孤儿核受体Nur77基因多态性与脓毒症易感性的关联研究
2020-06-01尧国胜黄高吴晓文农荣卯黄非凡黄宝雍陆政唐宁廖品琥
尧国胜黄高吴晓文农荣卯黄非凡黄宝雍陆政唐宁廖品琥
(1. 右江民族医学院附属医院,广西 百色 533000;2. 百色市人民医院,广西 百色 533000;3. 广西壮族自治区人民医院,广西 南宁 530000;4. 广西医科大学附属武鸣医院,广西 武鸣 530100;5. 右江民族医学院重症医学教研室,广西 百色 533000)
脓毒症是指宿主对感染产生失控性炎症反应而导致的危及生命的器官功能障碍的一组综合征,其发病率与死亡率均居高不下,且发病机制尚未完全阐明[1]。Nur77是核受体(Nuclear Receptor,NR)超家族成员之一,作为转录因子,是一类早期即刻反应基因,细胞在感受到环境的生理与病理刺激时,该基因将迅速反应,合成相应的蛋白以应对这些刺激。既往研究表明Nur77通过与肿瘤坏死因子受体相关基因6(Tumor Necrosis Factor Receptor-Associated Factor 6,TRAF6)、核因子-κB(Nuclear Factor-κB,NF-κB)等相互作用,在炎症反应中发挥双向调控作用[2]。脓毒症是环境因素与遗传因素共同作用的多基因性疾病,基因多态性,尤其是单核苷酸多态性(single nucleotide polymorphisms,SNPs)是决定个体脓毒症易感性、表型、治疗反应和临床转归的重要因素[3]。现国内外有报道Nur77基因多态性与糖尿病肾病[4]、运动障碍[5]等疾病相关,但是,对于Nur77基因多态性与脓毒症的关系尚缺乏研究。本文拟通过研究广西人群中Nur77基因相关位点的等位基因及其基因型的分布情况,进一步探讨Nur77基因单核苷酸多态性与脓毒症易感性的关联,以期阐明Nur77基因多态性在脓毒症发生、发展中的作用。
1 对象与方法
1.1 实验对象 选取2018年6月—2019年7月右江民族医学院附属医院、百色市人民医院、广西壮族自治区人民医院、广西医科大学附属武鸣医院的重症医学科收治的脓毒症患者共207例作为脓毒症组,男性150例,女性57例,平均年龄(58.03±13.34)岁。纳入标准:①18岁≤年龄≤80岁;②相互间无生物学亲缘关系;③符合脓毒症和脓毒性休克第三版国际共识定义[1]诊断标准。