下降的趋势,在0.9% NaCl处理下,叶绿素含量降到最低值;0.3%NaCl处理的叶绿素含量与对照间无显著差异,0.6%、0.9%NaCl处理的叶绿素含量均与对照、0.3%NaCl处理间存在显著差异。在0.3%、0.6%、0.9% NaCl胁迫处理下,酸柚苗的叶绿素含量分别比对照降低4.56%、15.98%、33.33%。
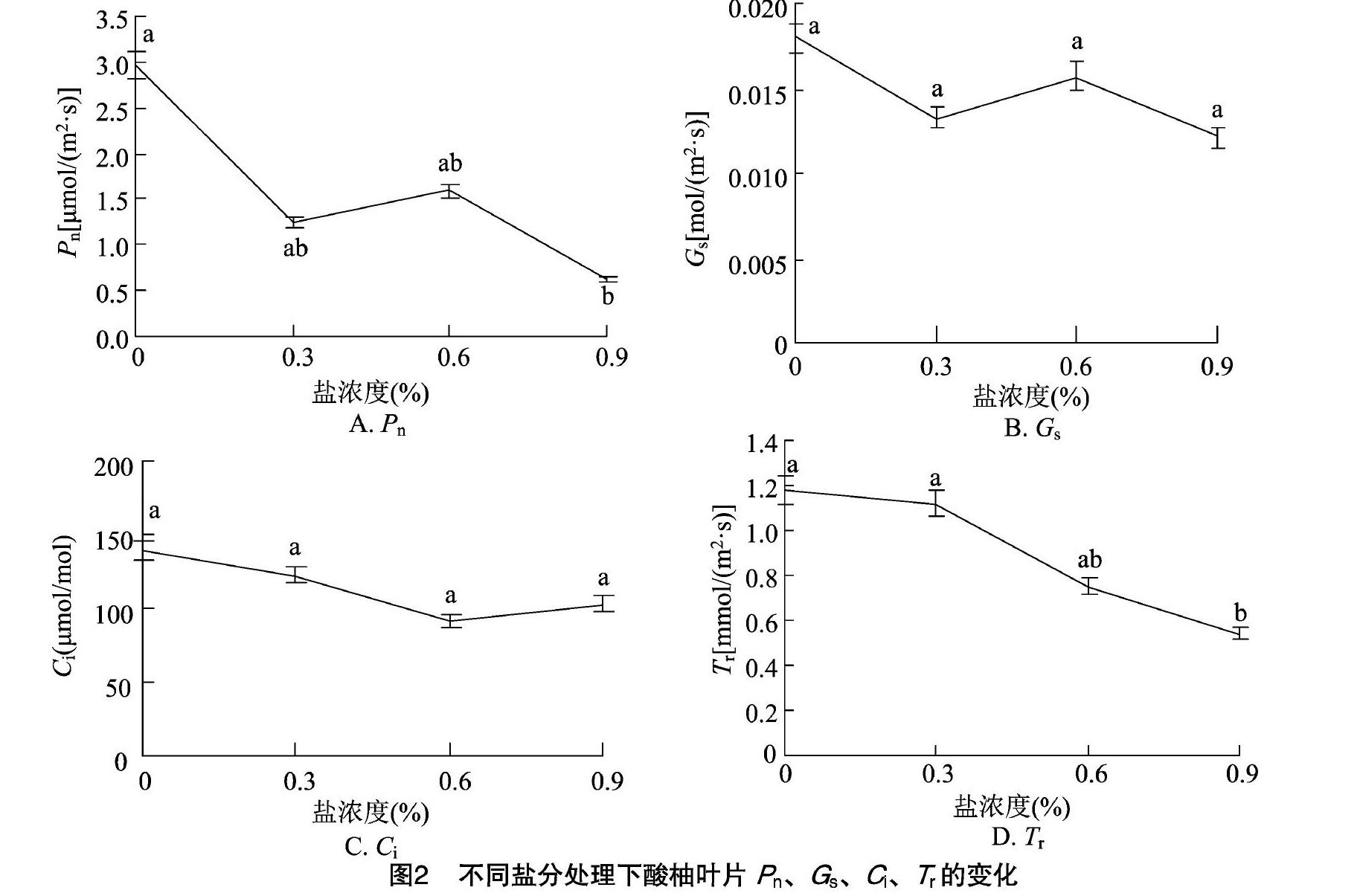
2.2 盐胁迫对酸柚苗光合作用的影响
2.2.1 净光合速率 由图2-A可以看出,在盐胁迫49 d后,随着盐胁迫浓度的增加,2年生酸柚苗净光合速率呈现先下降后上升再下降的变化趋势;0.3%、0.6% NaCl胁迫处理的净光合速率与对照间均无显著差异,0.9% NaCl胁迫处理的净光合速率与对照间存在显著差异,但與0.3%、0.6% NaCl处理间的差异不显著。在0.3%、0.6%、0.9%NaCl胁迫处理下,酸柚苗的净光合速率分别比对照降低了57.78%、46.28%、79.39%。
2.2.2 气孔导度 由图2-B可见,盐胁迫使酸柚苗的气孔导度有所下降,呈现先下降后上升再下降的变化趋势,与净光合速率的变化趋势相似,但是不同处理间的气孔导度差异均不显著。在0.3%、0.6%、0.9% NaCl胁迫处理下,酸柚苗的气孔导度分别比对照降低25.69%、12.29%、32.40%。
2.2.3 胞间CO2浓度 如图2-C所示,随着盐胁迫浓度的增加,酸柚苗的胞间CO2浓度呈先下降后上升的趋势,盐胁迫后的胞间CO2浓度均比对照有所降低,但是不同处理间的胞间CO2浓度差异均未达到显著水平。在0.3%、0.6%、0.9% NaCl胁迫处理下,酸柚苗的胞间CO2浓度分别比对照降低了11.87%、34.09%、26.18%。
2.2.4 蒸腾速率 如图2-D所示,随着盐处理浓度的增加,酸柚苗的蒸腾速率呈下降趋势;0.3%、0.6% NaCl胁迫处理的蒸腾速率与对照间无显著差异,0.9% NaCl胁迫处理的蒸腾速率与对照、0.3%处理间存在显著差异,而与0.6%处理间的差异不显著。在0.3%、0.6%、0.9% NaCl胁迫处理下,酸柚苗的蒸腾速率分别比对照降低了5.13%、36.75%、53.85%。
