珍稀植物西康玉兰的果实性状特征及其表型多样性
2020-06-01袁丛军李鹤陈锐余德会
袁丛军 李鹤 陈锐 余德会

摘要:为了揭示贵州省西康玉兰主要原产地的果实表型变异程度和变异规律,以该种主要分布区内的4个天然群体果实为试材,对果质量FG、果宽FW、果长FL、果径FD、果型指数FL/FD等2类14个表型性状进行系统的比较分析,采用方差分析、相关分析等方法对不同种源地、不同表型性状的多样性进行探讨。结果表明:(1)同一果实表型性状在不同种源地间、不同果实表型性状在各种源地表现各异。(2)不同果实表型性状间的相关性不同,果质量FG、果径FD、果厚FT、果长FL、果宽FW 5个性状指标间均呈极显著正相关(P<0.01);果型指数FL/FD除与种实率TSWC/FG呈负相关、与单粒种子质量TSW/SN呈显著正相关(P<0.05)外,与其余性状指标均呈极显著正相关(P<0.01);果实体积指数FVI除与种实率TSWC/FG、单粒种子质量TSW/SN呈正相关外,与其余性状指标均呈极显著正相关(P<0.01)。(3)海拔、年均温与多数果实性状间呈显著相关关系,说明它们对西康玉兰果实内、外表型性状的影响较大。(4)西康玉兰14个果实性状中,平均多样性指数介于1.538~1.827;除果径FD、果宽FW、果长FL等3个果实性状表型变异在赫章六曲河镇(HZLQ)种源地最为丰富外,其余11个果实性状均在水城玉舍林场(SCYS)种源地最为丰富。群体的果实外表型性状多样性指数(1.674)高于内表型性状多样性指数(1.671)。(5)不同种源地的果实内、外表型性状多样性指数不同,其中赫章六曲河镇(HZLQ)、盘州老黑山(PZHS)种源地的果实外表型性状多样性指数均分别高于果实内表型性状多样性指数;水城玉舍林场(SCYS)、水城营盘乡(SCYP)种源地的则相反。研究结果为西康玉兰种质资源的保存和育种策略的制定奠定基础。
关键词:西康玉兰;果实性状特征;聚合蓇葖果;表型多样性指数;变异特征;贵州省
中图分类号: S685.150.1文献标志码: A
文章编号:1002-1302(2020)08-0142-07
收稿日期:2019-03-19
基金项目:贵州省林业厅青年科技人才培养对象专项(编号:黔林科合J字 [2015]15号);贵州省林业科学研究项目(编号:黔林科合[2016]15号);贵州省科技计划 (编号:黔科合平台人才[2016]5711)。
作者简介:袁丛军(1990—),男,贵州安顺人,硕士,助理研究员,主要从事恢复生态学、植物学分类与保护研究。E-mail:ycongjun2012gzdx@163.com。
通信作者:李 鹤,贵州铜仁人,硕士,工程师,主要从事木本植物分类学及濒危植物保育研究。E-mail:1043630529@qq.com。