排除标准:①年龄<18岁或>80岁;②心脏停搏者; ③非感染因素所导致的炎性病变,如出血性休克、免疫介导的器官损伤等;④长期或近期接受过免疫抑制剂或糖皮质激素治疗;⑤其他特殊病例,如肿瘤、人类获得性免疫缺陷病毒感染、自身免疫功能紊乱的病例;⑥近期接受化疗或放疗的病人。随机选取本院同期无亲缘关系的健康体检者210例作为对照组,其中男性158例,女性52例,平均年龄(56.17±12.38)岁。两组年龄和性别比较差异无统计学意义(P>0.05),具有可比性。本研究经右江民族医学院附属医院医学伦理委员会批准,征得患者及其家属同意,并签署知情同意书。
1.2 实验方法
1.2.1 基因组DNA提取 用EDTA-K2抗凝采血管采集入住ICU 24h内的脓毒症患者外周静脉血3ml,对照组受试者采集清晨空腹外周静脉血。使用天根生化[(北京)科技有限公司的血液基因组DNA提取试剂盒(货号:DP348-03)]并采用离心柱法严格按说明操作提取全基因组DNA。
1.2.2 PCR扩增 根据SNP位点序列信息,引物用在线Primer3设计(见表1),由上海生工公司合成,扩增PCR反应体系(20 μl)包含1× GC-I buffer(Takara)、3.0 mmol/L Mg2+、0.3 mmol/L dNTP,1 μmol/L HotStarTaq polymerase (Qiagen) 、1 μl样本DNA和1 μl多重PCR引物。扩增参数:第一步,95℃预变性2 min;第二步,94℃变性20 s,65℃退火40 s,每个循环降低0.5℃,72℃延伸1.5 min,共11个循环;第三步,94℃变性20 s,59℃退火30 s,72℃延伸1.5 min,共24个循环;第四步,72℃延伸2 min,产物保存于4℃备用。
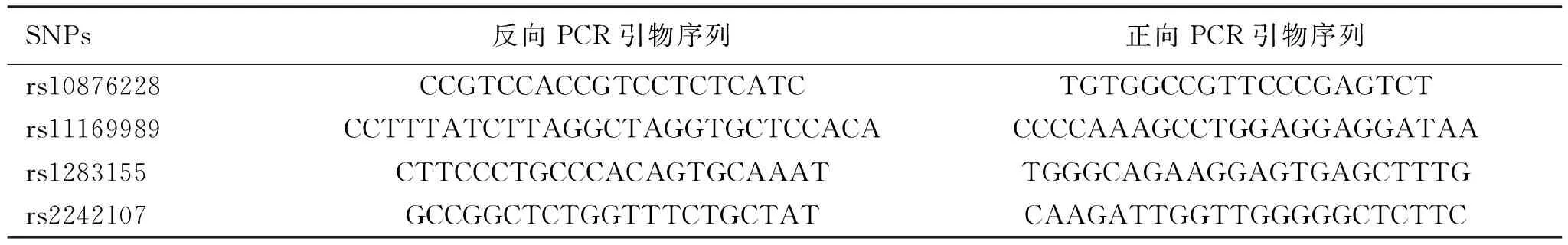
表1 Nur77基因位点PCR引物序列
注:SNP:Single Nucleotide Polymorphism,单核苷酸多态性;PCR:polymerase chain reaction,聚合酶链反应
1.2.3 PCR产物纯化 在10 μl PCR产物中加入5 U虾碱酶(Promega,货号:M9910)和2 U外切酶I(Epicentre,货号:UEM04100),37℃温浴1 h后75℃灭活15 min。
