植物次生物质对美国白蛾食物利用的影响1)
2020-05-29刘晓霞姜礅孟昭军严善春
刘晓霞 姜礅 孟昭军 严善春
(东北林业大学,哈尔滨,150040)
美国白蛾(Hyphantriacunea)别名秋幕毛虫,属鳞翅目(Lepidoptera)灯蛾科(Arctiidae),是世界性检疫害虫之一,于1979年在我国辽宁省丹东市首次发现[1],随后迅速扩散蔓延到山东、陕西、河北、天津和上海等省市。美国白蛾食性杂、繁殖量大、适应性强,对其所到地区的林木都造成了严重危害。尽管国家对美国白蛾的防控高度重视[2-3],采取了一系列防治措施,其危害范围仍然不断扩大,据国家林业局2018年公布结果显示,美国白蛾在我国已扩散到11个省份,县级疫区达572个,其中15个为2018年新增疫区[4-5]。
植物自身拥有对害虫的防御机制,次生物质是此类防御机制运行的重要“执行者”,在昆虫危害植物时,影响昆虫的取食行为[6]。Gershenzon et al.[7]的研究发现,植物中的倍半萜类次生物质阻碍了鳞翅目昆虫口器对葡萄糖、蔗糖和肌醇刺激的响应,达到抑制昆虫取食的目的。一些植物次生物质会引起昆虫中肠组织发生病变,破坏其消化系统,造成其消化功能异常,最终导致昆虫死亡[8]。Kostal et al.[9]研究发现植物中存在大量可以作为蛋白酶抑制剂的防御物质,当昆虫取食植物后,植物中的蛋白酶抑制剂会降低昆虫之后对其的取食。严俊鑫等[10]认为,单宁是植物中一类具有多抗性的酚类物质,会影响昆虫的取食。董钧峰等[11]研究结果表明,棉酚可显著降低烟青虫(Helicoverpaassulta)的食物消耗率,番茄苷不仅抑制烟青虫的取食,对其食物消耗率和利用率也有显著抑制作用;辣椒素会刺激烟青虫的取食;而棉酚对棉铃虫(Helicoverpaarmigera)却有辅食作用,番茄苷对棉铃虫各营养指标均无影响。单宁可引起棉铃虫幼虫对食物的消耗率减小、转化率增大,而食物利用率无显著变化[12]。用添加没食子酸的人工饲料饲喂美国白蛾4龄幼虫后,其食物消耗率下降,食物利用率却上升[13]。人参茎叶总皂苷明显抑制小菜蛾(Plutellaxylostella)的取食[14]。
美国白蛾可危害300多种林木、果树、农作物和其他野生植物,它为何能逾越如此众多植物的化学防御?本研究在人工饲料饲中添加不同质量分数单宁和水杨苷,饲喂美国白蛾幼虫,分析其4~6龄幼虫的取食量、排粪量、食物消耗率、转化率以及食物利用率,以期探明不同次生物质对美国白蛾食物利用的影响,分析其对寄主植物广谱适应机理,为探索防控美国白蛾新途径提供理论依据。
1 材料与方法
1.1 供试虫源及饲养方法
美国白蛾卵块及人工饲料由中国林业科学院森林生态环境与保护研究所提供。水杨苷又称水杨甙,化学式为C13H18O7,购自上海源叶生物科技有限公司。单宁又称丹宁,化学式为C76H52O46,购自上海麦克林生化科技有限公司。试验前,先用体积分数为10%的甲醛溶液将卵块浸泡30 min,之后用蒸馏水冲洗3次,晾干后将卵块置于养虫盒中,放入光照培养箱内孵化饲养。饲养条件为,温度(25±1)℃,湿度70%~80%,光周期16L∶8D。待幼虫孵化后,用人工饲料饲喂至3龄,之后将刚蜕皮的3龄幼虫分为9组,每组3个重复,每个重复30多头幼虫。在人工饲料中分别添加质量分数为0.50%、0.75%、1.00%、1.50%的水杨苷,或质量分数为0.20%、0.40%、0.60%、0.80%的单宁,对照组不添加次生物质,饲养幼虫。每天更换1次饲料,直至所有幼虫化蛹或死亡。
