螺旋藻多糖提高小白菜抗盐能力
2020-05-21程宇娇刘哲臧敦秀马浩天季春丽张春辉岳爱琴李润植崔红利
程宇娇 刘哲 臧敦秀 马浩天 季春丽 张春辉 岳爱琴 李润植 崔红利
摘 要: 在水培条件下探究不同浓度的PSP(螺旋藻多糖)能否提高‘奶油四季小白菜的抗盐性。生理生化检测显示,NaCl(5 g·L-1)胁迫显著抑制小白菜根长和鲜质量,且造成严重的生理伤害。水培液中添加PSP使得小白菜根长、鲜质量和叶绿素含量增加。PSP处理提高了小白菜体内超氧化物岐化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)的活性,增加了抗坏血酸、类胡萝卜素和脯氨酸含量。然而,PSP处理明显降低了小白菜体内MDA和H2O2含量。研究结果表明,PSP可通过提高盐胁迫下小白菜抗氧化酶活性、非酶类抗氧化物质含量以及积累渗透调节物质,有效清除过量的活性氧和缓解氧化损伤,进而增强小白菜抗盐胁迫的能力。作为一种绿色的植物生长促进剂,PSP可用于高值蔬菜的有机生产。
关键词: 小白菜;螺旋藻多糖;NaCl胁迫;生理特性;抗氧化性
中图分类号:S634.3 文献标志码:A 文章编号:1673-2871(2020)04-043-07
Abstract: In this study, polysaccharide from Spirulina platensis (PSP) was used as a biostimulant. The effect of PSP with different concentration on salt resistance of pakchoi (Brassica chinensis L.) under hydroponics was investigated. Physiological and biochemical analysis showed that NaCl stress (5 g·L-1) significantly inhibited the root length and fresh weight of pakchoi plants and caused severe physiological damage. Adding different dosages of PSP in hydroponic solution resulted in the enhancement in root length, fresh weight, and chlorophyll content of pakchoi. Moreover, PSP treatments increased the activities of superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT) in pakchoi, together with the elevated levels in the contents of ascorbic acid, carotenoid and proline, but reduction in MDA and H2O2. These results indicated that PSP could increase antioxidant enzyme activities, the content of non-enzyme antioxidants, and the accumulation of