利用SSR标记筛选水稻富γ-氨基丁酸后代材料
2020-06-01王康恺赵娜马嘉欣王迎超田玲杨治伟洪缘缘田蕾张银霞杨淑琴李培富
王康恺 赵娜 马嘉欣 王迎超 田玲 杨治伟 洪缘缘 田蕾 张银霞 杨淑琴 李培富

摘要:以高粱稻-1和宁农黑粳的杂交F2代和F4代为试验材料,在实验室前期已获得的稻米籽粒中控制γ-氨基丁酸(γ-aminobutyric acid,GABA)含量的3个数量性状基因座(quantitative trait locus,QTL)位点基础上,进一步利用简单重复序列(simple sequence repeats,SSR)标记对其进行基因型检测,获得了不同基因组合类型,在此基础上利用高效液相色谱法,测定各基因组合类型籽粒中GABA含量,探讨GABA含量与其类型间的关系。结果表明,在F2代和F4代的所有组合类型中,代表高含量纯合基因组合类型(BBA组合)中GABA含量的检测结果均值最高,代表低含量纯合基因组合类型(AAB组合)中GABA含量的检测结果均值最低,其检测结果与基因型表现一致。显著性分析表明,GABA含量与不同基因组合类型之间存在极显著相关性。同时还表明,相比F2代,F4代的BBA组合中GABA的平均含量增高,AAB组合中GABA的平均含量降低,且变异程度均有降低,特别是低含量纯合基因组合类型材料的符合度从53%上升到75%,其结果表明,低含量纯合基因组合类型材料有趋于稳定的趋势。试验结果可应用于利用分子标记辅助选择富γ-氨基丁酸杂交后代材料的研究中。
关键词:水稻;γ-氨基丁酸;高效液相色谱;糙米;籽粒;基因型组合;SSR标记
中图分类号: S511.032文献标志码: A
文章编号:1002-1302(2020)08-0078-07
收稿日期:2019-03-18
基金项目:国家自然科学基金(项目编号:31360324、31760374、31401361);宁夏农业育种专项课题(编号:2018NYYZ0302)。
作者简介:王康恺(1991—),男,陕西华阴人,硕士研究生,主要研究方向为水稻遗传育种。E-mail:1033678950@qq.com。
通信作者:李培富,博士,教授,主要从事水稻遗传育种研究。E-mail:peifμLi@163.com。
γ-氨基丁酸(γ-aminobutyric acid,GABA)又称γ-氨酪酸,是一种天然存在的非蛋白组成性氨基酸[1]。γ-氨基丁酸不仅在植物组织中存在,同时也在哺乳动物和人类的中枢神经系统中存在,是中枢神经系统中很重要的抑制性神经递质,参与人体内多项代谢及生理活动,具有降血压、健脑、活化肝肾及调节免疫等功能,被誉为“大脑的天然镇静剂”[2]。为了实现“药食同源”,调节人体生理功能,尤其是对GABA摄入量有特殊要求的人群而言,研究并开发富含GABA的水稻新品种显得非常必要。