1.2 营养指标测定
每天称量各处理组取食前、取食后饲料的质量、空白组饲料初始质量、空白组饲料最终质量,以及幼虫粪便的质量,同时测定各龄幼虫的龄初体质量和龄末体质量。用未接虫的饲料测算校正失水率,用于计算校正失水率、体质量增长量、取食量、食物消耗率、食物转化率和食物利用率[15-16]。各指标的计算方法如下:
校正失水率=(空白组饲料初始质量-空白组饲料最终质量)/空白组饲料初始质量;
体质量增长量=龄末体质量-龄初体质量;
取食量=(取食前饲料质量-取食后饲料质量)/(1-校正失水率);
食物消耗率=((取食量-排粪量)/取食量)×100%;
食物转化率=(昆虫体质量增长量/(取食量-排粪量))×100%;
食物利用率=(昆虫体质量增长量/取食量)×100%。
1.3 数据处理与分析
用SPSS 21.0软件进行单因素方差分析,比较美国白蛾幼虫在不同次生物质处理组的取食量、排粪量、食物消耗率、食物转化率及食物利用率的差异显著性。
2 结果与分析
2.1 单宁和水杨苷对美国白蛾取食量和排粪量的影响
单宁和水杨苷各处理组美国白蛾4~6龄幼虫取食量和排粪量的情况见表1、表2。由表1可知,单宁各处理组的取食量,4~5龄均不同程度地高于对照组,在4龄时,单宁质量分数为0.20%和0.60%的处理组显著高于对照组(P<0.05);5龄时,单宁各处理组均显著高于对照组(P<0.05);6龄时,单宁各处理组的取食量均显著低于对照组(P<0.05)。不同龄期相同质量分数处理组的取食量,从4龄到5、6龄均先增加后减少,且6龄时的取食量均小于4龄时的。同一龄期不同质量分数处理组间的取食量,在4龄时,0.20%和0.60%处理组均显著高于0.40%和0.80%处理组;5龄时各处理组间均无显著差异;6龄时,0.80%处理组显著低于0.60%处理组(P<0.05)。单宁各处理组的排粪量,在4龄时均高于对照组,但差异不显著(P>0.05);5~6龄时,除了5龄的0.80%处理组与对照组无显著差异(P>0.05)外,其他处理组均显著低于对照组(P<0.05)。不同龄期相同质量分数各处理组间的排粪量,0.20%和0.40%处理组从4~6龄逐渐减少;0.60%和0.80%处理组先增加后减少,且6龄低于4龄;同一龄期不同质量分数处理组间的排粪量多无显著差异。
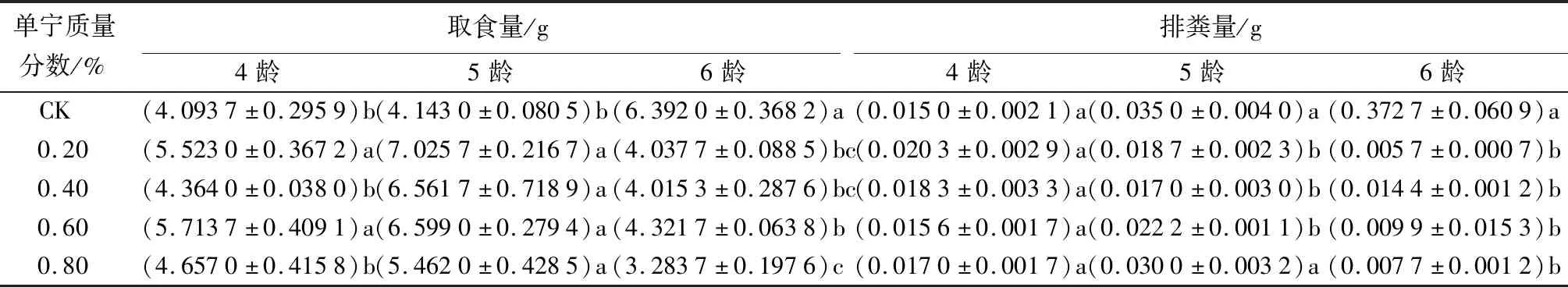
表1 单宁对美国白蛾取食量和排粪量的影响
注:表中数据为平均值±标准误,n=9。同列不同小写字母表示同龄期不同处理组之间差异显著(P<0.05)。
由表2可知,水杨苷各处理组幼虫的取食量,在4龄时,水杨苷质量分数为0.75%处理组低于对照组(P>0.05),其余处理组均高于对照组,但其中只有0.50%处理组与对照组差异显著(P<0.05);5龄时,1.00%处理组的取食量低于对照组,其余处理组均显著高于对照组(P<0.05);6龄时,各处理组与对照组均无显著差异(P>0.05)。不同龄期相同质量分数处理组的取食量,从4龄到5、6龄,0.50%质量分数处理组先增后减,并在6龄时低于4龄;1.00%质量分数处理组的取食量先减少后增加,至6龄时高于4龄;0.75%和1.50%处理组的取食量逐渐增加。同一龄期不同质量分数处理组间的取食量,在4~5龄时,0.50%处理组的取食量均显著高于其他处理组;6龄时,0.50%处理组的取食量显著低于1.50%处理组的(P<0.05)。