osmotic adjustment substances, and consequently result in the effective remove of excess reactive oxygen species and alleviation of oxidative damages caused by salt stress, leading to improvement in salt stress resistance of pakchoi plants. As a green growth-promoting regulator, PSP has potential application in the organic cultivation of high-value vegetables.
Key words: Pakchoi; Polysaccharide from Spirulina platensis; NaCl stress; Physiological characteristics; Antioxidant ability
隨着现代农业生产强度的增加,土壤盐渍化逐渐成为一个全球性的生态问题[1]。全球约20%的耕地和50%的灌溉土地受到了不同程度的威胁,且盐渍化土地面积仍在逐年增加[2],给农业生产带来了巨大的挑战。盐胁迫也成为抑制植物生长、降低农作物产量的主要环境因素之一。土壤中高浓度的盐会降低植物根系的吸水能力,严重影响植物新陈代谢[3]。盐胁迫会使植物体内产生大量活性氧(ROS),引起氧化胁迫,导致细胞损伤甚至死亡,最终造成植物产量或品质降低[4]。
因此,提高植物抗盐胁迫的能力是农业生产一个重要因素。为应对盐胁迫,植物形成了各种抗盐机制[5],包括增强抗氧化酶活性和非酶类抗氧化物质含量,维持自由基的代谢平衡[6],以及合成渗透调节物质,以调节渗透压,保护细胞免受伤害[7]。植物抗盐机制的表达亦可通过外施一些化合物调控。郭伟等[8]研究发现,盐胁迫下活性氧的积累和清除能力会受到多种内源和外源物质的影响。Bose等[9]也提出抗盐机制可以通过应用外源性生物刺激素得到进一步改善。
近年来,应用外源生物刺激素提高植物抗盐性备受关注。生物刺激素包含功能性物质、微生物及其次生代谢产物等,可增加作物对养分的吸收及利用效率,增强作物抗逆性、提高农作物质量[10]。黄腐酸在干旱胁迫条件下可促进作物生长, 提高作物产量[11]。平菇多糖能不同程度上提高盐环境下小白菜种子的发芽率, 促进根和芽的生长, 对NaCl胁迫具有缓解作用[12]。海藻糖处理能有效地通过刺激抗氧化防御和乙二醛酶系统而减轻盐胁迫对水稻的损伤[13]。海藻提取物也越来越多地用于农作物及瓜果蔬菜栽培管理。螺旋藻(Spinulina sp.)属于蓝藻门 (Cyanoptha)、颤藻科(Oscilatoriaece),是一种光自养的原核生物。生长周期短、可食用且营养丰富[14],是多种食品及保健品的主要原材料。螺旋藻多糖(polysaccharide from Spirulina platensis,PSP)是从螺旋藻中提取出来的水溶性多糖,含量高达螺旋藻干质量的14%~16%,且成分安全、易获取。具有抗氧化、降血糖和抗炎等生理活性[15],但其在农业上的应用以及诱导植物抗逆性等方面鲜有报道。
目前,土壤盐渍化问题严重影响了品质优良但抗盐性较差的小白菜品种的种植。因此,本文以‘奶油四季小白菜为试材,在水培条件下,从抗氧化以及渗透调节和植物生长等方面解析PSP能否促进其生长和提高抗盐胁迫水平,以期为绿色、安全的高附加值蔬菜瓜果生产提供新的生物刺激素和科学参考。