目前,国内外对于γ-氨基丁酸的研究主要集中在其生理功能[3-5]、测定方法[6-7]以及发芽糙米工艺优化[8]等方面,对富含GABA新品种培育的研究相对较少。目前,张标金通过测定巨胚稻和常规稻糙米中的GABA含量发现,巨胚稻糙米中GABA的平均含量显著高于常规稻,并从中筛选出富含GABA的巨胚稻GE091[6];Maeda等以金南风(Kinmaze)的巨胚突变体作为杂交亲本,培育出了巨胚水稻新品种Haiminori,Haiminori籽粒中的GABA含量比日本晴、越光等普通水稻品种籽粒中的γ-氨基丁酸含量高3~4倍[9];日本利用九州大学的变异巨胚系统EM40和中国农试高产品种明光诺里杂交,选育出富γ-氨基丁酸水稻新品种海米诺里[10],其胚芽内的GABA含量比普通品种高约4倍。
本试验通过检测杂交F2代182份材料和杂交F4代109份材料的基因型,将所有材料按基因型组合进行分类,利用高效液相色谱法,测定各基因型组合籽粒中γ-氨基丁酸含量,分析不同类型材料γ-氨基丁酸含量与基因型的相关性,旨在为利用分子标记辅助选择培育富γ-氨基丁酸新品种提供参考。
1 试验材料与方法
1.1 试验材料
1.1.1 供试水稻材料
试验材料由宁夏大学农学院作物遗传育种实验室提供,包括高粱稻-1和宁农黑粳杂交F2,共182份单株材料。试验材料于2017年10月在海南省三亚育种基地种植,采用播后上水的方式,常规田间管理方法,成熟时单株收获。再于2018年5月在宁夏大学水稻育种基地种植,采用旱育秧方式育苗,单株移栽,田间采用常规栽培管理方法,成熟后收获单株材料109份。
1.1.2 试验用相关引物
本试验所用引物来自笔者所在实验室前期构建的简单重复序列(simple sequence repeats,SSR)引物连锁遗传图谱,以GABA含量为10.47 mg/100 g的高粱稻-1作为父本,以GABA含量为5.57 mg/100 g的宁农黑粳作为母本,以杂交F2群体为基础,筛选出贡献率最高的3个数量性状基因座(quantitative trait locus,QTL)位点,再对其进行优化。结果如表1所示,其中qGABA-8-1和qGABA-8-2来自母本宁农黑粳,qGABA-9来自父本高粱稻-1。引物由生工生物工程(上海)股份有限公司北京分公司合成。
1.2 试验方法
1.2.1 籽粒γ-氨基丁酸含量的测定
本试验采用水提法(参照程威威等的提取方法[7,11]并进行优化),提取水稻糙米中的GABA。將收获的182份杂交F2代和109份杂交F4代籽粒于60 ℃烘箱中烘干备用,使用砻谷机脱壳,磨粉机研磨,过80目筛,准确称取糙米粉(1.000 0±0000 2) g,每份材料重复3次,倒入50 mL离心管中,加入15 mL超纯水,混匀,60 ℃、120 r/min水浴振荡浸提90 min,12 000 r/min 离心20 min,将上清液移入圆底烧瓶中,重复上述操作,将2次上清液混合,用旋转蒸发仪在96 ℃下旋转蒸发,最后浓缩到5 mL,过 0.2 μm 有机滤膜,得到糙米中γ-氨基丁酸提取液,然后将提取液进行高效液相色谱(high performance liquid chromatography,HPLC)柱前衍生,使用安捷伦1220高效液相色谱仪进行测定,并绘制标准曲线,计算籽粒γ-氨基丁酸含量。