西康玉兰[Magnolia wilsonii (Finet et Gagn) Rehd.]为木兰科(Magnoliaceae)木兰属(Magnolia)天女木兰组(Sect. Oyama)植物,落叶灌木至小乔木,分布于四川中部和西部、云南北部,在贵州兴义、安顺、水城、纳雍、盘州等地有分布,该种属中国特有种,国家二级保护植物,由于其茎皮作为中药“厚朴”的代用品,所以资源破坏严重;此外,滥伐森林,生境恶化,天然繁殖能力较弱,使其自然更新十分困难,其成年植株已不多见[1-2]。目前,关于西康玉兰研究报道较少,郭应等研究了盘州西康玉兰群落结构、种群动态及繁殖特征[3];吴守清等开展了水城玉舍林场的西康玉兰种子储藏及种苗繁育技术研究[4];韓春艳等研究了西康玉兰的种子休眠、萌发及贮藏特性[5]。已有学者开展针对花[6-7]、果实[8]、种子[9-10]、叶片[11]、枝条[12]等不同繁殖体性状和海拔[13-14]、光照[15]等环境因子下的不同植物表型性状多样性研究。 西康玉兰是木兰属较原始种类,对本属的系统发育研究具有一定的科学价值。目前,有关西康玉兰种质资源变异研究较少,有关西康玉兰的果实和种子特征及性状表型变异研究未见报道。
表型(phenotype)一般指生物个体或群体,在特定条件下(如各类环境和生长阶段等)所表现出的可观察的形态特征[16]。植物表型研究核心是获取高质量、可重复的性状数据,进而量化分析基因型和环境互作效应(genotype-by-environment interactions,GxE)[17]。表型性状遗传变异如何影响种群个体的功能性状从而影响种群适合度[18],植物表型多样性作为遗传多样性与环境多样性的综合体现,是了解植物遗传变异的重要线索和生物多样性的重要内容[19]。刘艳萍等研究了广玉兰果实和种子的表型变异情况[20];宋杰等研究了云南昆明地区云南含笑天然分布的6个居群180个单株的14个表型性状的表型多样性特征[21];文珊娜等研究了灰木莲种源幼苗叶片性状表型多样性特征[22];在经典植物分类学中,植物的花、果实、种子性状是最为重视的植物形态学性状[23],果实和种子一样都是物种繁殖系统的重要特征,是在强大的选择压力下表现出较大适应性且受物种遗传控制较强,所以深入开展有关方面的研究,对于揭示物种系统发育、阐明物种濒危机制同样具有重要参考价值。本研究以贵州省分布的西康玉兰野生群落为对象,研究赫章县、盘州市、水城县等4个野生居群的果实主要特征性状及其表型多样性特征,以期揭示不同种源分布地的种实性状特征及其不同个体间的适应与变异程度,为该种种质资源的保存利用和遗传改良提供理论依据。
1 材料与方法
1.1 采样地点
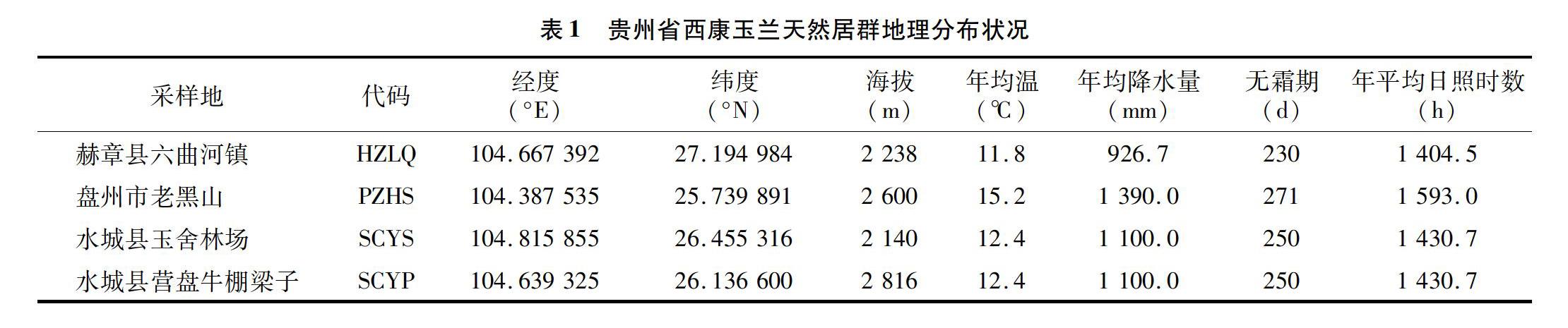
2017年9月选择贵州省西康玉兰典型分布区(贵州省赫章县、盘州市、水城县等地)进行果实采样,研究区的基本情况见表1。
1.2 表型性状的选取及测定