1 材料与方法
1.1 材料
PSP由山西农业大学分子农业与生物能源研究所提取纯化并储存,其平均分子量为29 600 Da,单糖组成主要为鼠李糖(24.7%),葡萄糖(16.15%)和半乳糖(13.32%),蒽酮—硫酸比色法测定其纯度为89.72%。
供试小白菜为‘奶油四季小白菜(pakchoi,Brassica chinensis L.)(河北沧州青县良达盛农技术推广中心),以下简称为小白菜。
1.2 方法
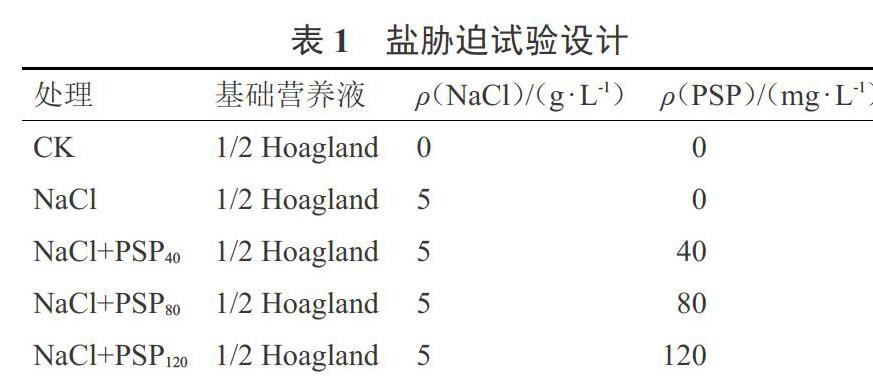
1.2.1 盐胁迫处理 试验于2019年8—10月在山西农业大学分子农业与生物能源研究所水培实验室进行。选取籽粒饱满,大小均匀一致的小白菜种子在育苗棉上发芽,2叶1心期移栽到水培营养盆中(25 cm×35 cm),每盆6株,漂浮板上间距7 cm,行距8 cm,室内温度25 ℃,每天光照时间14 h。待小白菜苗适应水培环境后,设计5组处理,见表1。每隔2 d换1次营养液,30 d后采收,进行各项生长及生理指标的检测。
1.2.2 生理指标的测定 测定小白菜根长及鲜质量时,用滤纸将其根部营养液吸干,测量其根长,然后用剪刀去掉地下部分,用天平测小白菜地上部分鲜质量,并记录。抗坏血酸(Ascorbic Acid,ASA)含量采用2,6-二氯靛酚滴定法测定[16]。叶绿素及类胡萝卜素(Carotenoids,CAR)的测定:釆用丙酮-乙醇提取,在665 nm、649 nm和470 nm 波长下比色测定(FW)。参考王学奎[16]的方法测定超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)和过氧化氢酶(catalase,CAT)的活性。参考张志良等[17]方法,在450 nm和532 nm处测定其吸光值,计算丙二醛(MDA)含量。采用Solarbio过氧化氢(H2O2)含量检测试剂盒测定H2O2含量。参考张志良等[17]方法,配置不同浓度的脯氨酸标准品,根据520 nm波长下吸光值绘制标准曲线,计算样品中萃取的脯氨酸含量。
1.3 试验数据統计学分析
各分组所得数据用DPS 7.05软件进行统计分析,采用单因素和LSD法进行方差分析和多重比较,数据利用GraphPad Prism8软件作图。
2 结果与分析
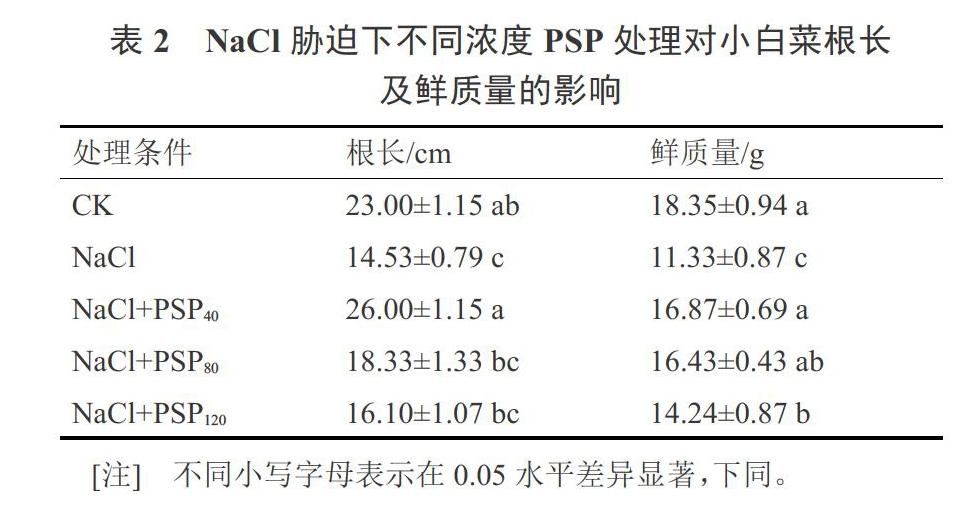
2.1 PSP对NaCl胁迫下小白菜根长及鲜质量的影响
小白菜在NaCl胁迫以及不同浓度PSP处理下生长30 d后收获,测得其根长及地上部分鲜质量如表2所示。NaCl胁迫下,小白菜单株根长和鲜质量都显著低于对照,当营养液中加入PSP后,对小白菜的根长及鲜质量的增长均有促进作用。PSP施用量为40 mg·L-1和80 mg·L-1时小白菜单株根长和鲜质量达到与对照相当的水平,差异不显著。
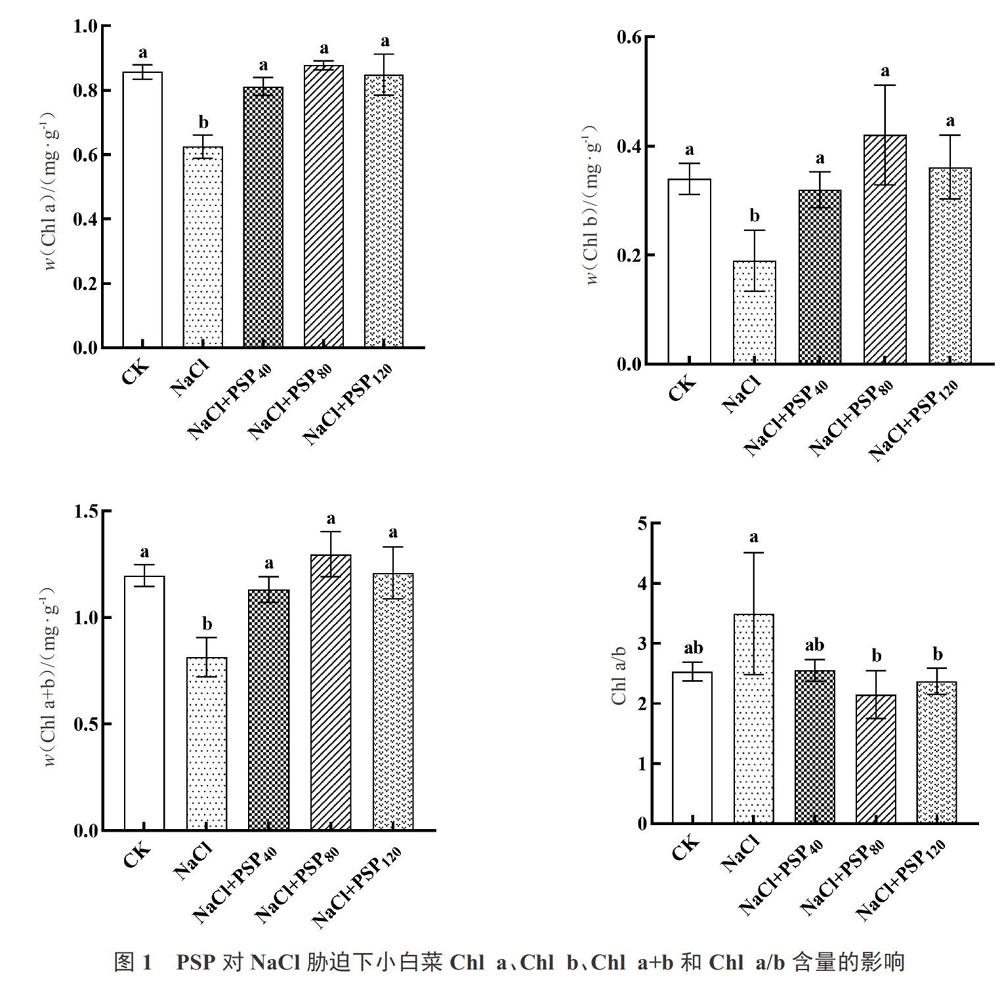
2.2 PSP对NaCl胁迫下小白菜叶绿素含量的影响
如图1所示,与CK组相比,盐胁迫显著降低了小白菜Chl a和Chl b的含量。添加PSP能显著增加NaCl胁迫下植株Chl a和Chl b的含量,且各浓度处理后其体内Chl a和Chl b含量均达到与对照相当的水平。与盐胁迫相比,PSP各浓度处理均能增加盐胁迫下小白菜体内Chl a+b的含量,增幅分别为38.99%、59.38%和48.56%。此外,40 mg·L-1和80 mg·L-1的PSP处理显著降低盐胁迫条件下小白菜体内Chl a/b。