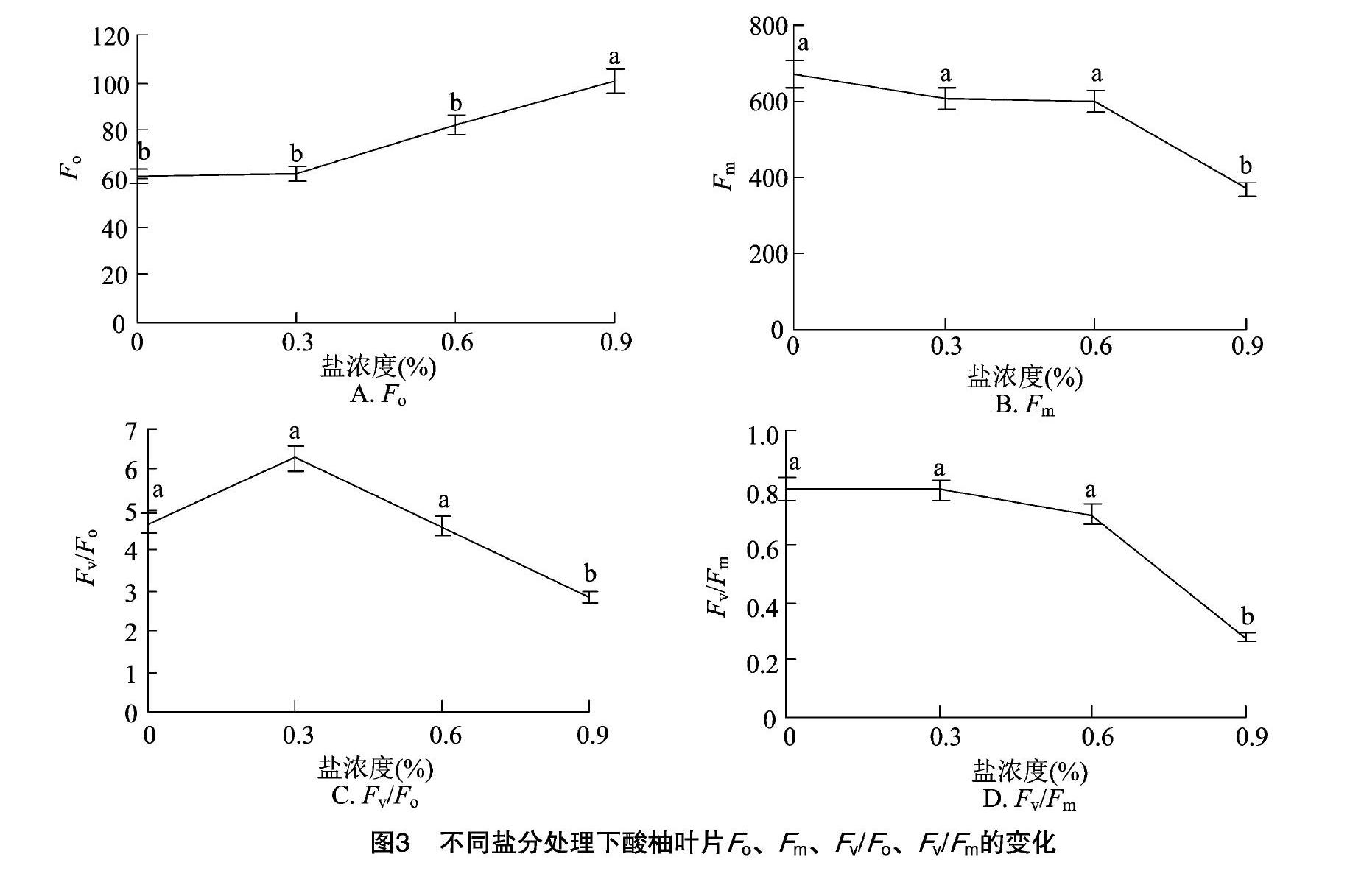
2.3 盐胁迫对酸柚苗荧光参数的影响
2.3.1 最大光化学效率 Fo为最小荧光,也称为初始荧光,是光系统Ⅱ反应中心处于完全开放状态时的荧光产量[9]。如图3-A所示,酸柚苗在受到盐胁迫后,Fo随着NaCl胁迫浓度的增加而逐渐升高;在0.9% NaCl胁迫处理下的Fo与对照、0.3%、0.6%处理间存在显著差异,其余处理间的差异均未达到显著水平。在0.9% NaCl胁迫处理下,酸柚苗的初始荧光比对照提高了63.99%。
Fm为最大荧光,是暗适应下PSⅡ反应中心处于完全关闭状态下的荧光产量,可以反映通过PSⅡ的电子传递情况[9]。如图3-B所示,Fm随着NaCl胁迫浓度的增加而逐渐降低,在0.3%、0.6%盐胁迫下,Fm的降低幅度较小,均与对照间无显著差异;在高浓度(0.9%)NaCl胁迫处理下,酸柚苗的Fm与其他处理间存在显著差异,比对照降低了41.53%。
Fv/Fo代表PSⅡ的潜在光化学效率,如图3-C所示,随着盐胁迫的加大,酸柚苗的Fv/Fo呈先上升后下降的变化趋势,0.3%、0.6% NaCl胁迫下的Fv/Fo与对照间均无显著差异;在高浓度(0.9%)NaCl胁迫下的Fv/Fo与其他处理间均存在显著差异。在0.9% NaCl胁迫处理下,酸柚苗的Fv/Fo比对照降低了39.23%。
Fv/Fm能够反映PSⅡ的原初最大光能利用效率,从而反映植物潜在的最大光合能力。如图3-D所示,Fv/Fm随着NaCl胁迫浓度的增加呈现降低的趋势,0.3%、0.6% NaCl胁迫处理的Fv/Fm与对照间均无显著差异;在高浓度(0.9%)NaCl胁迫下,酸柚苗的Fv/Fm与其他处理间均存在显著差异,比对照降低了64.56%。
2.3.2 荧光猝灭动力学 Fv′/Fm′是PSⅡ有效光化学量子产量。由图4-A可知,随着盐胁迫的加大,酸柚苗的Fv′/Fm′表现出下降的变化趋势;0.3%、0.6% NaCl胁迫处理的Fv′/Fm′与对照间均无显著差异;在高浓度(0.9%)NaCl胁迫下的Fv′/Fm′与其他处理间均存在显著差异,在0.9% NaCl胁迫处理下,酸柚苗的Fv′/Fm′比对照降低了62.96%。
