平潭光幼屿典型植物群落优势种生态位特征
2020-05-25郑建忠何雅琴史晓洁邓传远
魏 凯,郑建忠,杨 铭,何雅琴,史晓洁,邓传远
(1.福建农林大学园林学院,福建 福州 350002; 2.福建林业职业技术学院园林系,福建 南平 353000)
由于我国海洋经济的繁荣兴盛,推动海岛旅游与海洋渔业规模发展壮大,海岛植被破坏问题却越来越显著[1]。海岛生境特殊,岛上一般多裸露的岩石,淡水资源匮乏,常年受海风侵蚀,植被覆盖率低,海岛生态系统属于典型的脆弱型生态系统,海岛环境一旦破坏将很难恢复[2]。海岛由于四周海水环绕不利于物种交流,小面积内也可能分布大量特有种,造成小型海岛被破坏后更难进行植被恢复[3]。福建目前拥有大小海岛共1 433个,其中有居民海岛101个,无居民海岛1 332个,广布在我国东南沿海[4]。本次研究对象光幼屿为无居民海岛,位于福建省平潭县平潭岛东部,与流水镇北海岸相距约0.3 km。平潭岛由于人为开发,在1990—2013这二十多年间植被覆盖状况逐渐变差,平均覆盖度逐年降低,且最近十年降低速率更快[5]。与平潭岛隔水相望的无居民海岛光幼屿植被如何,是否受到人为干扰都有待调查。目前,我国海岛植被研究主要集中于物种多样性、植物区系、植被与环境间的关系,海岛植物功能性状以及海岛植物保护也有涉及[6-7]。生态位理论历来是生态学领域的热点之一,其广泛用于濒危植物保护、种间关系、群落演替、生态系统退化等方面的研究[8-11]。生态位是指种群利用各种资源的能力以及与相关种群间的功能关系,可以定量研究种内、种间以及生境之间的相互关系,测度不同种群在群落中的地位与作用[12-13]。生态位在生物多样性的保护以及濒危保护植物的评价方面具有重要作用[14]。有关海岛植被生态位的研究还较少,现有研究也多局限于乔木层或灌草层,缺乏对群落乔灌草3层次系统的生态位研究[15-17]。本研究选取小型海岛光幼屿为研究对象,对岛上典型植物群落展开样方调查,通过对样方中乔灌草优势种进行生态位研究,拟解决以下两个问题:(1)探究岛上不同物种在群落中的功能地位以及物种间的相互关系; (2)为海岛植物保护以及海岛植被恢复的树种选择提供理论基础。
1 研究地区与方法
1.1 研究区概况
光幼屿地处福建省平潭县,地理坐为北纬25°34′54″,东经119°50′04″,岸线长度1 075 m,面积0.062 km2,最高点高程24.2 m。岛体呈南北走向,基岩海岸,表层土主要为红壤土。光幼屿为南亚热带海洋性季风气候,年平均温20~25 ℃,岛上7级以上的大风每年可达125 d,常集中在7—9月,年均降雨量1 196.2 mm,蒸发量1 917.4 mm[2]。经实地踏勘,岛屿南部主要分布木麻黄林、木麻黄与台湾相思(Acaciaconfusa)混交林,北部常年受海风影响,分布稀疏的黑松林以及黑松与台湾相思混交林,岛上植被整体覆盖良好。