2.3 PSP对NaCl胁迫下小白菜抗氧化酶活力的影响
SOD、POD和CAT是植物体内清除过剩ROS的主要酶系。NaCl胁迫下添加不同浓度PSP对小白菜抗氧化酶活力有明显影响(图2)。SOD、POD和CAT具有相似的变化趋势。与CK相比,NaCl胁迫下植株内SOD和CAT活力显著提高,而POD活力差异不显著。当盐胁迫下的小白菜添加不同浓度PSP后,与NaCl处理相比,3种抗氧化酶活力均明显增强。添加不同浓度PSP显著增强SOD活力。其中,添加80 mg·L-1 PSP浓度水平时,小白菜体内POD和CAT活力最强,分别增加了0.71倍和1.15倍。这表明,一定浓度的PSP可以提高植物体内抗氧化酶活力。
2.4 PSP对NaCl胁迫下小白菜非酶类抗氧化物质含量的影响
对不同剂量PSP处理的小白菜测试结果(图3)显示,与CK相比,NaCl胁迫显著降低了小白菜体内ASA和CAR含量,分别减少了21.38%和44.37%。添加PSP后,ASA和CAR含量明显增加。与NaCl处理相比,PSP各浓度水平均显著增加了盐胁迫下小白菜体内ASA和CAR的含量,且增幅最大的为80 mg·L-1 PSP浓度。当NaCl胁迫后添加时,相比于CK组,80 mg·L-1 PSP 处理使小白菜ASA和CAR含量增加了8.28%和25.87%,但差异不显著。
2.5 PSP对NaCl胁迫下小白菜丙二醛(MDA)含量的影响
MDA浓度可以直接反映ROS对植物细胞的伤害程度。如图4所示,与CK相比,NaCl胁迫条件下小白菜体内MDA含量显著升高了34.77%。添加PSP对降低NaCl胁迫条件下小白菜体内MDA的含量效果明显,且各浓度PSP处理对降低NaCl胁迫条件下小白菜体内 MDA 含量均有显著效果,MDA含量与CK组差异不显著。
2.6 PSP对NaCl胁迫下小白菜H2O2含量的影响
由图5可知,与CK组的小白菜相比,盐胁迫使叶片H2O2含量显著增加。添加PSP处理,叶片H2O2含量低于NaCl胁迫的材料。不同浓度PSP均降低了小白菜叶片中H2O2含量。80 mg·L-1PSP处理时,叶片H2O2含量跟正常生长的CK组差异不显著。说明NaCl胁迫下添加PSP可以降低植株内H2O2含量。
2.7 PSP对NaCl胁迫下小白菜脯氨酸积累的影响
根据不同浓度的脯氨酸标准品在520 nm波长下吸光值绘制标准曲线,以OD值为纵坐标,脯氨酸质量浓度(μg·mL-1)为横坐标得出标准曲线y = 0.033 2x + 0.042 5(R2 = 0.990 6)。根據脯氨酸标准曲线计算各试验组小白菜脯氨酸含量如图6所示。与CK相比,NaCl胁迫导致小白菜体内脯氨酸含量显著增加。添加不同浓度的PSP后,脯氨酸含量比NaCl组更高。80 mg·L-1 PSP处理时,小白菜体内脯氨酸含量是NaCl组的1.12倍。
3 讨论与结论
现今,蔬菜种植土壤盐渍化阻碍了蔬菜的无公害生产。提高植物抗盐胁迫的能力是应对盐胁迫的一个有效方法。近年来,人们发现一些外源物质如,NO、赤霉素、微生物多糖和海藻糖等[18-21]均能提高植物抗逆性,它们在缓解植物盐害的同时也不会对植物、土壤和环境带来危害。因此,发掘类似的天然活性物质,对植物耐盐性机理研究以及植物耐盐栽培管理尤为重要。笔者探究了从螺旋藻中提取的螺旋藻多糖是否可以缓解盐胁迫对小白菜生长的影响,为高价值蔬菜的培育提供了新的思路。
