贵州3个斑鳜野生种群形态学差异
2020-05-25潘秋芝陈敦学马秀慧
黄 胜,潘秋芝,安 苗,余 科,李 珊,陈敦学,马秀慧
( 1.贵州大学 动物科学学院,贵州 贵阳 550025; 2.贵州农业职业学院,贵州 贵阳 551400 )
形态度量分析是鱼类种群生态学研究方法之一,传统形态性状测量法主要集中在鱼体的头、尾部和轴向,未能全面地利用鱼体躯干部提供的有用信息[1-2],对种间及种以上水平的鉴定有效但对种群间的鉴定往往效果较差[3]。使用多变量形态测量a指标,利用框架结构将鱼体划分为功能单元区来测量鱼的外部形态,这样可以更准确地反映鱼的形态差异[4]。基于传统形态学和框架结构数据的多元分析法已在卡拉白鱼(Chalcalburnuschalcoides)[5]和日本鲭(Scomberjaponicus)[6]等不同鱼类形态变异识别中被广泛运用。

1 材料与方法
1.1 试验材料
斑鳜样本于2016—2018年采自沅江、柳江和乌江3水系在贵州省境内的10个站点(图1),共计收集372尾样本。沅江水系7个站点共253尾,体长6.51~ 29.51 cm,体质量9.43~484.63 g;柳江水系2个站点共86尾,体长7.80~26.10 cm,体质量9.43~370.35 g;乌江水系思南点33 尾,体长7.90~28.50 cm,体质量10.98~472.63 g(表1)。

图1 斑鳜种群的采样点

表1 3个斑鳜种群基本信息
1.2 数据测量
共测量了372尾鱼的17 110个数据。数据分为两类。一类是传统形态学数据包括可数性状和可量性状两类[2,4,18]。可数性状包括:背鳍棘、背鳍条、胸鳍、腹鳍棘、腹鳍条、臀鳍棘、臀鳍条、尾鳍、脊椎骨、幽门垂、侧线鳞、第一鳃弓外鳃耙12项;传统可量性状测量了全长、体长、体高、体宽、头长、吻长、眼径、眼间距、尾柄长和尾柄高10项(图2);可数性状和可量性状共22项。另一类为框架数据,选取11个同源坐标点建立框架图[4,17,19](图3)测量24项框架参数。46项原始形态测量值精确至0.01 cm。
1.3 数据处理和分析
1.3.1 可量数据标准化和参数选择