1.2 样地设置与调查
根据种-面积曲线的研究方法,该地区常绿阔叶林取样面积达1 200 m2即具有代表性[18],于2018年9月进行植被调查,选取岛上典型植物群落为样地,根据岛上植被情况共设置6个20 m×20m的样地,每个样地划分为4个10 m×10 m的乔木样方,在每个样地的四角取4个5 m×5 m 的灌木样方,另外取5个1 m×1 m的草本样方,分别位于样地的四角以及中部位置,各样地信息见表1。分别记录每个样方的物种与环境信息,物种信息包括灌木与草本的种名、高度、冠幅、数量、盖度,乔木(胸径≥3 cm或树高≥3 m)[19]在灌草记录的指标上补充记录其胸径、枝下高,环境信息包括经纬度、坡度、坡向以及坡位。对在野外调查中不能鉴定的植物进行特征部位拍照并采集标本,带回实验室通过查阅《福建植物志》[20]、《中国植物志》[21]或请教专家进行鉴定。岛上植被根据实地调查具体可分为木麻黄群系、木麻黄+台湾相思群系、黑松群系、黑松+台湾相思群系。本文以这4个群系作为“资源状态”,依据每个种群在不同群系中的重要值计算各个物种的生态位宽度以及不同物种间的生态位重叠值,分析各物种在群落中的功能地位。调查结果中,外来物种以及入侵物种参照《福建植物志》与《中国入侵植物名录》[22]来确定。
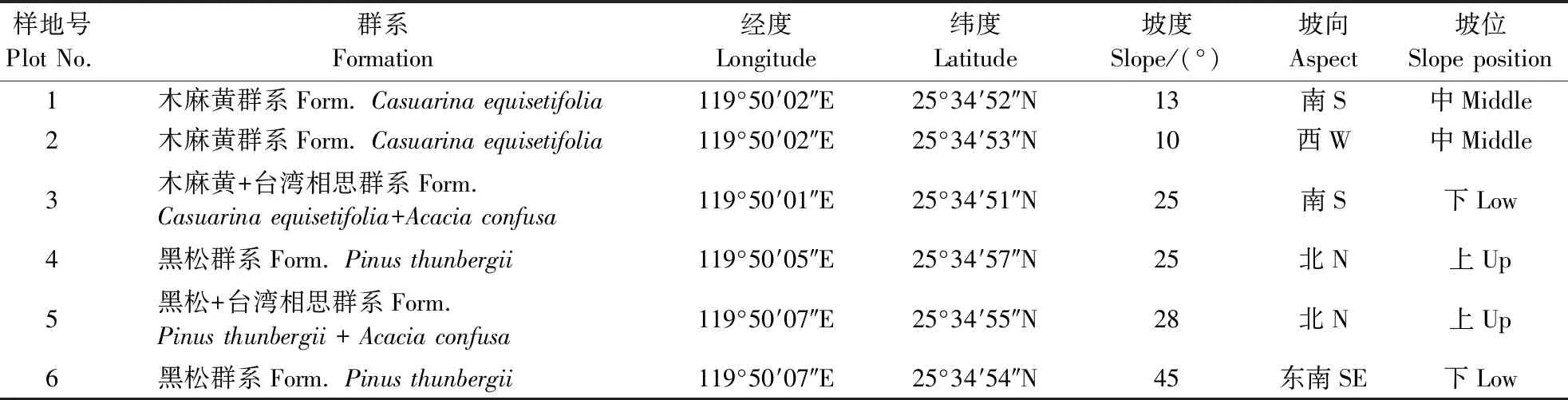
表 1 光幼屿样地基础信息Table 1 Basic information of the sample plots of Guangyou Island
1.3 数据计算
(1)重要值计算
各层物种的重要值计算方法如下[23]:
乔木层: 重要值=(相对多度+相对显著度+相对频度)/3
(1)
灌木层、 草本层: 重要值=(相对频度+相对盖度)/2
(2)
式中:相对多度表示某一物种在全部样方中的个体数与所有物种个体数总和的比值;相对显著度表示某一物种在全部样方中的胸高断面积与所有物种胸高断面积总和的比值;相对频度表示某一物种在全部样方中出现的频度与所有物种频度总和的比值;相对盖度表示某一物种在所有样方中的盖度与所有物种盖度和的比值。
(2)生态位宽度计算
(3)
式中:Bi为种i的生态位宽度,Pij=nij/Ni,nij为物种i在资源j上的重要值,本研究中用不同群系表示各资源位,Ni表示物种在所有资源位上的重要值之和,r表示资源位的数量,即群系数量[24]。
(3)生态位重叠值计算
(4)
式中:Oik为物种i和k的生态位重叠值,Pkj为种k在第j个资源位上的重要值[24]。
1.4 数据处理