根据表2中的数据,以γ-氨基丁酸标准品的浓度为横坐标,峰面积为纵坐标,绘制出γ-氨基丁酸浓度与峰面积的标准曲线(图1),拟合标准曲线方程为:y=1 133 314x-1 023.11,相关系数r2=0997 4。
1.2.2 叶片DNA的提取及检测
本试验采用十二烷基硫酸钠(sodium dodecyl sulfate,SDS)法提取叶片中的DNA,以DNA为模板进行PCR扩增,配制20 μL反应体系,其中10×Buffer(含Mg2+)2.0 μL,2.5 μmol/mL dNTP 0.4 μL,引物(2 mmol/L)15 μL,Taq酶0.4 μL,模板DNA 2.0 μL,最后用ddH2O补足20 μL。PCR反应程序:95 ℃ 5 min;95 ℃ 50 s,58 ℃ 50 s,72 ℃ 1 min,35個重复;最后72 ℃延伸10 min。扩增产物用8%的聚丙烯酰胺凝胶(PAGE)电泳检测,银染显色,包胶,并于胶片观察灯下观察,读胶。
1.3 试验数据处理
电泳后,读取杂交后代群体的条带,将与宁农黑粳带型一致的条带记为B,与高粱稻-1带型一致的条带记为A,双亲杂合型条带记为H,缺失记为“-”。在Microsoft Excel表中整理统计4种带型数据,用Microsoft Excel完成试验数据的处理及绘图,并用SPSS19.0分析软件进行方差分析。
2 结果与分析
2.1 杂交后代群体基因型分析
将所用引物合成,按要求稀释,然后进行PCR扩增反应,聚丙烯酰胺凝胶电泳检测,部分结果如图2所示,统计全部结果得出,其中有效条带占全部条带的 94.65%,数据可靠性较高,可用于下一步检测分析。
2.2 杂交后代单株籽粒基因型组合及其对应γ-氨基丁酸含量
对杂交F2代和F4代材料的单株籽粒基因型进行分类,并对其相应GABA含量进行统计,结果如表3、表4所示。由表3可以看出,F2群体中,GABA含量最高为 15.02 mg/100 g,其基因组合类型为BBH,最低为377 mg/100 g,其基因组合类型为AAB,其中纯合基因型组合材料中GABA含量最高为12.15 mg/100 g,对应基因组合类型为BBA;由表4可以看出,F4群体中,GABA含量最高为 12.89 mg/100 g,其基因组合类型为BBB,最低为299 mg/100 g,其基因组合类型为AAB。综上,在F2、F4代群体中,GABA含量最低的材料基因型组合均为AAB,但最高含量来自不同基因型组合。
2.3 杂交后代籽粒中γ-氨基丁酸含量与其基因组合类型的差异显著性分析