树木果实的形态是树木较稳定的性状之一,也是树木分类以及遗传研究的重要指标[12]。本研究选择遗传性状较为稳定且易于获取的表型性状,具体包括果质量、果长、果宽、果厚、果径、果形态指数、果体积指数、小蓇葖果数、种子数、种实率、果壳质量、去皮前种子质量、去皮后种子质量等共计13个果实表型性状。聚合蓇葖果的长、宽通过游标卡尺进行测量,精度为0.01 mm;果或壳质量通过电子天平测定,精度为0.01 g,每单株随机对100 粒种子进行称量,并换算成千粒质量,重复5次,取平均值,千粒质量测量精度为0.001 g。
为了便于简化和分析不同果实表型性状特征,本研究定义了西康玉兰果实的外在表型性状,指以聚合蓇葖果为研究单元,包含了聚合蓇葖果外在形态结构(果长、果宽、果厚、果径、果形态指数、果体积指数6个性状)和质量性状指标(果质量、果壳质量2个性状),简称外表型性状;内表型性状是指以解剖后的聚合蓇葖果为研究单元,包括了果实内部的数量特征(种子数、小蓇葖果数量2个性状)和质量性状指标(种实率、去皮前种子质量、去皮后种子质量、种子单粒质量4个性状)(表2)。
1.3 数据分析
分别计算果实表型性状的平均值、标准差、变异系数(CV),其中变异系数(CV)=标准差/平均值,利用变异系数(CV)表征性状变异的离散程度[24]。采用Shannon-Wiener 多样性指数[25]评定西康玉兰不同地理种源的果实形态变异频率差异及变异程度,是在计算形态频率分布和多样性指数时,对原始数值性状分级采集数据,数值性状的划级方法为:先计算所有西康玉兰参试材料各性状的总体平均值(M)和标准差(σ),再划分为10级,从第1级[Mi<(M-2σ)]到第10级Mi>(M+2σ)],每0.5σ为1级,每级的相对频率用于计算多样性指数。多样性指数公式为:H=-∑ni=1Piln Pi,式中:H为Shannon-Wiener多样性指数,Pi为西康玉兰果实某个性状第i级别内的材料数占总数的比例。
采用SPASS 21.0和Excel 2013对获得的数据进行方差分析,用LSD多重比较方法进行多重比较,并对西康玉兰果实各性状间的相关性进行分析。
2 结果与分析
2.1 不同居群果实性状指标的特征
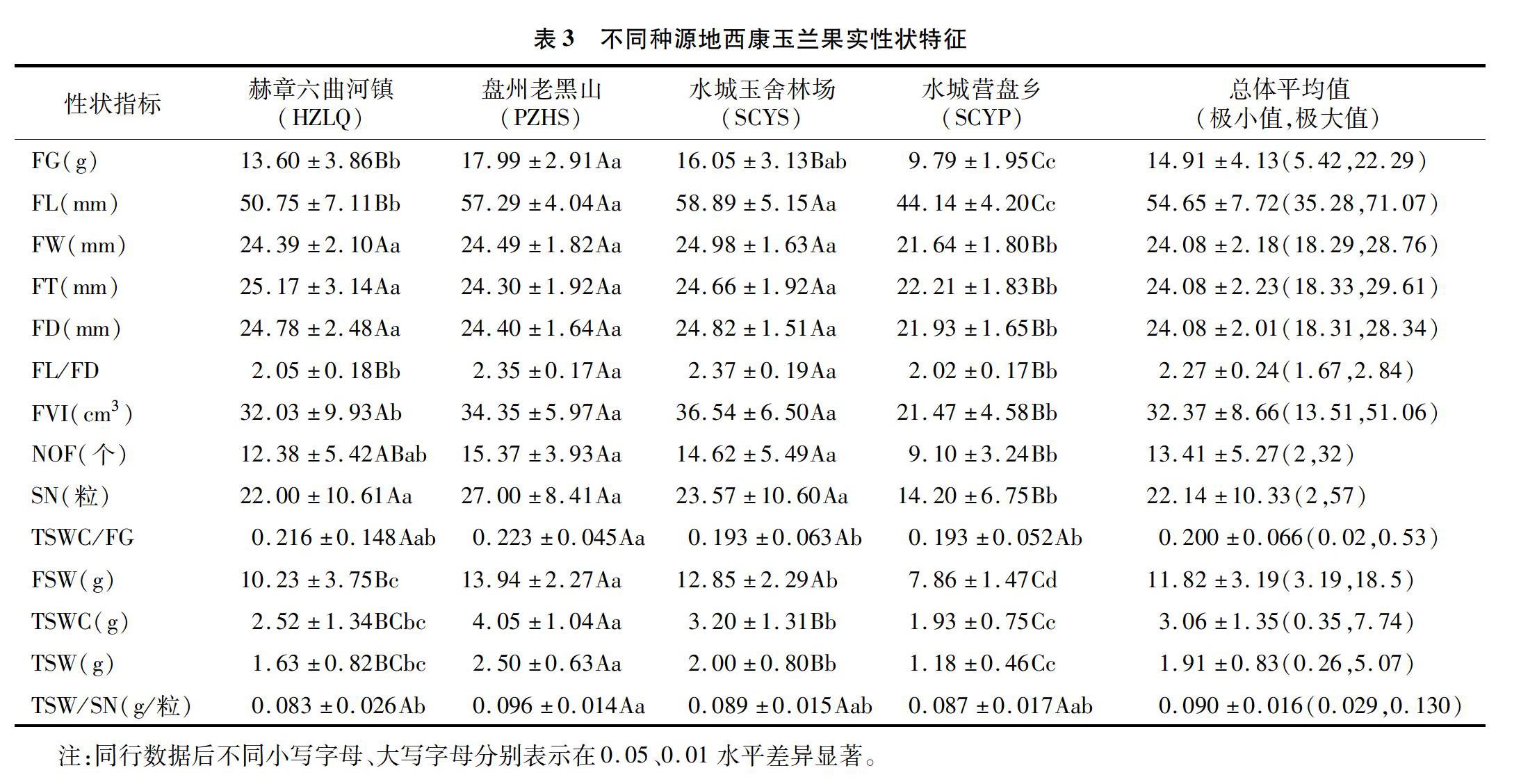
如表3所示,同种果实表型性状在不同种源地间、不同果实表型性状在各种源地的大小表现各异。其中,果厚FT平均值为24.08 mm,不同种源地大小依次为赫章六曲河镇>水城玉舍林场>盘州老黑山>水城营盘乡;果径FD平均值为 24.08 mm,不同种源地大小依次为水城玉舍林场>赫章六曲河镇>盘州老黑山>水城营盘乡;果長FL平均值为54.65 mm,果宽FW平均值为 24.08 mm,果型指数FL/FD平均值为2.27,果实体积指数FVI平均值为32.37 cm3,不同种源地大小均依次为水城玉舍林场>盘州老黑山>赫章六曲河镇>水城营盘乡;果质量FG平均值为14.91 g,小蓇葖果数NOF平均值为13.41个,种子数SN平均值为22.14粒,果壳质量FSW平均值为11.82 g,去皮前种子质量TSWC平均值为3.06 g,去皮后种子质量TSW平均值为1.91 g,不同种源地大小均依次为盘州老黑山>水城玉舍林场>赫章六曲河镇>水城营盘乡;单粒种子质量TSW/SN平均值为0.090 g,不同种源地大小依次为盘州老黑山>水城玉舍林场>水城营盘乡>赫章六曲河镇;种实率TSWC/FG平均值为0.20%。