为确保研究结果的准确性,去除调查中的偶见种以及重要值较小的物种,最终乔灌草三层分别选择4种、9种、10种共20(部分种重叠)种优势种进行生态位计算。数据计算和统计分析采用Excel 2016软件和SPSS 25.0软件。
2 结果与分析
2.1 植物群落物种组成
调查样方中共有种子植物38种(表2),隶属于22科38属,全部为单种属,其中菊科物种数最多,包括9属9种,占全部物种数的23.7%。含两种物种的科分别为禾本科(Poaceae)、百合科(Liliaceae)、豆科(Fabaceae)、大戟科(Euphorbiaceae)、鼠李科(Rhamnaceae)、马鞭草科(Verbenaceae),占总科数的27.3%。其余科为单种科,占总科数的68.2%,主要为胡颓子科(Elaeagnaceae)、苋科(Amaranthaceae)、酢浆草科(Oxalidaceae)。乔木层植物生长良好,高度、胸径、枝下高平均值分别为4.32 m、 7.1 cm、 1.04 m,平均密度为0.2 株·m-2,但是物种数偏少,只有4种,分别是木麻黄、黑松、台湾相思、桑(Morusalba)。灌木层共有植物15种,平均高度为0.78 m,平均密度为0.9 株·m-2,主要有木麻黄、黑松、雀梅藤(Sageretiathea)、铁包金(Berchemialineata)。草本层有植物22种,主要有山菅(Dianellaensifolia)、狗牙根(Cynodondactylon)、假还阳参(Crepidiastrumlanceolatum)、积雪草(Centellaasiatica)。海岛特有植物11种,占全部物种数的29.0%,主要有黑松、木麻黄、台湾相思、滨海白绒草(Leucaschinensis)、华南狗娃花(Heteropappusciliosus)、烟豆(Glycinetabacina)。外来物种8种,占全部物种数的21.1%,分别是黑松、木麻黄、桑、烟豆、马缨丹(Lantanacamara)、滨海白绒草、假还阳参、假臭草(Praxelisclematidea),其中马缨丹和假臭草为恶性入侵植物。
2.2 重要值与生态位宽度
从乔灌草三层次物种重要值与生态位宽度可知,乔木层中黑松的重要值最大(1.619),其次是木麻黄(1.575),两者相差仅0.044,属于共优势种,台湾相思与桑的重要值偏小,分别为0.622、 0.184,属于伴生种(表3)。台湾相思的生态位宽度最大(1.028),黑松的生态位宽度最小(0.665)。灌木层中,木麻黄的重要值最大(1.025),其次是铁包金、雀梅藤和黑松,紫珠(Callicarpabodinieri)的生态位宽度最大(1.089),其次是小叶黑面神(Breyniavitis-idaea)、铁包金、木麻黄。