由图3可以看出,HHA、BBA基因组合类型的GABA含量高于BBB、HBH、HHH、HAH、HHB、AAA、AAH、HAB、AAB基因组合类型,AAH、HAB、AAB类型含量低于BBH、HHA、BBA类型,且均呈极显著差异。其中HHA、BBA基因组合类型材料的GABA平均含量较高,分别为10.08、9.87 mg/100 g,AAB基因组合类型材料平均含量最低,为 5.98 mg/100 g,前者分别是后者的1.69、1.65倍。从遗传学角度来看,杂合材料存在分离且稳定性不高,所以选择纯合BBA类型进行后代遗传。由图4可以看出,BBA类型GABA含量高于HHB、BBH、AHB、HHH、HBB、HAB、AAB类型,AAB类型含量低于BBA、BBB、BAB、BHH类型,也均呈极显著差异。其中BBA类型材料的GABA平均含量最高,达 10.12 mg/100 g;AAB类型材料平均含量最低,为3.30 mg/100 g;BBA类型材料是AAB的3.07倍。综上,F2、F4代群体中,GABA平均含量最低的均为AAB类型材料,且含量有下降趋势,BBA类型材料从F2到F4代变为最高,且平均含量有所提高。
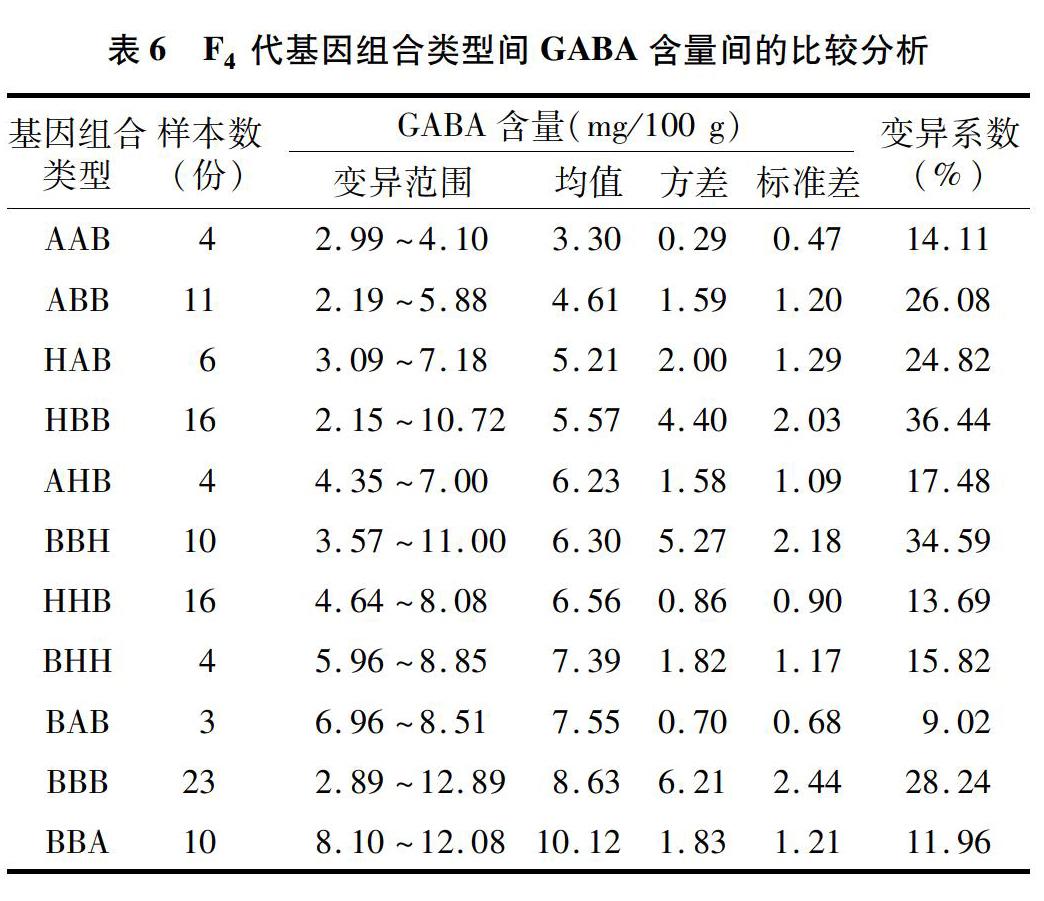
2.4 不同基因组合类型间GABA含量的比较
由表5可以看出,F2代中,纯合AAB基因组合类型材料γ-氨基丁酸含量的平均值为(5.98±1.15) mg/100 g,变异范围为3.77~7.98 mg/100 g,变异系数为1930%;纯合BBA基因组合类型材料γ-氨基丁酸含量的平均值为(9.87±1.24) mg/100 g,变异范围为 8.18~12.15 mg/100 g,变异系数为1256%。由表6可以看出,F4代中,纯合AAB基因组合类型材料γ-氨基丁酸含量的平均值为(3.30±0.47) mg/100 g,变异范围为2.99~4.10 mg/100 g,变异系数为14.11%;纯合BBA基因型材料γ-氨基丁酸含量的平均值为(10.12±121) mg/100 g,变异范围为8.10~12.08 mg/100 g,变异系数为11.96%。综上, 从F2代到F4代,纯合AAB、BBA基因组合类型材料变异系数均有降低,其中AAB降低幅度较大,且均值都有向其最低或最高方向变化的趋势。由表5、表6还可以看出,杂合基因型材料变异系数不稳定。
2.5 纯合高含量与低含量基因组合类型材料符合度的比较
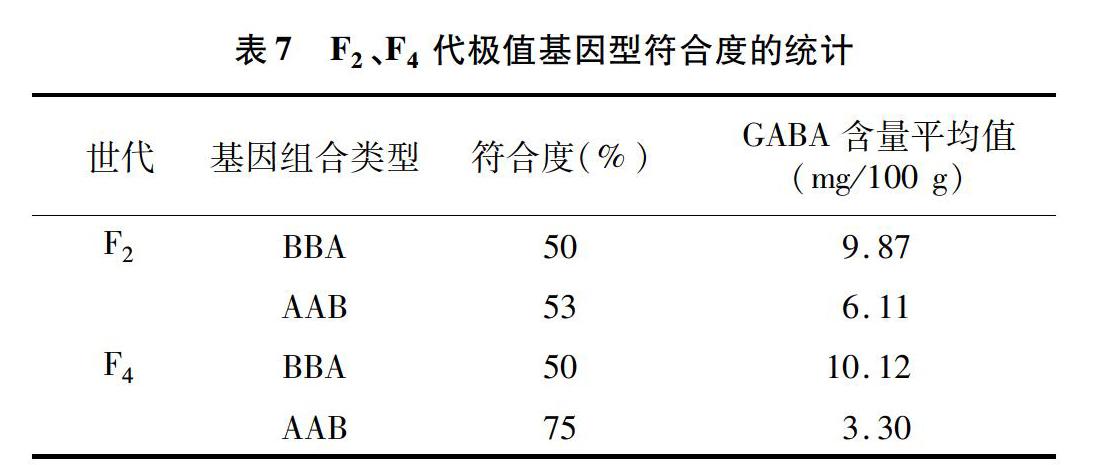
为进一步探究纯合高含量基因组合类型BBA和纯合低含量基因组合类型AAB材料的稳定性,对这2种组合类型的符合度进行了统计。结果(表7)表明,F2、F4代中BBA类型材料的符合度均为50%,AAB类型材料的符合度分别为53%、75%,相比纯合高含量基因组合类型材料,纯合低含量基因组合类型材料的符合度随着世代增加有所增加,说明纯合低含量基因组合类型材料稳定性更高。
3 结论与讨论
3.1 水稻籽粒中γ-氨基丁酸含量测定方法比较