为消除鱼体规格大小对试验结果的影响,34个传统可量性状和框架数据除以体长并采用公式标准化:
Madj=log10[(M/L)+1][20]
式中,M为原始测量数据,Madj为标准化后的数据,L为体长(cm)。
基于标准化的数据,先进行方差齐性检验,再采用单因素方差分析方法和Pearson相关性分析方法[20],筛选种群间具有显著差异的参数用于下一步分析。数据处理和分析采用Excel 2019和SPSS 15.0软件完成。

图2 斑鳜形态测量指标

图3 斑鳜框架结构示意
1.3.2 差异性检验
参照文献[21]对亚种进行识别和划分规则,对3个种群12项比例性状的差异系数进行差异性分析,计算公式如下:
CD=|M1-M2|(SD1+SD2)
式中,CD表示差异性系数,M1、M2分别代表2个种群某种比例性状的平均值,SD1和SD2为对应的标准差。
1.3.3 可数性状处理
将12项可数性状经单因素方差分析[22]和差异性检验,数据处理和分析采用Excel 2019和SPSS 20.0软件完成。
1.3.4 主成分分析
使用KMO和Bartlett检验性状参数是否适合因子分析[20],通过建立相关系数矩阵,以主成分法提取公因子,选择特征值大于1的因子为主要因子,并采用方差最大正交旋转法对因子负荷矩阵进行旋转,从而获得各主成分的特征值和贡献率。基于各因子得分,绘制主成分散点图[17-18,20]。
1.3.5 逐步判别函数分析
采用逐步判别法建立分类判别函数,据此对各种群进行预测分类,建立典型判别函数得分图,选出对判别贡献较大的参数建立各种群的判别公式,计算判别准确率和综合判别率。
1.3.6 聚类分析
分别求出34项形态比例参数的平均校正值,然后用平均校正值进行聚类分析,所采用聚类方法为欧氏距离的最短系统聚类法,绘出聚类关系图。
2 结 果
2.1 参数选择
33项标准化性状逐步进行齐性检验(P>0.05)、单因素方差分析(P<0.05)和Pearson相关性分析,最终筛选出12项(体宽/体长、眼间距/体长、尾柄长/体长、2-3/体长、4-3/体长、6-8/体长、8-7/体长、8-10/体长、10-5/体长、11-9/体长、7-5/体长和5-3/体长)用于差异性分析、主成分分析、逐步判别函数。