2.2 西康玉兰果实性状的变异特征
变异系数是统计学上表示变异程度的重要指标,可以反映性状值的离散特征,数值越大说明性状的离散度越大。由表4可知,西康玉兰果实各性状在种群内个体间的变异大小因性状而异,种群内各性状的平均变异系数为0.083~0.467,14个表型性状的平均变异系数大小依次为果实种子数量SN(0.467)、去皮前种子质量TSWC(0.441)、去皮后种子质量TSW(0.435)、小蓇葖果数量NOF(0393)、种实率TSWC/FG(0.350)、果质量FG(0277)、果壳质量FSW(0.270)、果体积指数FVI(0268)、种子单粒质量TSW/SN(0.179)、果长FL(0141)、果型指数FL/FD(0.106)、果厚FT(0093)、果宽FW(0.091)、果径FD(0.083)。通过对各群体种实表型性状变异系数的均值比较可知,盘州老黑山种源的平均变异数最低,赫章六曲河镇种源的平均变异系数最高,不同种源果实性状的平均变异系数依次为赫章六曲河镇(0.318)>总体(0257)>水城营盘乡(0.221)>水城玉舍林场(0217)>盘州老黑山(0.165),表明西康玉兰果实外表型性状和内表型性状变异不具有同步性,而同一性状在不同群体内的变异幅度的差异性,在一定程度上表明西康玉兰生长环境异质性可能是导致群体果实表型变异的主要原因之一。西康玉兰果实为聚合蓇葖果,统计分析8个果实的外表型性状的平均变异系数(0.166)低于其余6个果实内表型性状的变异系数(0.378),不同种源地西康玉兰果实外表型性状平均变异系数均低于果实内表型性状,表明西康玉兰果实外表型性状的稳定性高于果实内表型性状。
2.3 果实性状指标间及其与环境因子间的相关性
采用Pearson相关性分析计算14个性状之间的相关性系数,结果如表5所示,果质量FG、果径FD、果厚FT、果长FL、果宽FW 5个性状指标间均呈极显著正相关(P<0.01)。果质量FG除与单粒种子质量 TSW/SN、TSWC/FG呈负相关外,与其余各项性状指标均呈极显著正相关(P<0.01);果长FL除与单粒种子质量TSW/SN呈正相关外,与其余各项性状指标均呈极显著正相关(P<0.01);果厚FT除与果型指数FL/FD、种实率TSWC/FG呈正相关,与单粒种子质量TSW/SN呈负相关外,与其余各项指标均呈极显著正相关(P<0.01)。果径FD除与单粒种子质量TSW/SN呈负相关、与种实率TSWC/FG呈正相关、与果型指数FD/FL呈显著正相关(P<005),与其余各项性状指标呈极显著正相关(P<0.01)。果型指数FL/FD除与种实率TSWC/FG呈负相关、与单粒种子质量TSW/SN呈显著正相关(P<0.05)外,与其余各项性状指标均呈极显著正