1.2.4 SNaPshot多重单碱基延伸反应 采用SNaPshot Multiplex kit(ABI,货号:4323159)进行延伸反应。10 μl延伸反应体系中包含5 μlSNaPshot Multiplex Ready Mix,2 μl纯化后PCR产物,2 μl超纯水及1 μl延伸引物混合物。反应程序为:第一步,96℃预变性1 min;第二步,96℃变性10 s,55℃退火5 s,60℃延伸30 s,共28个循环;产物保存于4℃备用。
1.2.5 延伸反应产物纯化 将1 U 虾碱酶加入10 μl延伸产物中,37℃温浴1 h,而后75℃灭活15 min。
1.2.6 测序及数据分析 将0.5 μl 纯化延伸产物与9 μl Hi-Di 及0.5 μl Liz120 SIZE STANDARD混匀,于95 ℃变性5 min,在ABI 3730xl测序仪上样检测。所得数据采用GeneMapper 4.1(Applied Biosystems,美国)进行分析。

2 结果
2.1 脓毒症组患者临床特征 本研究纳入的脓毒症患者中,胸腹部为主要感染部位(85.00%),其次为泌尿系、胆道、四肢及颅脑,联合感染者8例。病情发展至脓毒性休克者85 例,见表2。
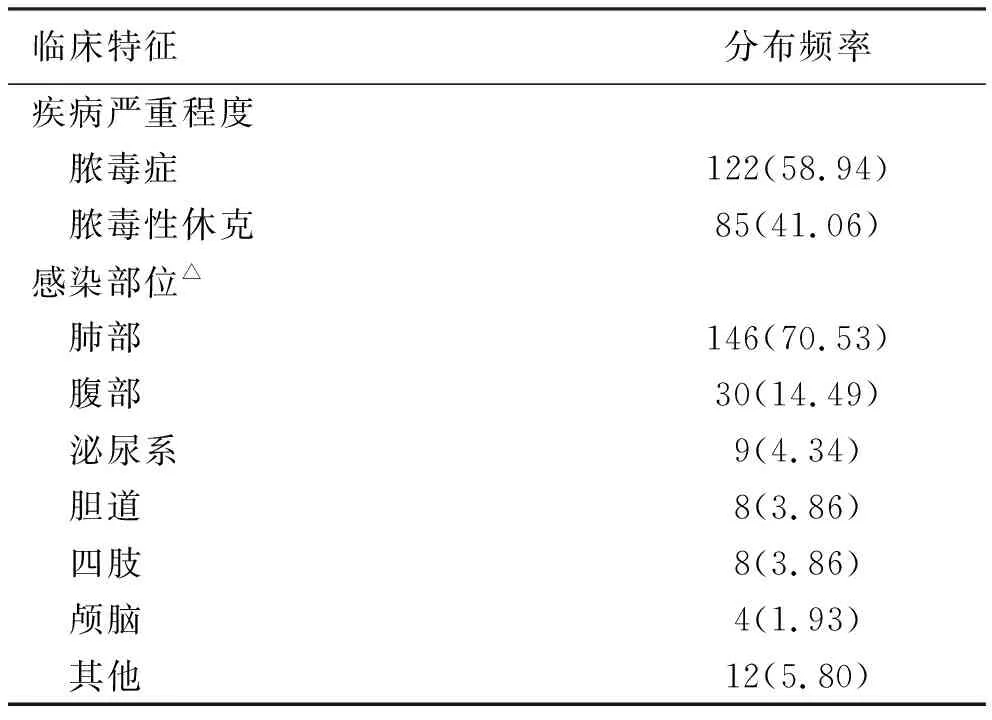
表2 脓毒症患者临床特征
注:①△:多部位感染8例;②表内计数资料数据用[n(%)]表示
2.2 Nur77基因多态性与脓毒症易感性的关联分析 所选4个SNPs在对照组及病例组均满足Hardy-Weinberg平衡定律,提示研究样本具有群体代表性。校正了年龄、性别等混杂因素后,脓毒症组和对照组之间rs10876228、rs11169989、rs1283155、rs2242107各基因型频率在各种遗传模型下差异均无统计学意义(P>0.05),见表3。
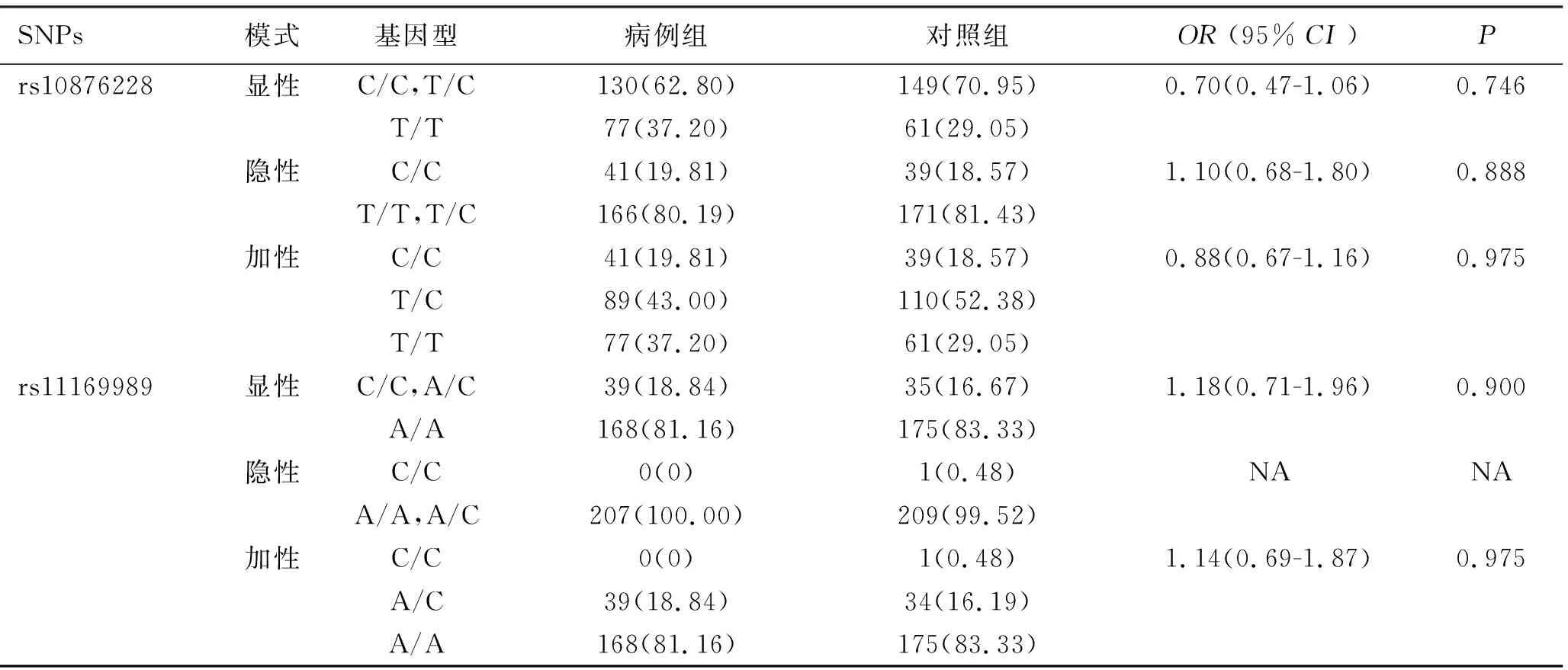
表3 Nur77基因多态性与脓毒症易感性的关联分析
注:①所有OR值均经性别及年龄等因素的校正;②表内计数资料数据用[n(%)]表示;③NA:No Available,因病例组C/C基因型病例数为0,无法进行统计学分析