乔灌层均呈现重要值大的物种生态位宽度不一定大,重要值小的物种生态位宽度不一定小。草本层中山菅、狗牙根、假还阳参的重要值最大,分别为1.052、 0.878和0.314,山菅、酢浆草(Oxalisgriffithii)、狗牙根的生态位宽度最大,分别为1.230、 0.968和0.690,山菅的重要值与生态位宽度都最大,利用资源的能力最强,在草本层中占据绝对优势。假还阳参和积雪草利用资源的能力弱,生态位宽度最小,分别为0.596和0.458。

表 2 光幼屿样方调查植物名录Table 2 List of plants surveyed in Guangyou Island
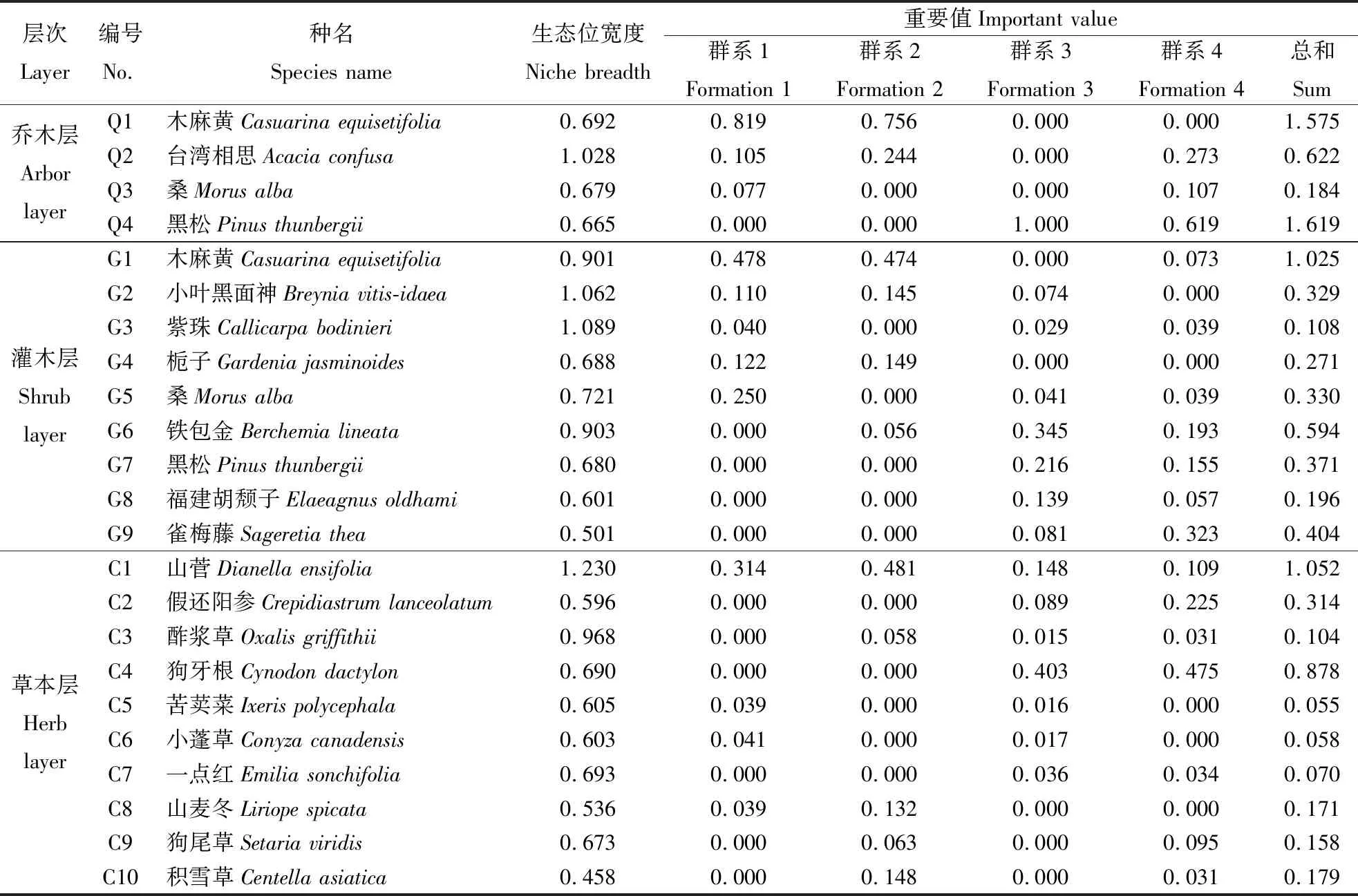
表 3 优势种生态位宽度与重要值Table 3 Niche breadth and important value of dominant species
2.3 生态位重叠
乔木层中各物种间的生态位重叠值分布在0~0.743,台湾相思的生态位宽度最大,与其他物种生态位重叠程度也最高,同木麻黄和桑的生态位重叠值分别为0.636和0.743(表4)。黑松和木麻黄分布在不同群系,生态位重叠值为0。灌木中各优势种按照生态位重叠大小可分为3组:生态位重叠值为0.515~0.989的19对,占总对数的52.8%;生态位重叠值为0.041~0.427的14对,占总对数的38.9%;生态位重叠值为0的3对,占总对数的8.3%(表5)。灌木层中紫珠的生态位宽度最大,整体上同其他物种的重叠程度也最高,与栀子(Gardeniajasminoides)的生态位重叠程度最小其重叠值达到0.402,紫珠所利用的资源与其他物种相近,生存竞争激烈。雀梅藤的生态位宽度最小,与小叶黑面神的生态位重叠值为0.091,与栀子的生态位重叠值则为0。草本中生态位重叠值为0.533~0.996的19对,占总对数的42.2%;生态位重叠值为0.087~0.495的18对,占总对数的40.0%,生态位重叠值为0的7对,占总对数的15.6%;生态位重叠值为1.000的1对,占总对数的2.2%(表6)。与灌木相比,草本植物中生态位重叠在0.515以上的比重少,生态位重叠值为0的占比更大,草本层各物种间生态位重叠较灌木层弱。生态位宽度较宽的物种间生态位重叠一般较大,生态位宽度大与生态位宽度小的物种间也可能产生较大的生态位重叠,如生态位宽度最大的山菅与生态位宽度最小的积雪草生态位重叠值为0.818。生态位宽度小的物种间也能产生较大的生态位重叠,生态位宽度为0.458的积雪草与生态位宽度为0.536的山麦冬(Liriopespicata)生态位重叠位为0.940,生态位宽度为0.603的小蓬草(Conyzacanadensis)与生态位宽度为0.605的苦荬菜(Ixerispolycephala)生态位完全重叠。