2.2 性状差异性分析
对3个斑鳜种群的12项显著性状参数进行差异性分析,其结果见表2。乌江种群与沅江种群和柳江种群在尾柄长性状上的差异系数分别为1.579、1.405,达到了亚种分类的差异;而沅江种群与柳江种群之间达不到亚种差异分类这个阀值(1.28)[21]。
2.3 可数性状
经单因素方差分析,3个斑鳜种群在可数性状上,除了腹鳍棘、臀鳍棘和脊椎骨数3项性状上无显差异之外,其余9项可数性状种群间存在显著差异。在差异性系数上均小于1.28,达不到亚种差异阀值(表3)。
2.4 主成分分析
经KMO统计量(KMO=0.846)和Baetlett球型检验结果(P<0.01),如果相关系数矩阵不是单位阵,适宜进行因子分析降维。

表2 3个斑鳜种群间12项显著性状的差异系数
注:*表示差异系数大于亚种分类阈值1.28.
Note:* denotes that the difference coefficient is greater than the subspecies classification threshold of 1.28.

表3 3个斑鳜种群可数性状
注:同行中上标不同字母的平均值表示差异显著(P<0.05).
Note: the means with different letters in the same line are significant difference (P<0.05).
主成分前6项累积贡献率达72.668%,其中基础特征值>1的前4项累积贡献率为58.894%,即前4项包含了12个相互独立变量58.894%的变异信息(表4)。3个斑鳜种群在主成分1中起关键作用的性状是尾柄长/体长、11-9/体长、4-3/体长和10-5/体长,主要体现其尾柄长、尾鳍背部起点至尾鳍腹部起点、头部末端至胸鳍和背鳍末端至臀鳍起点的差异(表5);主成分2中起关键作用的性状是眼间距/体长,反映了眼间距之间的大小;主成分3中起关键作用的性状是体宽/体长,反映了鱼体宽度间的差异。