众所周知,在高盐环境中,植物根系生长会先受到抑制,进而影响植物地上部分[22]。本试验中,NaCl胁迫使小白菜根长和鲜质量明显减少,然而添加PSP后则缓解了这种生长受抑制的现象。相比于其他剂量处理,80 mg·L-1 PSP处理效果最好。光合作用是影响植物生产力的重要因素,有研究表明NaCl胁迫会导致光合作用速率降低,从而影响植物的生长、产量和品质[23]。叶绿素则是判断光合能力强弱的重要指标。本试验中盐胁迫后添加PSP使得小白菜体内Chl a和Chl b含量均有所提高,表明添加一定浓度的PSP能抑制叶绿体的降解,进而提高光合速率,促进植物生长。SOD、POD和CAT是清除植物体内活性氧,维持活性氧代谢平衡的主要酶系,增强这3种抗氧化酶能够避免盐胁迫后过量的活性氧对植物造成的损伤[24-25]。本研究结果表明,当小白菜受到盐胁迫后,与CK组相比,SOD、POD和CAT酶活性都有不同程度的增强。这可能是在胁迫条件下植物自身清除活性氧能力的激活,植物要在一定程度上忍耐或减缓胁迫对其造成的影响[26]。而添加不同浓度的PSP后,小白菜体内这3种酶活性均显著大幅度提高,对高盐环境下植株生长起到保护作用,这与Tewari等[27]的研究结果类似。植物体内除酶促活性氧清除系统外还有非酶类抗氧化剂,如ASA和CAR等,可以有效抵抗胁迫所造成的伤害[28]。有研究表明,ASA含量的增加可以增强植物抗逆能力,同时改善产品的营养品质。CAR含量的提高不仅可以提升植物捕获光能的能力,也具有帮助植物抵御自由基伤害的抗氧化能力[29]。本试验中,NaCl胁迫显著降低了小白菜体内ASA和CAR含量,而添加PSP后,ASA和CAR含量明显增加甚至高于对照组。综上,施用PSP可以通过提高抗氧化酶的活性以及非酶类抗氧化物质的含量来降低高盐胁迫对小白菜正常生长的影响。
MDA是植物体内膜脂过氧化的产物,其浓度高低可反映胁迫对细胞氧化损伤的程度[30]。本研究发现,NaCl胁迫会显著提高小白菜体内MDA含量,当水培液中添加PSP后,MDA含量明显降低,并与CK组相比无显著差异。这表明PSP可显著降低植物细胞膜脂过氧化,减少活性氧的产生,有效缓解盐胁迫对植株的过氧化伤害。丁振中等[31]研究也表明,NaCl胁迫下,油菜苗叶片中MDA含量增加,而壳寡糖在一定程度上会抑制油菜苗细胞内MDA含量的增加。H2O2是植物体内一类重要的活性氧[32],POD、CAT活性的增强、ASA的积累以及MDA含量的减少,都能够有效清除植物体内的H2O2。本试验结果表明,添加一定浓度PSP后,小白菜体内H2O2含量明显减少,达到正常组水平。
高盐环境下,植物还可以通过积累渗透调节物质来应对胁迫导致的细胞内外渗透压失衡,脯氨酸则是植物细胞内主要渗透调节物质之一[33]。本试验结果显示,盐胁迫下小白菜体内脯氨酸含量增加,说明小白菜可通过自身调节减缓盐胁迫对其造成的影响。有研究表示,施用木寡糖对盐胁迫下小白菜体内脯氨酸含量的积累有显著影响[34],本研究发现,添加PSP多糖后,脯氨酸含量大量的积累,这有助于调节植物细胞渗透势,平衡体内水分,使其在盐渍环境下正常生长。
笔者以水培小白菜为对象,探究PSP是否可以提高其抗盐性。结果表明,NaCl明显影响了小白菜的正常生长,而水培液中添加一定浓度的PSP可显著增加小白菜的根长、鲜质量和叶绿素含量。PSP处理提高了抗氧化酶的活性、非酶类抗氧化物质的含量和脯氨酸含量,抑制了MDA和H2O2的积累。这说明施用天然PSP能提高小白菜抗盐胁迫的能力,这为绿色健康蔬菜的有机生产提供了新知识和方法。
参考文献