相关(P<0.01);果实体积指数FVI除与种实率TSWC/FG、单粒种子质量TSW/SN呈正相关外,与其余各项性状指标均呈极显著正相关(P<0.01)。
由表6可知,西康玉兰外表型性状与经度地理生态因子主要表现为正相关(仅与果实质量FG、果壳质量FSW呈负相关,但未达显著水平),而与果实内表型性状均表现为负相关,其中与种实率TSWC/FG(r=0.175)、去皮前种子质量TSWC(r=0198)、去皮后种子质量TSW(r=0.187)的相关性达显著水平(P<0.05);纬度地理生态因子除与果实外表型性状果长FL、果宽FW、果厚FT、果径FD、果实体积指数FVI呈正相关外,其余均表现为负相关,其中与果厚FT、果径FD的相关性分别达到显著水平(P<0.05),与果实内表型性状TSWC、TSW、TSW/SN均呈显著负相关(P<0.05)。海拔环境生态因子与果实8个外表型性状均呈极显著负相关(P<0.01),与果实内表型性状小蓇葖果数NOF(r=0.304)和种子数SN(r=0.239)呈极显著负相关(P<0.01),与去种皮前质量TSWC(r=0.191)和去种皮后质量TSW(r=0.213)呈显著负相关(P<0.05),与种实率TSWC/FG(r=0060)和单粒种子质量TSW/SN(r=0.070)呈正相关,但未达显著水平。年均温、年均降水量、年平均日照时数3个气候生态因子与各果实性状指标间均呈正相关,其中与果实质量FG、果型指数FL/FD、果壳质量FSW、种子数SN、去种皮前质量TSWC和去种皮后质量TSW呈极显著正相关(P<0.01),与果长FL、小蓇葖果数NOF、单粒种子质量TSW/SN呈显著正相关(P<0.05)。综上所述,各地理生态因子中,海拔、年均温与多数果实性状间呈显著相关关系,两者对西康玉兰果实内外表型性状的影响较大,而果实内外表型性状与地理生态因子间不同的相关性,应是果实内外表型性状对环境条件适应的外在表现。
2.4 果实性状多样性指数分析
参照苏应雄等的方法[14]采用Shannon-Weaver信息指数(H)计算西康玉兰4个种源地不同果实性状表型多样性指数,由表7可以看出,14个果实性状中,平均多样性指数介于1.538~1.827,果厚FT的平均多样性指数最大,为1.827,果质量FG的最小,为1.538,说明群体水平上,西康玉兰在果厚FT性状表型多样性高于其余果实性状,果质量FG性状表型多样性低于其余果实性状。不同种源地各性状表型多样性显示,水城玉舍林场(SCYS)的果质量FG(1.704)、果厚FT(1.894)、果型指数FL/FD(1.824)、果体积指数FVI(1.775)、小蓇葖果数NOF(1.973)、种子数SN(1.981)、种实率TSWC/FG(1.726)、果壳质量FSW(1.784)、去种皮前种子质量TSWC(1.961)、去种皮后种子质量TSW(1926)、单粒种子质量TSW/SN(1.936)等果实性状表型多样性指数最高,说明该种源地的西康玉兰果实性状变异较为丰富;赫章六曲河镇(HZLQ)种源的果实表型多样性指数的果径FD、果宽FW、果长FL等3个性状取得最大值,说明该种源的这3个性状表型变异较为丰富。