由主成分散点图(图4)可见,沅江与柳江斑鳜种群在主成分1轴-1的右侧,而乌江斑鳜种群主要分布在主成分1轴-1的左侧;沅江斑鳜种群与柳江斑鳜种群部分交叉重叠,沅江和柳江种群与乌江种群基本上不重叠,可以明显的从中分离出来。

表4 3个种群主成分分析的特征值和积累贡献率(主成分1~6)
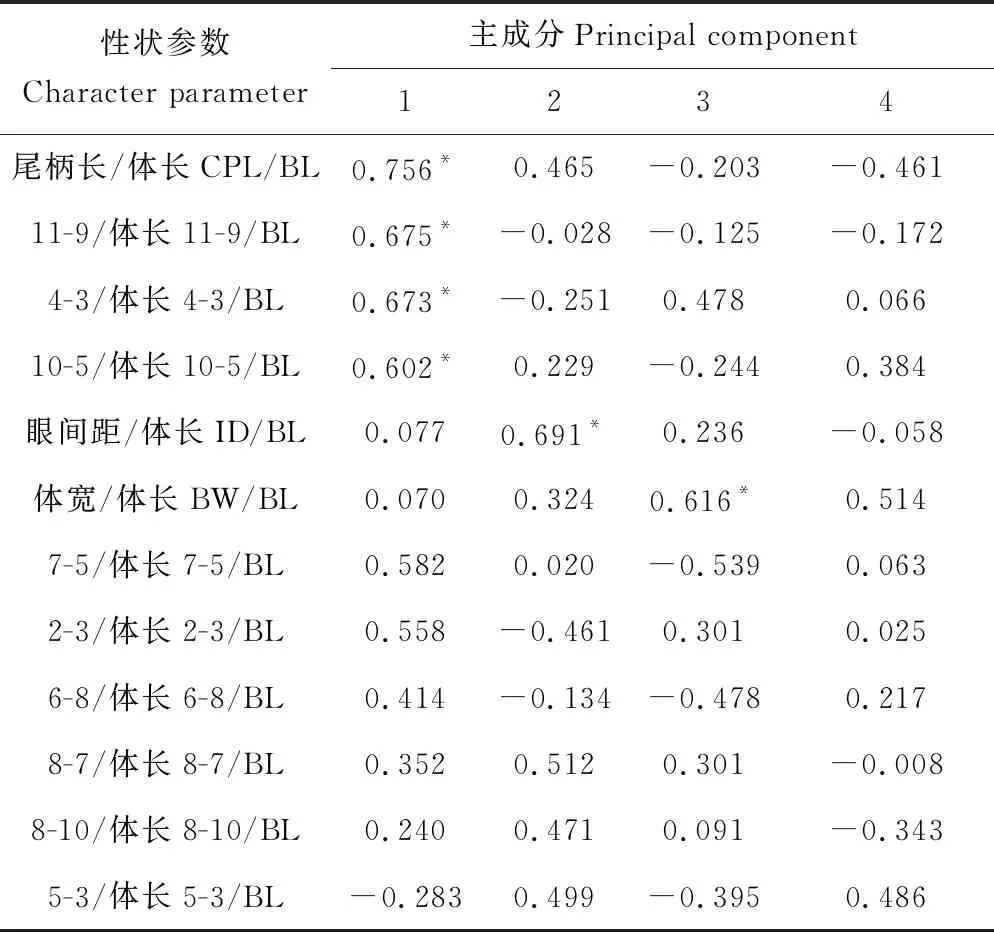
表5 12个性状在主成分分析中的因子负荷值
注:*表示负荷值>0.600. Note:*indicates load value>0.600.
2.5 判别分析
对12项参数性状,进行逐步判别分析后,发现判别分析处理中综合判别率较低。为提高综合判别率,除了筛选出的12项参数性状之外,增加了头长/体长、体高/体长和尾柄高/体长。这3项性状也刚好处于鱼体的头部、躯干部和尾部。在传统的测量方法[1-2]和结合框架测量中[17]都具有一定的代表性。筛选出10项判别贡献大的性状(体宽体长、眼间距/体长、尾柄长/体长、尾柄高/体长、体高/体长、4-3/体长、6-8/体长、7-5/体长、8-7/体长、11-9/体长)(表6)。
采用这10项性状参数的判别函数系数(表6)建立了3个地理种群的判别函数方程。
沅江种群:
y=742.615x1-290.747x2+94.787x3+521.545x4-249.267x5+119.505x6+346.960x7+572.493x8-517.198x9-129.144x10-539.736
柳江种群:
y=713.422x1-292.650x2+109.060x3+497.023x4-264.802x5+95.767x6+372.274x7+561.096x8-490.175x9-126.732x10-512.370
乌江种群:
y=819.789x1-255.325x2-30.216x3+510.9405x4-231.842x5+133.048x6+328.255x7+601.286x8-541.747x9-107.034x10-561.040

图4 3个斑鳜种群前两个主成分散点图
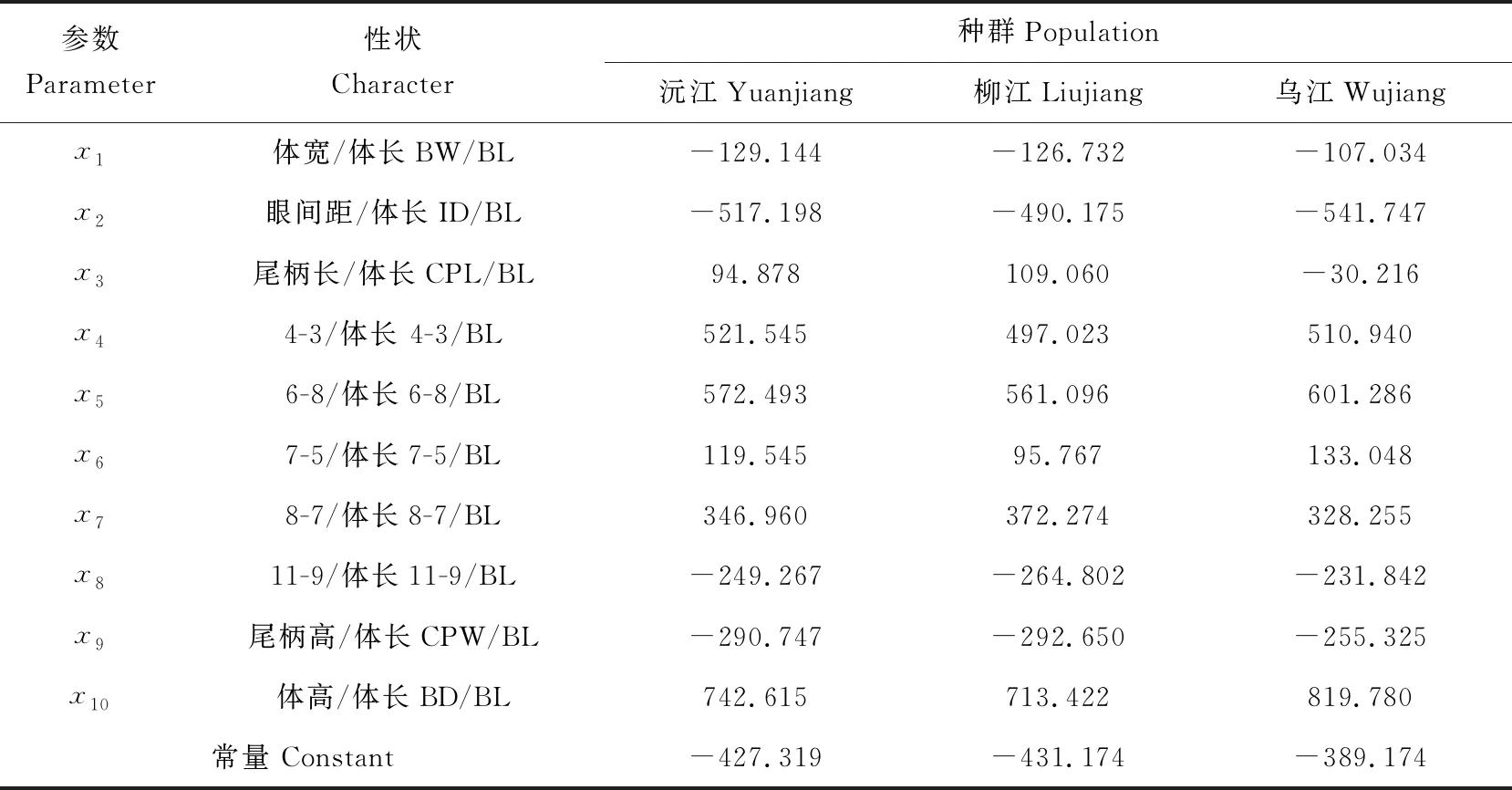
表6 3个斑鳜种群的Fisher线性判别函数系数