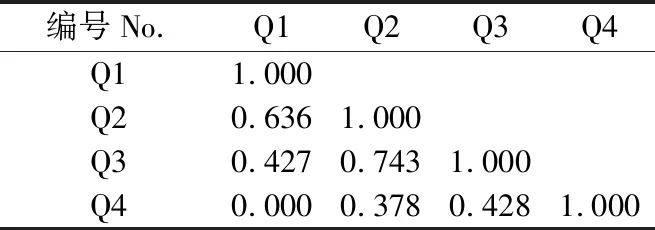
表 4 乔木层物种生态位重叠Table 4 Niche overlap of species in tree layer
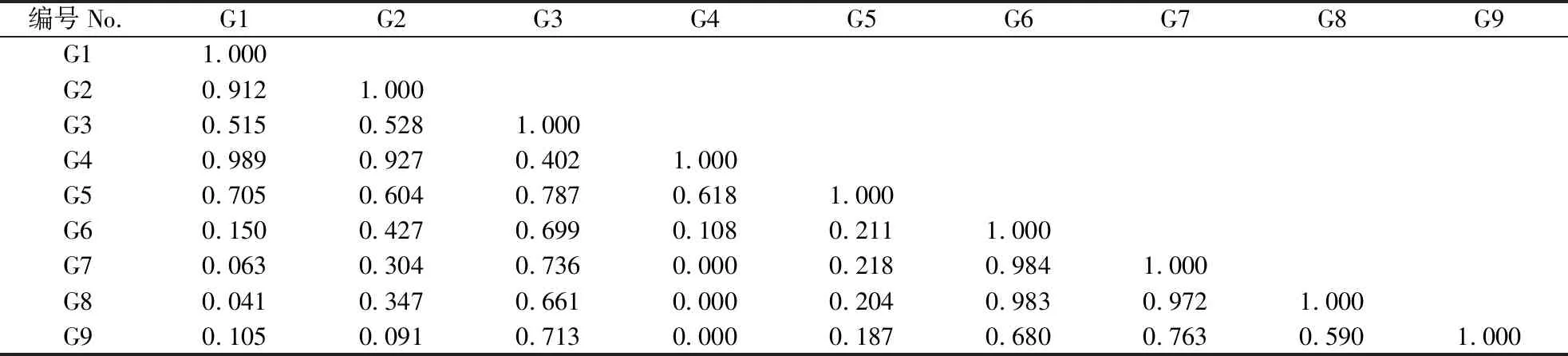
表 5 灌木层优势种生态位重叠值Table 5 Niche overlap value of dominant species in shrub layer
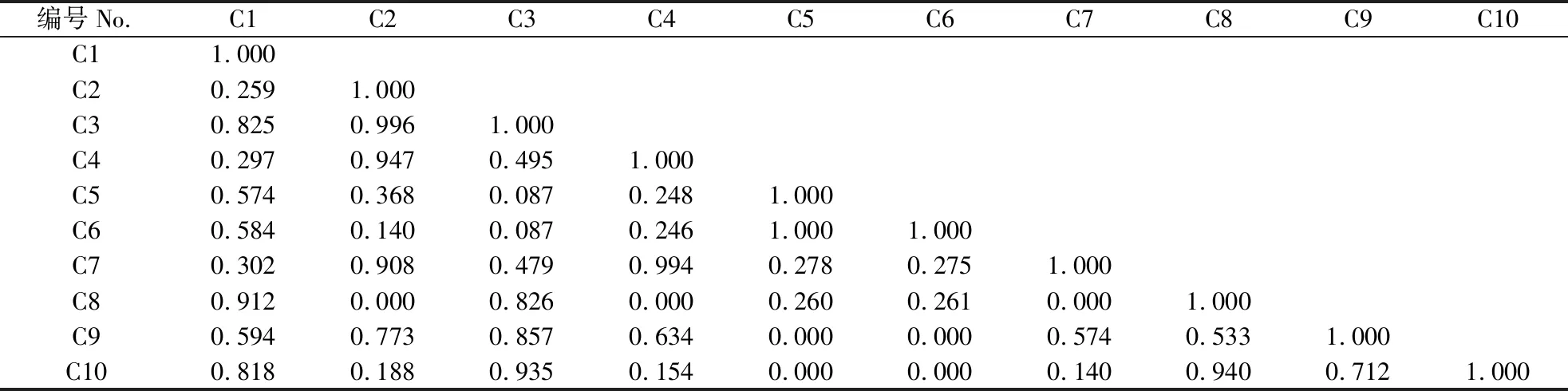
表 6 草本层优势种生态位重叠值Table 6 Niche overlap values of dominant herbaceous species
3 讨论与结论
重要值体现了物种在群落中的优势程度,而生态位宽度则反映了植物种群对环境资源的利用能力与生态适应性,两者都可作为度量物种在群落中地位与作用的指标[25]。光幼屿呈南北走向,西面背靠海坛岛,岛上南部受海风侵袭少,环境相对稳定,海岛北部多裸露的岩石,且直接受到海风的侵袭,海岛上复杂多样的生境为不同物种的生长提供生存空间,为优势物种占据更多的生境提供了可能,也成为测定物种潜在生态位的天然实验场所。将乔灌草共20种优势种重要值与生态位宽度排序进行比较,39.1%的物种生态位宽度较重要值排序降低,43.5%的物种上升,仅17.4%的物种保持不变,重要值与生态位宽度没有明显的相关性(P>0.05)。黑松在乔木层重要值最大,生态位宽度降为第4,木麻黄在灌木层重要值最大,生态位宽度降为第4,对比乔灌层中生态位宽度最大的台湾相思、紫珠可知,后两种植物调查中分布较广,在各群系分布较为均匀,导致生态位宽度最大。张贵平等[26]也研究表明生态位宽度在很大程度上取决于物种的分布频数。调查结果显示,同一群落中植物间存在相互影响,木麻黄林下共统计到12种植物,灌木与草本各6种,呈现出林下植物种类少,数量分布不均匀的特点,灌木层中木麻黄数量占灌木总数的66.1%,草本中出现次数少于5次的物种占总物种数的66.7%,相反,同等样方面积的黑松林下共统计到23种植物,灌木8种,草本15种,其中黑松数量占灌木总数最多(15.6%),草本中出现次数少于5的物种占总物种数的53.3%,造成这种情况的原因可能是木麻黄对其他物种存在化感作用,李慧敏等[27]研究表明木麻黄除自身合成化感物质外,土壤微生物的代谢产物也是化感物质的主要来源,这些化感物质的存在对其他物种的生存产生抑制作用。