qP是光化学反应猝灭系数,可以反映光化学活性的高低,其值越大,说明PSⅡ的电子传递活性越大。由图4-B可知,随着盐胁迫的加大,酸柚苗的qP呈现下降的变化趋势,0.3% NaCl胁迫处理的qP与对照间无显著差异,而0.6%、0.9% NaCl胁迫处理的qP与对照间均存在显著差异,0.3%、0.6%、0.9% NaCl胁迫处理间的qP不存在显著差异。与对照相比,0.3%、0.6%、0.9% NaCl胁迫处理的qP分别降低了17.07%、24.39%、31.71%。
NPQ反映了光能耗散为热的能力,由图4-C可知,盐胁迫使酸柚苗的NPQ逐渐提高,在高浓度(0.9%)NaCl胁迫处理下,酸柚苗的热耗散能力较强,与对照及0.3%、0.6% NaCl处理间的差异均显著,比对照增加了182.47%。由此看出,过剩的光能以热能的形式耗散,减小了对光系统的破坏。
ΦPSⅡ反映植物在光照下PSⅡ的实际光化学量子效率,较高的ΦPSⅡ能促进碳同化的高效运转和有机物的积累。由图4-D可知,随着盐浓度升高,ΦPSⅡ总体呈现出下降的趋势,0.3%、0.6%、0.9% NaCl胁迫处理的ΦPSⅡ均与对照间差异显著。0.3%、 0.6% NaCl 胁迫处理与0.9% NaCl胁迫处理间的差异显著。与对照相比,0.3%、0.6%、0.9% NaCl胁迫处理的ΦPSⅡ分别下降34.78%、43.48%、68.69%。
3 讨论与结论
在不同逆境胁迫下,一般会造成植物叶片的叶绿素含量降低、净光合速率下降[10-12]。本研究结果表明,在盐胁迫后,酸柚苗的叶绿素含量、气孔导度、净光合速率、蒸腾速率、胞间二氧化碳浓度均比对照有所下降,说明盐胁迫可能使植物的叶绿体结构破坏,但酸柚苗的气孔导度、胞间二氧化碳浓度在各处理间无显著差异,说明在NaCI胁迫下,酸柚苗仍能很好地维持气孔开放。净光合速率是一个对逆境胁迫比较敏感的生理指标,净光合速率下降既有气孔因素,又有非气孔因素[13-15]。