注: Fisher 的线性判别式函数. Note: Fisher′s linear discriminant function.
3个斑鳜种群判别函数系数见表6,根据建立的判别公式,可以更简便地判断野生斑鳜的种群归属。判别归类结果见表7。由表7可知,非交互验证时,沅江、柳江、乌江种群分别有19尾、40尾、4尾判别错误,判别准确率分别为92.5%、53.5%、81.8%,综合判别准确率为82.5%。交互验证时,沅江、柳江、乌江交互验证准确率分别为91.3%、52.3%、81.8%,交互判别准确率为81.5%。交互验证结果与直接使用判别公式进行判别的结果的判别准确率接近,说明判别公式是可信的。乌江种群相对独立,且沅江斑鳜种群与柳江斑鳜种群间形态较相似(图5)。

表7 3个斑鳜种群的判别分析结果
2.6 聚类分析
采用欧式距离的最短系统聚类法。对34项形态比例参数的平均校正值作聚类分析,结果见图6,3个野生斑鳜种群主要分为两支,沅江与柳江先聚为一个支,然后再与乌江种群聚在一起。
3 讨 论
3.1 多元统计分析在鱼类研究中的应用
具有相同形态特征的类群称之为一个种群,鱼类种群间的变异可为其种群结构的划分和种群鉴定提供一定基础数据和理论基础[19]。形状特征和性状差异是种群识别、分类和系统发育最有用的基础,并在形态学研究中发挥重要作用[23]。传统形态性状测量法主要集中在鱼体的头、尾部和轴向[1-2],未能全面地利用鱼体躯干部提供的有用信息,容易忽略一些重要的指标,难以全面反映鱼体形态差异的特征。随着对鱼类外形差异研究深入,形态特征研究也不再局限于视觉观察和宏观描述,演变为多变量统计分析,可以更准确地确定鱼群的形态特征[24]。李思发等[4]对尼罗罗非鱼(Oreochromisniloticus)5个品系间形态差异进行分析表明,框架分析法和传统分析法相结合能显著提高种群的判别分析能力。主成分分析、聚类分析、判别分析、差异性系数分析相结合的多元统计分析对鱼类差异性研究分析得到了广泛的应用,Mohaddasi等[18]对4个种群的卡拉白鱼(Alburnuschalcoides)形态差异分析时,先对参数进行KMO统计量和Baetlett球型检验,再做主成分分析提高了分析问题的深度和精度[17-19,23]。