调查发现的木麻黄幼苗证明木麻黄林存在自然更新能力,杨彬等对海南岛海防林研究认为木麻黄自身无法实现天然更新[28]。已有研究表明水分是制约木麻黄种子萌发的主要限制因子,并且木麻黄天然更新能力弱的结论大都是在木麻黄纯林中得出,常年纯林经营导致土壤贫瘠,且林下木麻黄枯枝不易分解,凋落层厚,种子即使萌发后根系不能及时扎入土壤,造成种子发芽率低自然更新困难[29]。光幼屿上的木麻黄一般是20世纪六七十年代人为种植[30],经过多年结实繁殖,部分种子萌发长出幼苗,且幼苗分布在木麻黄与台湾相思混交林中,地面凋落层薄,且位于生境较好的南部,土壤含水量相对较高,这些有利条件利于木麻黄种子的萌发。草本层中,山菅的重要值与生态位宽度最大,可通过分株和种子繁殖,在半阴或光线充足的环境都可生长,对资源利用力强,在竞争中处于有利地位,是草本层的优势种。一点红(Emiliasonchifolia)、小蓬草、苦荬菜的重要值排最后三位,但是生态位宽度在10种植物中分别排第3位、第7位和第6位,原因可能是这3种都是菊科1年生植物,靠种子传播繁殖,种子小且轻,加上岛上海风频繁,种子传播距离远,分布范围广,但海岛恶劣的环境导致种子发芽率低,物种数量少,因此出现重要值小而生态位宽度大的情况。
在环境恶劣资源短缺的情况下,物种的生态位宽度呈现增大的趋势,生态位宽度越大植物才越有可能获取满足自身生存的资源,更能在恶劣的环境存活[31]。本次海岛植被调查中各物种生态位宽度值在0~1.230的范围内,生态位宽度大于0.600的物种数占总数的82.6%,高伟等[17]对东山岛海岸带潺槁树林37种植物用不同公式进行生态位宽度计算,分别有48.6%、 32.4%的物种生态位宽度大于0.600,光幼屿上各物种生态位宽度值普遍偏大,符合植物在海岛恶劣环境下的生存策略。生态位宽度越大,不同物种利用资源发生重合的概率越高,物种间发生生态位重叠的可能性越大,甚至与生态位宽度较小的物种也可能存在较大的生态位重叠[32]。如生态位宽度较大的小叶黑面神与木麻黄的生态位重叠值达0.912,山菅和酢浆草的生态位重叠值达0.825。生态位宽度最大的紫珠与生态位宽度最小的雀梅藤生态位重叠值可达0.713,两者性喜半阴半湿的环境,一般分布在林下,常生长于土壤含水量高的地方,相似的生态习性导致生态位宽度差异较大的两物种同样拥有较高的生态位重叠。整体来看,乔灌草三层次中生态位重叠值大于0.500的对数占总对数的47.1%,各物种生态位重叠普遍较高,各物种间资源竞争较为激烈,导致该结果的原因可能是海岛资源匮乏,各物种占据较宽的生态位引起较大的生态位重叠,也有可能是群落演替的原因,或者两者兼有。苏志尧等[33]和陈子林等[34]研究发现,处于演替阶段中的物种间生态位重叠值较高,当演替至顶级群落时各物种间达到一种相对平衡,种间竞争才有所降低。