在低浓度NaCl胁迫下,当酸柚叶片的气孔导度下降时,胞间二氧化碳浓度也随之下降,说明在低浓度NaCl胁迫下,光合速率下降主要是由于不良环境导致气孔关闭,而在高浓度NaCl胁迫下,酸柚叶片的胞间二氧化碳浓度反而升高,说明气孔没有对光合作用的原料CO2造成限制,也表明在此浓度盐胁迫下,酸柚苗光合速率降低的原因由气孔限制转为非气孔限制,这与馆溪蜜柚的研究结果类似[16],说明酸柚具有较强的耐盐性。
荧光参数的变化主要反映植物光合作用中的能量传递与转化,通过测量叶绿素荧光参数的变化可以推知光能用于光化学反应及用于热耗散等过程的多少[17-21]。本研究结果表明,在盐胁迫下,Fm、Fv/Fm呈下降趋势,Fv/Fo呈先升高后下降的趋势,说明酸柚叶片在受到盐胁迫时,适度过量的光照引起了碳同化量子效率下降。Fo上升,表明盐胁迫后的光系统Ⅱ受到伤害,使得光系统Ⅱ的原初光能转换效率降低,叶绿素吸收的能量中以荧光和热耗散途径散失的比例升高,这与张璐颖等对盐胁迫下台湾桤木的研究结果[21]类似。在盐胁迫下,酸柚叶片的Fv′/Fm′、ΦPSⅡ、qP均呈下降趋势,而NPQ呈上升趋势,说明在盐胁迫下,酸柚叶片在光能捕获后能有效保持光化学电子传递和热耗散途径,从而保护了光合机构,减缓了浄光合速率的下降[9,22]。
NaCl胁迫降低了酸柚叶片的净光合速率,由于胁迫处理后酸柚叶片叶绿素含量、蒸腾速率下降,气孔导度先升高后降低,胞间CO2浓度先降低后升高。0.3%、0.6% NaCl低浓度盐胁迫对酸柚苗的影响不大,光合速率下降的原因主要是气孔限制;0.9% NaCl高浓度盐胁迫对酸柚苗的影响较大,光合速率下降的原因主要是非气孔限制。在NaCl胁迫下,最大荧光、最大光化学效率、PSⅡ实际光化学效率、PSⅡ原初最大光能利用效率、光化学反应猝灭系数下降,初始荧光和非光化学猝灭系数逐步上升,说明低浓度盐胁迫造成了光合速率下降,当盐浓度较高时,酸柚苗通过热耗散途径来缓解盐胁迫对光合作用的影响。
参考文献:
[1]李凤霞,郭永忠,王学琴,等. 不同改良措施对宁夏盐碱地微生物及苜蓿生物量的影响[J]. 中国农学通报,2012,28(30):49-55.