γ-氨基丁酸作为一种新型的功能性因子,其测定方法目前在我国还没有明确统一标准[12-13]。比色法和高效液相色谱法(HPLC)是这些年比较常用的2种测定γ-氨基丁酸含量的方法,但是均有优缺点。比色法利用游离氨基酸与苯酚和次氯酸钠反应,生成有色物质,进行比色测定。这一方法操作简便,设备简单,适合大规模测定。但是水稻糙米中的GABA含量普遍较低,所以在对其进行检测时,其他物质产生的颜色会对分光光度计显色反应造成影响,使其准确度降低。HPLC检测的原理是γ-氨基丁酸与一些特殊的化学试剂发生柱前衍生反应后,生成一种能被紫外-可见光检测到的稳定荧光物。这一方法分离度较高、重复性较好、精密度较高、结果比较准确,但试剂昂贵,程序也较为繁琐。程威威等对比分析了邻苯二甲醛(OPA)、丹磺(Dansyl-Cl)、6-氨基喹啉基-N-羟基琥珀酰亚氨基甲酸酯(AQC)等作为柱前衍生试剂,检测发芽糙米中的γ-氨基丁酸含量结果,发现OPA虽然衍生时间比较短,但其衍生产物较不稳定,易分解[7,14];Dansyl-Cl衍生过程比较复杂,不仅需要避光保存,而且其衍生产物也相对较不稳定[15];而AQC作为一种较为新型的柱前衍生试剂,衍生反应时不仅反应迅速,灵敏度较高,而且衍生产物也较为稳定[16]。唐涛等在研究中均证实了AQC的优越性[17-19]。因此,本试验采用高效液相色谱法,并用AQC作柱前衍生,检测籽粒中γ-氨基丁酸含量。
3.2 水稻籽粒γ-氨基丁酸含量与基因型的相关关系
王迎超研究发现,高粱稻-1与宁农黑粳杂交组合F1代籽粒γ-氨基丁酸含量介于双亲之间,F2代籽粒γ-氨基丁酸含量呈偏正态分布,并出现超亲现象[2]。本试验在此基础上,依据3个QTL位点,对杂交F2代和F4代进行基因型组合分类,具体分析结果表明,不同基因组合类型与γ-氨基丁酸含量之间呈极显著相关,代表高含量纯合基因型(BBA组合)中GABA的检测结果均值最高,代表低含量纯合基因型(AAB组合)中GABA的检测结果均值最低,其检测结果与基因型表现一致。显著性分析结果表明,GABA含量与其基因组合类型之间存在极显著相关性。F2代BBA类型材料的GABA平均含量 (9.87 mg/100 g) 是AAB类型材料平均含量(5.97 mg/100 g)的1.65倍,F4代BBA类型材料平均含量(10.12 mg/100 g)是AAB类型材料GABA平均含量(3.30 mg/100 g)的3.07倍。结果表明,相比于F2代,F4代的BBA组合中GABA的平均含量增高,且F4代的AAB组合中GABA的平均含量降低,变异程度也均有所降低,特别是低含量纯合基因型材料符合度从53%上升到75%,其结果表明,低含量材料γ-氨基丁酸含量趋于稳定程度较高。
3.3 富γ-氨基丁酸水稻品种的选育
本试验在王迎超的研究基础[2]上进行延伸,利用分子标记辅助技术筛选富γ-氨基丁酸的水稻新材料,不仅可以缩短育种年限,也能快速选择出富γ-氨基丁酸的水稻新品种。有关水稻籽粒中控制γ-氨基丁酸含量的遗传学研究起步较晚,目前,籽粒中控制γ-氨基丁酸含量的QTL位点并没有得到精细定位。本试验结果证明,控制GABA基因定位的准确性,可为分子标记辅助选择富含γ-氨基丁酸的水稻新品种以及控制γ-氨基丁酸含量的候选基因筛选奠定基础。
参考文献:
[1]王迎超,王全兴,王 浩,等. 富γ-氨基丁酸水稻种质筛选及与籽粒性状的相关性分析[J]. 植物遗传资源学报,2016,17(6):1116-1122.