本研究结果表明,传统可数性状,可量化性状和现代框架性状的结合,利用多元统计分析方法获得3个斑鳜种群的形态差异,可以获得准确的结果,故多元统计分析在鱼类外形差异和变异等评价中具有很高的价值[23]。经方差分析,沅江、柳江和乌江3个斑鳜种群之间,背鳍棘、背鳍条、胸鳍条、腹鳍条、臀鳍条、尾鳍条、幽门垂、侧线鳞和第一鳃弓外鳃耙9项可数性状(P<0.01)存在显著差异性。统计学中,单因素方差分析具有简单易懂,易于理解的优点,缺点是在划分和识别鱼类种群时,无法给出精确的判别[25]。为弥补方差分析中的缺陷,有学者在分析中引入主成分分析、聚类分析和逐步判别分析[4]。主成分分析可以消除无关性状的相关影响,减少性状选择的工作量。
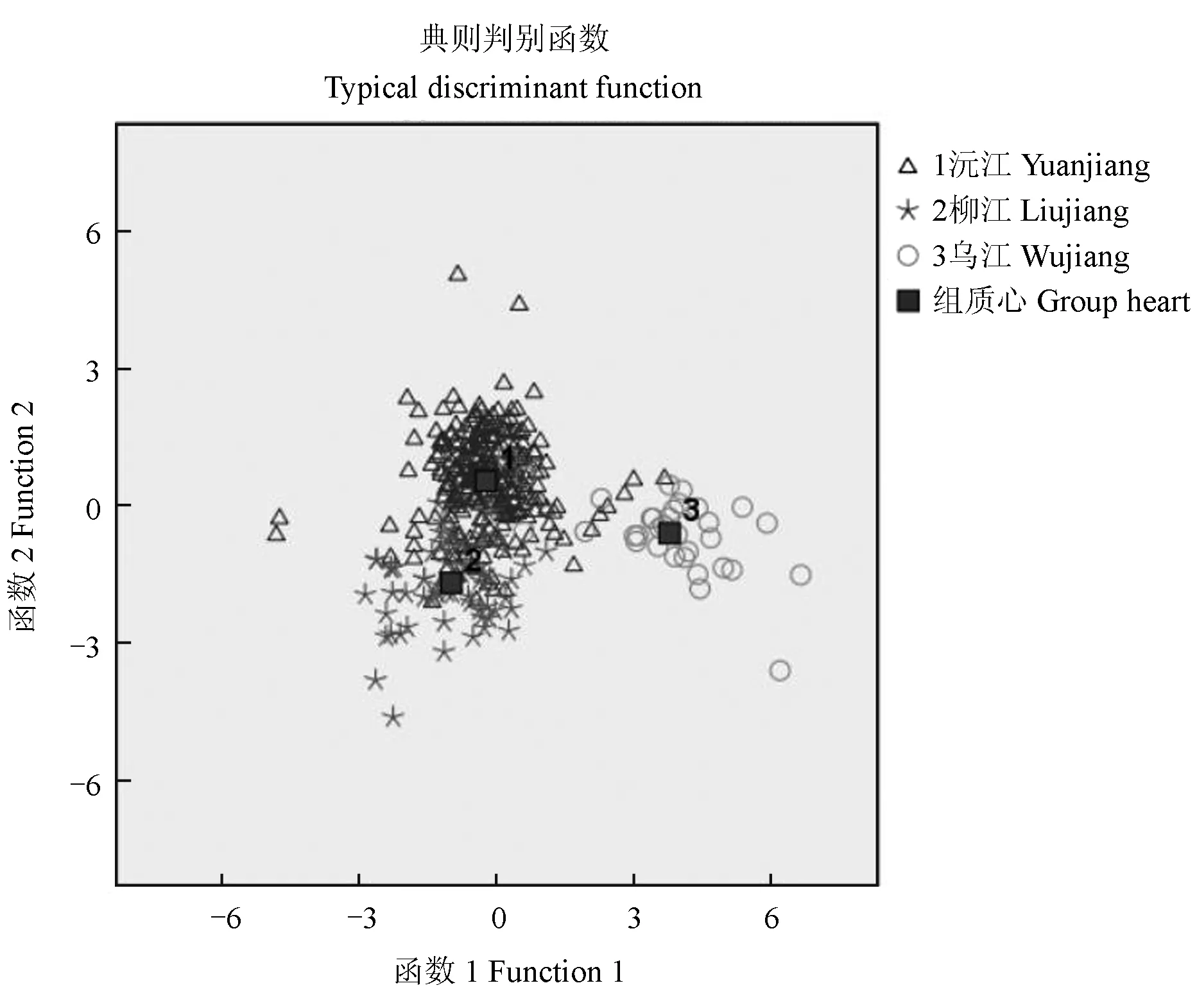
图5 判别函数散点图
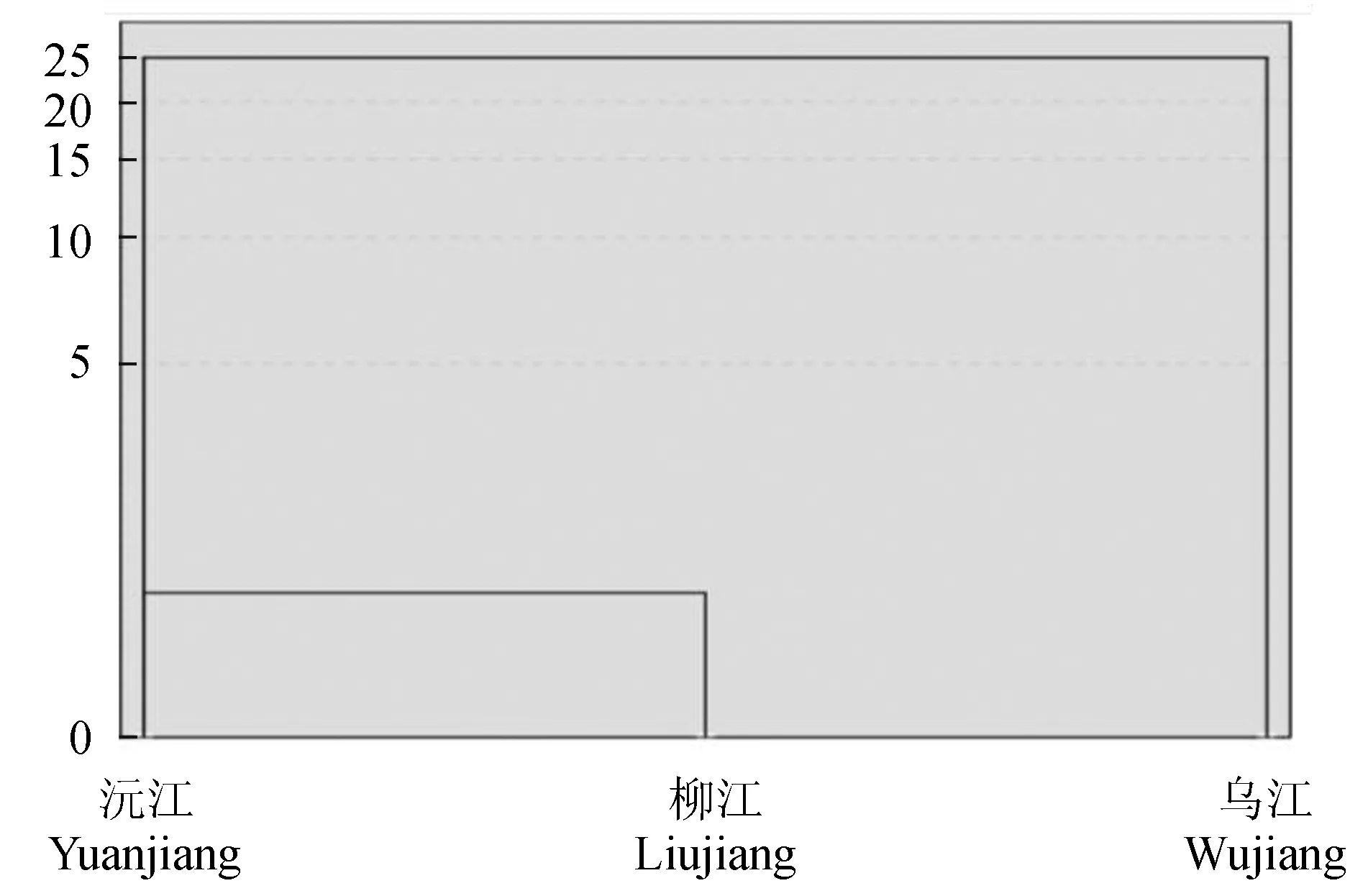
图6 3个种群聚类分析
3.2 3个斑鳜野生种群的形态鉴别
主成分分析结果显示,沅江斑鳜种群与柳江斑鳜种群的形态较为接近,而与乌江斑鳜种群的形态差异较为明显(图4)。主成分1中主要反映鱼体尾柄部、尾鳍背部起点至尾鳍腹部起点、头部末端至胸鳍、背鳍末端至臀鳍起点差异;主成分2中反映了鱼体眼间距的大小;主成分3中反映了鱼体之间体宽的差异。就这3个主成分而言,头部和尾柄以及体宽的部分性状特征的差异,是导致这3个斑鳜种群差异的关键性因素,这与对大眼鳜(Sinipercakneri)形态差异的研究结果[17,19]一致。判别分析结果可知,3个斑鳜种群的综合判别准确率为82.5%能够精确地判别斑鳜的类群归属;判别公式中贡献较大的变量多分布于鱼的尾柄部,3个种群的主要形态差异体现在尾柄部。聚类分析是通过将物理或抽象对象的集合分组,量化差异,直接反映不同种群间的亲缘关系[26],由形态学聚类分析结果可见,沅江斑鳜种群和柳江斑鳜种群先聚为一支,再与乌江斑鳜种群聚为一支;3种多元统计分析结果类似,却从不同的角度反映种群之间形态差异,起到相互印证,相互补充的效果[4]。