水杨苷各处理组的排粪量,在4~5龄时均高于对照组,其中,4龄时,质量分数为1.00%的处理组与对照组差异显著(P<0.05);5龄时,各处理组均显著高于对照组(P<0.05)。6龄时均低于对照组,其中质量分数为0.50%和0.75%处理组与对照组差异显著(P<0.05)。不同龄期相同质量分数处理组间的排粪量,0.50%处理组从4~6龄先增加后减少,1.00%处理组从4~6龄先减少后增加,其余处理组均随着龄期的增长而增加;同一龄期不同质量分数组间的排粪量均无显著差异。

表2 水杨苷对美国白蛾取食量和排粪量的影响
注:表中数据为平均值±标准误,n=9。同列不同小写字母表示同龄期不同处理组之间差异显著(P<0.05)。
2.2 单宁和水杨苷对美国白蛾食物利用的影响
各处理组美国白蛾4~6龄幼虫对食物利用情况见表3、表4。由表3可知,单宁处理组的食物消耗率随龄期的增长而增大,4龄时,质量分数为0.20%和0.60%的处理组高于对照组,质量分数为0.40%和1.00%的处理组低于对照组,但差异均不显著(P>0.05);5~6龄时,单宁各处理组的食物消耗率均显著高于对照组(P<0.05);同一龄期不同质量分数组间多无显著差异。单宁各不同处理组的食物转化率与利用率多显著低于对照组,在同一质量分数处理组,随龄期的增长与对照的差异越来越大,如单宁质量分数0.20%的处理组,食物转化率,4龄是对照的0.754倍,5龄是对照的0.097倍,6龄是对照的0.044倍;食物利用率,4龄是对照的0.755倍,5龄是对照的0.098倍,6龄是对照的0.046倍。单宁各不同质量分数处理组的食物转化率和利用率从4~6龄先减小后增大,但最终都小于4龄;同一龄期的单宁各不同质量分数组间无显著差异。
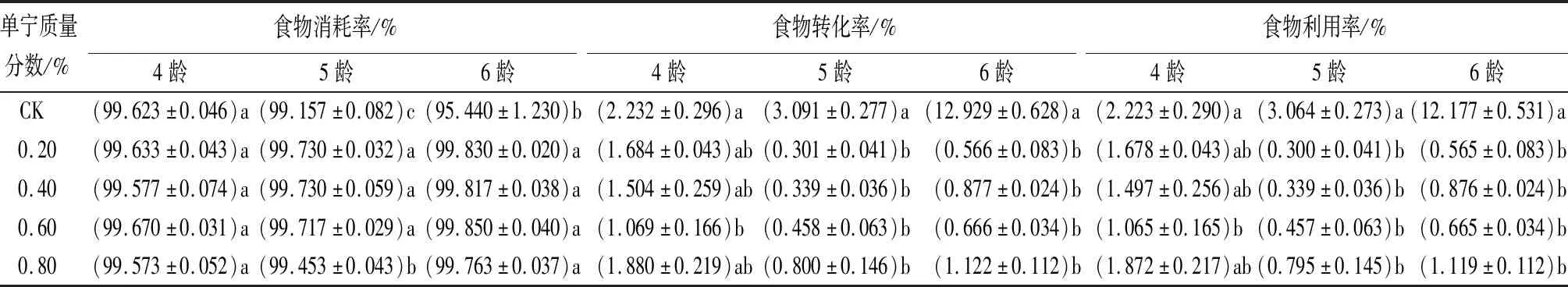
表3 单宁对美国白蛾食物利用的影响
注:表中数据为平均值±标准误,n=9。同列不同小写字母表示同龄期不同处理组之间差异显著(P<0.05)。
由表4可知,水杨苷处理组的食物消耗率,随着龄期的增长而减小,与对照相比,在4~5龄时,1.00%质量分数处理组显著低于对照(P<0.05),其余处理组均与对照组差异不显著(P>0.05);在6龄时,1.00%质量分数处理组与对照无显著差异,其余处理组均显著高于对照组(P<0.05)。同一龄期不同质量分数处理组间的食物消耗率,在4~5龄,0.50%处理组均显著高于1.00%处理组(P<0.05),其余各不同质量分数组间均无显著差异;6龄时,各不同质量分数组间均无显著差异。水杨苷各不同处理组的食物转化率和利用率均随着龄期的增长而增大,但不同质量分数处理组与对照的差异显著不同,如0.50%处理组,在4~5龄均低于对照,但差异不显著(P>0.05),6龄时显著低于对照(P<0.05);0.75%处理组,在各龄期与对照之间均无显著差异(P>0.05);1.00%处理组,在4龄时高于对照但差异不显著(P>0.05),5龄时显著高于对照(P<0.05),6龄时低于对照但差异不显著(P>0.05);1.50%的处理组,4龄时低于对照但差异不显著(P>0.05),5龄时显著高于对照(P<0.05),6龄时显著低于对照(P<0.05)。同一龄期不同质量分数处理组间的食物转化率和利用率,4龄时,0.75%、1.00%处理组均显著高于0.50%处理组(P<0.05);5龄时,1.00%处理组均显著高于0.50%、0.75%和1.50%处理组(P<0.05),1.50%处理组显著高于0.50%处理组(P<0.05);6龄时,0.75%、1.00%处理组均显著高于0.50%和1.50%处理组(P<0.05)。