[2]王迎超. 水稻γ-氨基丁酸含量的遗传分析及QTL定位[D]. 银川:宁夏大学,2016.
[3]张祥喜,袁林峰,刘 凯,等. 富含γ-氨基丁酸(GABA)的巨胚功能稻研究进展[J]. 江西农业学报,2007,19(1):36-39.
[4]张琳琳,舒小丽,卢怀江,等. 富含γ-氨基丁酸降压功能稻米研究进展[J]. 核农学报,2006,20(3):218-220.
[5]王 辉,项丽丽. γ-氨基丁酸(GABA)的功能性及在食品中的应用[J]. 食品工業,2013,6(34):186-189.
[6]张标金. 用基于邻苯二甲醛柱前衍生的高效液相色谱法测定稻米的γ-氨基丁酸含量[J]. 江西农业学报,2013,25(11):105-107.
[7]程威威. HPLC法测定发芽糙米中γ-氨基丁酸中不同衍生方式的比较研究[J]. 中国农学通报,2014,30(9):279-284.
[8]李常钰,王超超,杨慧萍. 发芽糙米中γ-氨基丁酸的富集与测定[J]. 粮食与饲料工业,2011(2):4-7.
[9]Maeda H,Nemoto H,Iida S,et al. A new rice variety with giant embryos,“Haminori”[J]. Breeding Sci,2001,51:211-213.
[10]姜 宾. 日本育出巨大胚水稻新品种[J]. 天津农林科技,2001(4):45.
[11]房克敏,李再贵,袁汉成,等. HPLC法测定发芽糙米中γ-氨基丁酸含量[J]. 食品科学,2006,27(4):208-211.
[12]田小磊,吴效岚. γ-氨基丁酸在高等植物逆境反应中的作用[J]. 生命科学,2002,14(4):215-219.
[13]徐瑞萍. γ-氨基丁酸在发芽糙米中检测方法与综述[J]. 山东化工,2012,41(11):35-37.
[14]陈 稚. 食品营养强化剂及保健食品中的违禁药物分析方法研究[D]. 长沙:湖南师范大学,2006.
[15]Huang X,Chen J W,He L P,et al. Detection of monoamine neurotransmitters and its metabolites by high performance liquid chromatograph after pre-colum derivatization of dansyl chloride column[J]. Europe Pub Med Central,2012,46(12):51-53.
[16]覃婷婷,白海娇,黄哲苏,等. AQC柱前衍生法在氨基酸分析测定中的应用[J]. 天津药学,2010,22(5):51-53.
[17]唐 涛. 氨基酸柱前衍生化HPLC方法发展及应用[D]. 南京:南京理工大学,2006.
[18]吕莹果,张 晖,孟 祥. 氨基酸分析测定及衍生化方法[J]. 粮食与油脂,2009(7):35-38.
[19]王秀中,王清清,宋海峰. 衍生化技术在氨基酸分析中的应用进展[J]. 药物分析杂志,2010,30(6):1162-1166.