鱼类形态变异是其对特定栖息环境条件最直观的适应性变化,鱼类形态特征除了受自身遗传因子影响外,还与所处环境有关[26-28]。研究表明,导致鱼类形态学差异的因素是多方面的。除遗传因子外,鱼类所处的外部环境所带来的影响也不容忽视[17,19]。不同地理种群之间的形态变异通常是由于当地生境差异所引起的获得性结果。水温、水深、水流速度、透明度、食物组成均会是导致鱼类形态学差异的关键因子。早在20世纪90年代,有学者研究发现,地理位置较近的种群之间,遗传相似度较高,地理隔离对物种遗传分化和形态差异有很高的影响[19,29]。本研究中,沅江种群和柳江种群之间差异性系数值较小,差异系数值均小于1.28,可能与这2个种群所在水域地理位置较近,生境类似有关。乌江种群与沅江种群和柳江种群在尾柄长/体长上差异性系数值均大于1.28,达到亚种分化的水平。乌江种群采集于思南河段,此河段水流湍急,河谷纵深,与其他2个斑鳜种群水域地理位置相差较远,种群差异较大。且受禁渔期影响,3个种群在采样时间不同(表1),乌江种群采集于冬季,而沅江和柳江种群采集于夏季,不能完全覆盖鱼类形态的完整性,会存在一定的局限性[6]。在体质量、体长上也有不同(表1),乌江种群在体质量和体长的均值上,均大于沅江种群和柳江种群,虽经过标准化处理,消除了部分因素影响,但随着鱼类年龄的增长,鱼体上的指标不一定会按比例增长,所以体质量和体长均值大,可能也是造成乌江斑鳜种群与其他2个种群差异性的原因之一[4,6]。根据Mayr等[21]提出的75%亚种识别和鉴定的标准,一个物种在不同流域分布的种群之间存在形态特征差异,即Ⅰ种群75%的个体和Ⅱ种群97%的个体能够分开,那么这2个种群之间的形态特征差异系数值应为1.28,则2个种群应分属于不同的亚种,即1.28是亚种分化的阈值。本研究中,乌江斑鳜种群与沅江斑鳜种群和柳江种群虽在尾柄长/体长性状上差异系数值大于1.28,但在鱼类分类中,仅仅以形态特征的值1.28作为亚种划分依据,研究者须持谨慎态度[22]。造成种群之间形态差异的原因是多种多样的,具体的原因还有待今后进一步借助遗传学[26]和分子标记手段[9-11,30]来进行确认。