群体的果实外表型性状多样性指数(1.674)稍高于内表型性状多样性指数(1671),不同种源地的果实内、外表型性状多样性指数不同,其中赫章六曲河镇(HZLQ)、盘州老黑山(PZHS)种源地的果实外表型性状多样性指数均分别高于果实内表型性状多样性指数;水城玉舍林场(SCYS)、水城营盘乡(SCYP)种源地的则相反。
3 讨论与结论
对西康玉兰4个种源地14个果实表型多样性研究表明,西康玉兰果实表型性状在群体间、群体内变异都较为丰富,这些变异多是由于遗传和环境因素共同作用的结果,群体间、群体内多层次的变异为西康玉兰优良种质资源保育和利用提供了物质基础。
同类果实表型性状在不同种源地间、不同果实表型性状在各种源地的表现各异。不同果实表型性状间的相关性也不同,其中果质量FG、果径FD、果厚FT、果长FL、果宽FW 5个性状指标间均呈极显著正相关(P<0.01)。果质量FG除与单粒种子质量 TSW/SN呈负相关外,与其余各项性状指标均呈极显著正相关(P<0.01);果型指数FL/FD除与种实率TSWC/FG呈负相关、与单粒种子质量TSW/SN呈显著正相关(P<0.05)外,与其余各项性状指标均呈极显著正相关(P<0.01)。
海拔、年均温与多数果实性状间呈显著相关关系,说明它们对西康玉兰果实内外表型性状的影响较大,而果实内外表型性状与地理生态因子间不同的相关性,应是果实内外表型性状对环境条件适应的外在表现。
西康玉兰14个果实性状的平均多样性指数介于1.538~1.827;除果径FD、果宽FW、果长FL等3個果实性状表型变异在赫章六曲河镇(HZLQ)种源地最为丰富外,其余11个果实性状均在水城玉舍林场(SCYS)种源地最为丰富。群体的果实外表型性状多样性指数(1.674)稍高于内表型性状多样性指数(1.671)。不同种源地的果实内、外表型性状多样性指数不同,其中赫章六曲河镇(HZLQ)、盘州老黑山(PZHS)种源地的果实外表型性状多样性指数均分别高于果实内表型性状多样性指数;水城玉舍林场(SCYS)、水城营盘乡(SCYP)种源地的则相反。
植物的表型特征是其长期进化过程中遗传与环境因素综合作用的结果[16],同时也是物种适应性进化的外在表现形式,植物物种的表型性状既有变异性又有稳定性,其与该物种本身的遗传特性及其所处的环境密切相关,因此,通过植物表型多样性的分析可以揭示群体的遗传规律与变异大小。贵州省是西康玉兰的重要分布区之一[2-3],其果实丰富的表型变异为其资源良种化和产业化提供了广阔的前景,但目前针对该树种的种质资源评价、回归引种、良种选育、栽培管理等更进一步的研究尚缺乏系统性,本研究结果对西康玉兰种质资源收集、遗传多样性保护以及遗传改良工作具有一定的指导意义。
参考文献:
[1]汪 松,解 焱. 中国物种红色名录[M]. 北京:高等教育出版社,2004:327.