[1] 李建国,濮励杰,朱明,等.土壤盐渍化研究现状及未来研究热点[J].地理学报,2012,67(9):1233-1245.
[2] 汪泓洋.玉米苗期耐盐性状的关联分析[D].江苏扬州:扬州大学,2016.
[3] GRAZIA A M,CIARMIELLO L F,PASQUALINA W,et al.Durum wheat roots adapt to salinity remodeling the cellular content of nitrogen metabolites and sucrose[J].Frontiers in Plant Science,2016,7:2035.
[4] ZOU P,LU X,ZHAO H,et al.Polysaccharides derived from the brown algae lessonia nigrescens enhance salt stress tolerance to wheat seedlings by enhancing the antioxidant system and modulating intracellular ion concentration[J].Frontiers in Plant Science,2019,10:48.
[5] 王艳群,彭正萍,薛世川,等.过量施肥对设施农田土壤生态环境的影响[J].农业环境科学学报,2005,24(S1):81-84.
[6] 朱迎春,刘君璞,邓云,等.不同浸种液对盐胁迫下西瓜种子发芽的影响[J].中国瓜菜,2019,32(9):14-17.
[7] LIU H,CHEN X,SONG X,et al.Polysaccharides from grateloupia filicina enhance tolerance of rice seeds (Oryza sativa L.) under salt stress[J].International Journal of Biological Macromolecules,2019,5(1)124:1197-1204.
[8] 郭伟,王庆祥.腐植酸浸种对盐碱胁迫下小麦幼苗抗氧化系统的影响[J].应用生态学报,2011,22(10):2539-2545.
[9] BOSE J,RODRIGO-MORENO A,SHABALA S.ROS homeostasis in halophytes in the context of salinity stress tolerance[J].Journal of Experimental Botany,2014,65(5):1241-1257.
[10] 张强,常瑞雪,胡兆平,等.生物刺激素及其在功能水溶性肥料中应用前景分析[J].农业资源与环境学报,2018,35(2):111-118.
[11] GARCIA A C,BERBARA R L L,FARIAS L P,et al.Humic acids of vermicompost as an ecological pathway to increase resistance of rice seedlings to water stress[J].African Journal of Biotechnology,2012,11 (13):3125-3134.
[12] 李肖宏,劉蕾,梁妍,等.平菇多糖对NaCl胁迫下小白菜种子萌发的影响[J].吉林农业,2018(1):61.