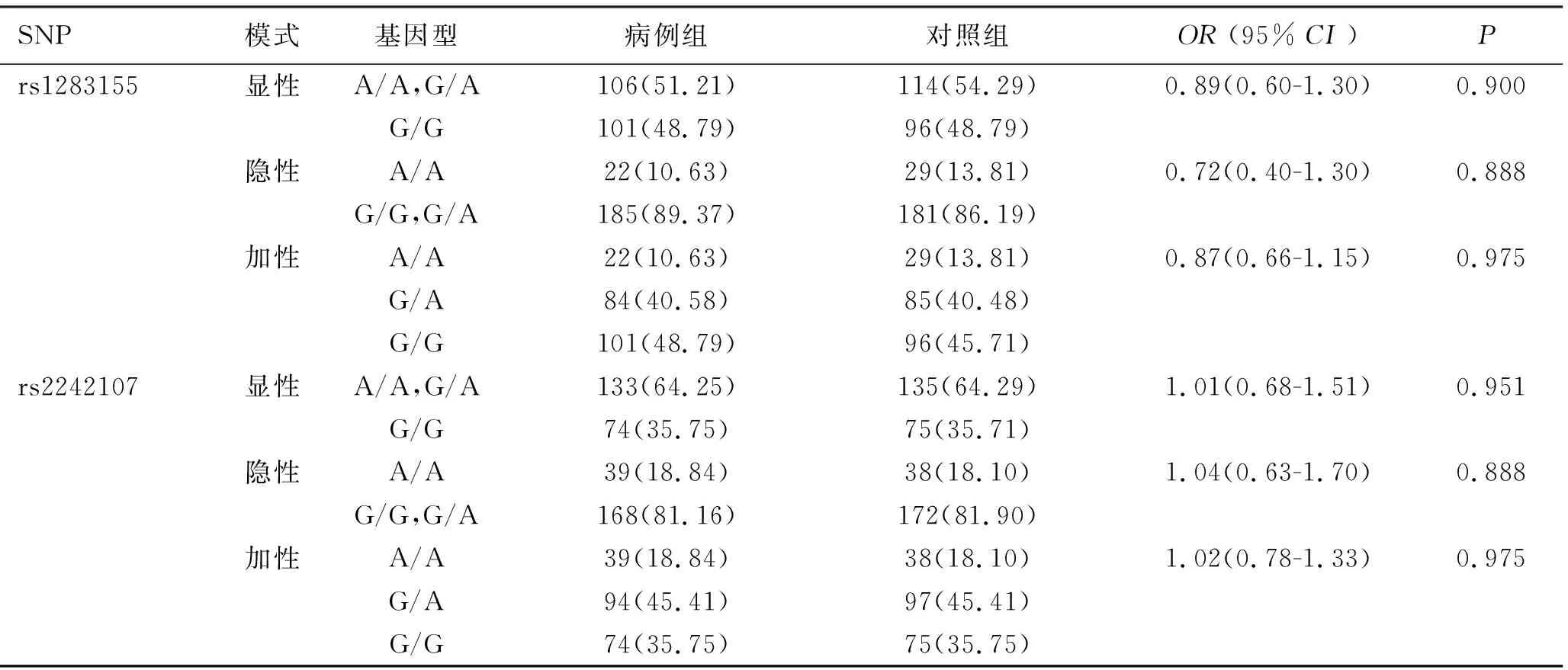
续表3 Nur77基因多态性与脓毒症易感性的关联分析
注:①所有OR值均经性别及年龄等因素的校正;②表内计数资料数据用[n(%)]表示;③NA:No Available,因病例组C/C基因型病例数为0,无法进行统计学分析
3 讨论
脓毒症是宿主对感染的失控反应,可导致危及生命的器官功能障碍[1],是严重威胁人类健康的主要疾病之一,给全球医疗卫生系统带来了巨大负担[4]。尽管脓毒症指南不断更新,其发病率仍有持续增长的趋势,住院病死率依然居高不下,成为ICU非心脏重症患者最常见的死亡原因之一。Vincent JL等[6]一项基于全球脓毒症流行病学数据的研究结果显示,ICU中脓毒症发病率高达29.5%,病死率达25.8%。严重脓毒症(含脓毒性休克)在国内ICU 的发病率为37.3%,ICU 病死率及住院病死率分别为28.7%和33.5%[7]。
脓毒症发病机制复杂,涉及机体多个器官功能改变,对其发病本质的探索一直是全球的研究热点。近年来,国内外学者分别从炎症反应失衡[8]、免疫功能障碍[9]、凝血异常[10]、免疫网络[11]、线粒体损伤[10]、内质网应激[11]、细胞自噬[12]、基因多态性[3]等方面对脓毒症的发病机制进行了研究,并取得一定的进展。基因多态性是指基因序列上的变异,其决定了人体对应激刺激的易感性和耐受性、临床表现的多样性、对药物治疗效果的差异性。目前,关于脓毒症基因多态性的研究涉及了肿瘤坏死因子家族、白细胞介素家族、热休克蛋白70、CD14、纤溶酶原激活因子抑制剂等[13]。探索脓毒症与基因多态性的关系将有助于从基因水平揭示脓毒症的发病机制。
核受体(NR)是一类依赖配体激活的转录因子超家族,由于配体激活机制,核受体成为了机体环境与基因组之间的连接。核受体4亚家族A群(Nuclear Receptor Subfamily 4 Group A,NR4A)是一类早期即刻反应基因,细胞在感受到环境的生理与物理刺激时,该家族的基因就会迅速反应,产生相应的蛋白,作为转录因子来调节相应基因表达从而来应对这些刺激[14]。Nur77是NR4A家族中的一员,定位于12q13染色体上[15],Nur77由数个功能结构域组成,包括N末端激活功能1结构域,具有两个允许DNA相互作用的锌指的中心DNA结合结构域和C末端配体结合域,其中C末端配体结合域对于募集小亲脂性分子(配体)在这些受体的转录活性中起分子开关的作用。