[2]袁丛军,杨焱冰,戴晓勇,等. 贵州木兰科植物种类及资源分布格局特征[J]. 西部林业科学,2017,46(3):68-75.
[3]郭 应,张建华,路元礼,等. 贵州盘县西康玉兰调查研究[J]. 安徽农业科学,2014,42(24):8229-8231.
[4]吴守清,张廷忠. 西康玉兰种子育苗试验初报[J]. 贵州林业科技,1998,26(1):42-43,50.
[5]韩春艳,龙春林. 濒危植物西康玉兰种子休眠、萌发及贮藏特性[J]. 云南植物研究,2010,32(1):47-52.
[6]姚淑均,张守攻,王军辉,等. 滇楸花部性状的表型多样性研究[J]. 中南林业科技大学学报,2013,33(10):19-24.
[7]芦 娟,柴春山,蔡国军,等. 甘肃定西文冠果花的表型多样性研究[J]. 林业资源管理,2011,(6):49-53.
[8]许玉兰,蔡年辉,陈 诗,等. 云南松天然群体球果表型变异研究[J]. 种子,2018,37(1):62-67.
[9]何庆海,杨少宗,李因刚,等. 枫香树种群种子与果实表型性状变异分析[J]. 植物生态学报. 2018,42(7):62-67.
[10]乔 谦,丰 震,任红剑,等. 元宝枫天然种群种实表型多样性[J]. 生态学杂志,2017,36(10):2707-2715.
[11]田胜平,汪阳东,陈益存,等. 山苍子天然种群叶片和种实性状的表型多样性[J]. 生态学杂志,2012,31(7):1665-1672.
[12]张 莹,曹玉芬,霍宏亮,等. 基于枝条和叶片表型性状的梨种质资源多样性[J]. 中国农业科学,2018,51(17):3353-3369.
[13]王 丹,庞春华,高亚卉,等. 茶条槭不同海拔种群的表型多样性[J]. 云南植物研究,2010,32(2):117-125.
[14]苏应雄,张 雪,王文礼,等. 红棕杜鹃不同海拔种群的表型多样性研究[J]. 西北植物学报,2017,37(2):356-362.
[15]成向荣,邢文黎,苑海静,等. 披针叶茴香对变化光环境的表型可塑性[J]. 生态学报,2019,39(6):1935-1944.
[16]周 济,Tardieu F,Pridmore T,等. 植物表型组学:发展、现状与挑战[J]. 南京农业大学学报,2018,41(4):580-588.
[17]Tester M,Langridge P. Breeding technologies to increase crop production in a changing world[J]. Science,2010,327(5967):818-822.
[18]张俪文,韩广轩. 植物遗传多样性与生态系统功能关系的研究进展[J]. 植物生态学报,2018,42(10):977-989.
[19]阎爱民,陈文新. 苜蓿、草木樨、锦鸡儿根瘤菌的表型多样性分析[J]. 生物多样性,1999,7(2):112-118.
[20]刘艳萍,马群智,刘红贤,等. 广玉兰优树果实和种子的表型多样性[J]. 上海农业学报,2014,30(2):65-67.
[21]宋 杰,李世峰,刘丽娜,等. 云南含笑天然居群的表型多样性分析[J]. 西北植物学报,2013,33(2):272-279.
[22]文珊娜,仲崇禄,姜清彬,等. 灰木莲种源幼苗叶片性状表型多样性分析[J]. 植物研究,2017,37(2):288-297.
[23]司马永康,陆树刚,韩明跃,等. 木兰科系统分类学研究动态[J]. 西部林业科学,2012,41(1):116-127.
[24]葛 颂,王明庥,陈岳武. 用同工酶研究马尾松群体的遗传结构[J]. 林业科学,1988,24(4):399-409.
[25]姚小华,黄 勇,任华东,等. 小果油茶不同居群葉表型性状频率分布及多样性指数分析[J]. 西南林业大学学报,2014,34(5):47-52.