[2]李 宏,鄧江宇,张振春,等. 盐胁迫对盐桦幼树光合特性的影响[J]. 新疆农业科学,2010,47(2):213-217.
[3]葛江丽,石 雷,谷卫彬. 盐胁迫条件下甜高粱幼苗的光合特性及光系统Ⅱ功能调节[J]. 作物学报,2007,33(8):1272-1278.
[4]杨敏生,李艳华,梁海永,等. 白杨派杂种无性系及其亲本光合和生长对盐胁迫的反应[J]. 林业科学,2006,42(4):19-26.
[5]刘晓龙,徐 晨,徐克章,等. 盐胁迫对水稻叶片光合作用和叶绿素荧光特性的影响[J]. 作物杂志,2014(2):88-92.
[6]王东明,贾 媛,崔继哲. 盐胁迫对植物的影响及植物盐适应性研究进展[J]. 中国农学通报,2009,25(4):124-128.
[7]周 琦,祝遵凌. 盐胁迫对鹅耳枥幼苗光合作用和荧光特性的影响[J]. 林业科技开发,2015,29(2):35-40.
[8]高俊凤. 植物生理学实验指导[M]. 北京:高等教育出版社,2006:74-79.
[9]孙 璐. 高粱耐盐品种筛选及耐盐机制研究[D]. 沈阳:沈阳农业大学,2012:35-46.
[10]王清华,杨建平,张中华,等. 盐胁迫对不同品种辣椒种子萌发特性的影响[J]. 西北农业学报,2007,16(3):136-140.
[11]裘丽珍,黄有军,黄坚钦,等. 不同耐盐性植物在盐胁迫下的生长与生理特性比较研究[J]. 浙江大学学报(农业与生命科学版),2006,32(4):420-427.
[12]许大全,丁 勇,沈允钢. C4植物玉米叶片光合效率的日变化[J]. 植物生理学报,1993,19(1):43-48.
[13]周 建,杨立峰,郝峰鸽,等. 低温胁迫对广玉兰幼苗光合及叶绿素荧光特性的影响[J]. 西北植物学报,2009,29(1):136-142.
[14]赵相生,张承烈. 河西走廊芦苇在不同盐渍生境中RuBp羧化酶的比较研究[J]. 应用生态学报,1994,5(2):152-155.
[15]陈建明,俞晓平,程家安. 叶绿素荧光动力学及其在植物抗逆生理研究中的应用[J]. 浙江农业学报,2006,18(1):51-55.
[16]马翠兰. 柚(Citrus grandis Osbeck)对盐胁迫的生理反应及适应性研究[D]. 福州:福建农林大学,2002:45-57.
[17]孙 璐,周宇飞,李丰先,等. 盐胁迫对高粱幼苗光合作用和荧光特性的影响[J]. 中国农业科学,2012,45(16):3265-3272.
[18]刘 强,王庆成,王占武. 叶绿素荧光动力学参数作为植物抗盐性评价指标的有效性[J]. 广东农业科学,2014(21):37-40.
[19]林 晗,陈 辉,吴承祯,等. 千年桐种源间叶绿素荧光特性的比较[J]. 福建农林大学学报(自然科学版),2012,41(1):34-39.
[20]柯裕州,周金星,卢 楠,等. 盐胁迫对桑树幼苗光合生理及叶绿素荧光特性影响[J]. 林业科学研究,2009,22(2):200-206.
[21]张璐颖,文 笑,林勇明,等. 盐胁迫对台湾桤木幼苗光合作用和荧光特性的影响[J]. 福建林学院学报,2013,33(3):193-199.
[22]张守仁. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学通报,1999,16(4):414-448.