表4 水杨苷对美国白蛾食物利用的影响
注:表中数据为平均值±标准误,n=9。同列不同小写字母表示同龄期不同处理组之间差异显著(P<0.05)。
3 结论与讨论
在昆虫与植物的相互作用中,昆虫对食物的消耗率、转化率和食物利用率作为探讨植食性昆虫与寄主植物相互关系的重要参数[16-17],可反映昆虫对食物的利用情况[18]。植物化学成分尤其是次生物质在植食性昆虫寄主植物演变过程中起重要作用[19]。昆虫可通过自身的主动调节,最大程度地消化利用所取食的食物,这可能是其在长期进化过程中对不利环境条件的一种主动适应方式[18]。
食物利用率是食物消耗率和转化率的产物,能表征昆虫将取食的食物转换为其体质量的效率[20-21]。昆虫在不同的环境条件下,可通过调整食物消耗率和转化率之间的比例,以维持其正常生长发育所需的食物利用率[22-23]。本研究结果显示,与对照相比,随着龄期的增长,单宁处理组,美国白蛾幼虫的取食量和排粪量整体变化趋势一致,均表现为先促后抑。说明美国白蛾可通过调整取食量和排粪量之间的平衡来响应食物质量的变化。其食物消耗率、转化率和食物利用率,在4龄时多与对照差异不显著(P>0.05);在5~6龄时,食物消耗率均显著高于对照组,食物转化率和利用率多显著低于对照组(P<0.05),且与对照的差异越来越大。说明美国白蛾5~6龄幼虫的食物消耗率虽然在提高,但不足以弥补其转化率的降低,进而造成食物利用率显著低于对照组(P<0.05)。Feeny[24]研究表明,单宁能降低植食性昆虫对食物中营养物的消化利用,抑制其对食物的利用效率。有研究者认为,单宁的营养拮抗作用可能归结于它的毒性和抑食作用[25-26]。本研究中,美国白蛾幼虫的食物利用对单宁不同质量分数的响应未表现出明显的规律性,其原因可能是设定的质量分数梯度差异太小的缘故。
水杨苷多数处理组美国白蛾4~6龄幼虫的取食量高于对照组,而排粪量随龄期的增长逐渐低于对照,食物消耗率由4~5龄与对照差异不显著,到6龄显著高于对照,可能是由于水杨苷刺激了美国白蛾的取食。Yazdani et al.[22,27]发现,薰衣草(Lavandulaangustifolia)和百里香(Thymusvulgaris)精油均显著提高桑螟(Glyphodespyloalis)的食物消耗率。董钧锋等[11]研究发现,棉酚会使棉铃虫幼虫的取食量显著增加,辣椒素会提高棉铃虫的食物消耗率。植物次生物质对昆虫食物利用的影响与其浓度密切相关[28]。李路莎等[29]研究结果显示,类黄酮高质量分数抑制美国白蛾的食物利用,低质量分数则促进其食物利用。在本研究中,水杨苷处理下,美国白蛾的食物转化率和利用率,0.75%处理组在各龄期与对照均无显著差异(P>0.05);1.00%处理组在5龄时与对照差异显著且高于对照(P<0.05);低质量分数0.50%和高质量分数1.50%处理组则随着龄期的增长最终显著低于对照。说明水杨苷对美国白蛾幼虫食物利用的影响具有剂量效应,表现为0.75%质量分数无明显影响,1.00%质量分数有促进作用,质量分数过低(0.50%)或过高(1.50%)则为抑制作用。
综上所述,单宁对美国白蛾幼虫的食物利用具有明显的抑制作用,而水杨苷对其食物利用的影响与剂量相关,在一定的剂量阈限内表现为促进作用,超出剂量阈限则表现为抑制作用。