[13] MOSTOFA M G,HOSSAIN M A,FUjITA M.Trehalose pretreatment induces salt tolerance in rice (Oryza sativa L.) seedlings:oxidative damage and co-induction of antioxidant defense and glyoxalase systems[J].Protoplasma,2015,252(2):461-475.
[14] AINAS M,HASNAOUI S,BOUARAB R,et al.Hydrogen production with the cyanobacterium,Spirulina platensis[J].International Journal of Hydrogen Energy,2017,42(8):4902-4907.
[15] 于蕾妍,隋君霞,李华涛,等.螺旋藻多糖与银杏叶提取物复方对四氧嘧啶模型小鼠的降血糖作用[J].中国畜牧兽医,2019,46(7):2176-2182.
[16] 王学奎.植物生理生化实验原理和技术[M].2版.北京:高等教育出版社,2006.
[17] 张志良,翟伟菁,李小方,等.植物生理学实验指导 [M].4版.北京:高等教育出版社,2009.
[18] 赵奕翔.外源NO对盐胁迫下玉竹光合作用及抗氧化保护系统的影响[D].哈尔滨:东北农业大学,2017.
[19] 张丽丽,倪善君,张战,等.外源赤霉素对盐胁迫下水稻种子萌发及幼苗生长的缓释效应[J].中国稻米,2018,24(2):42-46.
[20] 张文平,杨臻,吴佩佳,等.乳酸菌胞外多糖对逆境胁迫下水稻种子萌发及幼苗生长的影响[J].核农学报,2019,33(1):138-147.
[21] 田礼欣,曲丹阳,毕文双,等.海藻糖对盐胁迫下玉米幼苗生长及生理特性的影响[J].草业学报,2017,26(8):131-138.
[22] 王康君,樊继伟,陈凤,等.植物对盐胁迫的响应及耐盐调控的研究进展[J].江西农业学报,2018,30(12):31-40.
[23] 赵莹,杨克军,赵长江,等.外源糖调控玉米光合系统和活性氧代谢缓解盐胁迫[J].中国农业科学,2014,47(20):3962-3972.
[24] 杨小环,赵维峰,孙娜娜,等.外源水杨酸缓解低温胁迫对玉米种子萌发和早期幼苗生长伤害的生理机制[J].核农学报,2017,31(9):1811-1817.
[25] TURK H,ERDAL S.Melatonin alleviates cold induced oxidative damage in maize seedlings by up-regulating mineral elements and enhancing antioxidant activity[J].Journal of Plant Nutrition and Soil Science-Zeitschrift fuer Pflanzenernaehrung und Bodenkund,2015,178(3):433-439.
[26] LIANG Y C,CHEN Q,LIU Q,et al.Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.)[J].Journal of Plant Physiology,2003,160(10):1157-1164.
[27] TEWARI S,ARORA N K.Multifunctional exopolysaccharides from Pseudomonas aeruginosa,PF23 involved in plant growth stimulation,biocontrol and stress amelioration in sunflower under saline conditions[J].Current Microbiology,2014,69(4):484-494.
[28] 罗银玲,宋松泉,兰芹英.酶促和非酶促抗氧化系统在玉米胚脱水耐性获得中的作用[J].云南植物研究,2009,31(3):253-259.
[29] GILL S S,TUTEjA N.Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants[J].Plant Physiology and Biochemistry,2010,48(12):909-930.
[30] 张小艾,李名扬,汪志辉,等.重金属及盐碱对二月兰幼苗生长和生理生化的影响[J].草业学报,2013,22(2):187-194.
[31] 丁振中,曾哲灵,龚劲松,等.NaCl胁迫下壳寡糖对油菜苗生长的影响[J].生物化工,2018,4(3):29-33.
[32] PETROV V D,VAN B F .Hydrogen peroxide-a central hub for information flow in plant cells[J].AoB Plants,2012,4(18):1-13.
[33] 范海霞,郭若旭,辛国奇,等.外源褪黑素对盐胁迫下芦苇幼苗生长和生理特性的影响[J].中国农业科技导报,2019,21(11):51-58.
[34] 陈薇薇.木寡糖促进小白菜生长及抗盐胁迫效应研究[D].武汉:华中农业大学,2016.