近期研究发现NR4A家族在多种细胞的增殖、分化、凋亡及物质代谢中起重要的调节作用。Nur77可通过调节细胞因子和生长因子的活性在多种疾病炎症反应过程中发挥重要作用[16]。临床研究发现,Nur77在炎性疾病,如:滑膜炎、结肠癌、银屑病、动脉粥样硬化斑块、多发性硬化都有异常表达。体外实验显示Nur77可被多种促炎性因子诱导快速表达,如脂多糖、肿瘤坏死因子-α、干扰素-γ、白细胞介素、氧化型低密度脂蛋白等[17]。巨噬细胞中,Nur77可在多种刺激作用下快速表达,其诱导机制主要是NF-κB依赖性的转录激活作用。NF-κB可以与Nur77启动子中的反应元件结合从而激活Nur77,进而激活多种炎症基因的表达,参与多种炎症性疾病的发生发展[18]。以往的研究表明,Nur77在炎症反应中可能具有双向的调控作用,即,Nur77既可促进炎症反应,亦可抑制炎症发生。然而,由于这些研究仅仅局限于体外的细胞水平,仍无法证实Nur77在体内炎症调控中的作用及功能。最近的研究强调了Nur77作为炎症反应的重要调节剂的作用。Nur77被炎症刺激迅速诱导,以非配体依赖的转录因子形式调节巨噬细胞炎症免疫功能,表明它们是引发炎症所必需的。然而,Nur77抗炎特性表明这种急性调节可能是一种旨在解决后期炎症的保护性反应。Wu H等[2]发现Nur77通过与TRAF6相互作用充当Toll样受体/白介素-1受体信号传导的负调节物,这种相互作用阻止了TRAF6的自身泛素化和寡聚化,并随后抑制NF-κB活化和促炎细胞因子的产生。因此表明,Nur77参与负反馈机制以维持炎症平衡[19]。Li XM等[20]发现敲除Nur77的小鼠可发生较严重的全身性免疫及炎症性疾病,且炎症因子白介素6的表达显著提高,而Nur77在克罗恩病以及溃疡性结肠炎患者病变组织中的表达明显低于正常组织,说明孤儿核受体Nur77可作为负性调节因子参与调控炎症性肠病的发生发展[2]。
综上,Nur77基因的rs10876228、rs11169989、rs1283155、rs2242107四个位点都存在三种基因型,这三种基因型分布频率在脓毒症组与健康对照组间差异无统计学意义,提示这四个Nur77基因位点多态性与脓毒症易感性可能无相关性。