光幼屿上植被属于亚热带植物群落,按照演替规律其植被演替方向为灌草丛—针叶林—针阔混交林—常绿阔叶林[35]。目前,光幼屿正处于针阔混交林阶段,所以各物种间生态位重叠程度大,竞争能力弱的物种被竞争能力强的物种替代,不断有新的适生物种加入演替过程,推动群落演替的进行。海岛环境特殊,受台风、海浪等因素的影响,岛上不同部位生境差异大,导致植物拥有不同的生活习性。乔灌草三层共有11对物种间生态位重叠值为0,如灌木层的福建胡颓子、雀梅藤同栀子的生态位重叠值为0,从实地调查来看,福建胡颓子与雀梅藤主要分布在黑松林、黑松与台湾相思混交林中,这两个群落中乔木少,冠幅小,林下阳光充足,栀子则主要分布在荫蔽条件好的木麻黄林下,生态习性的差异导致他们没有产生生态位重叠,正是这些物种生态位的分化对提高群落的生物多样性具有重要作用。
光幼屿植物种类和数量少,林下较为空旷,群落结构简单,生态系统脆弱,容易遭到破坏,应加强对植物的保护工作。本次调查中,硕苞蔷薇(Rosabracteata)、茅莓(Rubusparvifolius)、九里香(Murrayaexotica)、算盘子(Glochidionpuberum)、龙葵(Solanumnigrum)、烟豆、华南狗娃花这些物种在调查中零星分布,重要值小,未纳入生态位计算,但对于提高岛上物种多样性具有重要作用,并且烟豆属于国家二级保护植物,在海岛生境中应加强对这些植物的保护力度。生态位计算结果显示,桑、栀子、福建胡颓子、小蓬草、苦荬菜等植物,重要值小且生态位宽度小,与其它物种间生态位宽度重叠普遍较小,这些植物大多生长林下、石头缝等适宜的小生境中,生长的环境一旦被破坏,这些植物可能在岛上消失。木麻黄、台湾相思、铁包金、紫珠、小叶黑面神、山菅、狗牙根生态位宽度大,在海岛上分布广,可考虑作为海岛植被修复时的先锋物种。除物种选择外还要注意种间配置,比如木麻黄纯林林分单一,更新困难,可与台湾相思混植,紫珠喜阴凉环境,最宜种植在林下。种植时应考虑立地条件,土壤瘠薄的地方可先行种植灌木或草本,待养分积累到一定程度时再种植乔木。此外,引入外来物种时应对所选物种进行风险评估,虽然外来物种能解决当地物种单一的困境,如该岛上的外来物种木麻黄和黑松对提高海岛物种多样性,维持群落稳定具有重要作用,但是外来物种可能存在入侵风险,本次调查中发现恶性入侵植物假臭草与马缨丹。目前这两种入侵植物在调查中零星分布,种群数量少,重要值分别为0.118、 0.075,在中竞争处于弱势地位,对海岛植物群落产生的危害程度较低,但是这两种植物对周围植物存在化感作用[36-37],抑制其他物种生长,并且二者自身繁殖能力强,扩散速度快,对维持海岛植物群落的稳定具有潜在隐患。
