不同性别类型黄连木的花序类型与分布及亲缘关系1)
2020-05-13阮福娜白倩陈一帆刘阳杨子玥苏淑钗
阮福娜 白倩 陈一帆 刘阳 杨子玥 苏淑钗
(省部共建森林培育与保护教育部重点实验室(北京林业大学),北京,100083)
黄连木(Pistaciachinensis),又称中国黄连木,别名楷树、楷木、药木和黄连茶等,属于漆树科(Anacardiaceae)黄连木属(PistaciaL.)植物。黄连木是落叶乔木,在我国分布广泛,黄连木的最佳适生区位于太行山区的冀豫晋陕等集中地域,且河北邯郸地区分布最多[1-2]。以往一直认为黄连木属植物均为雌雄异株[3-4],且自然分布的黄连木雌株少雄株多,后来陆续在阿月浑子(P.veraL.)、大西洋黄连木(P.atlanticaDesf)发现雌雄同株树[5-9]。2008年林业专家在河南省林州市进行考查时,发现了7棵树龄30 a左右且分布相对集中的雌雄同株中国黄连木[10]。王文浩等[11]在河北省唐县发现了23棵雌雄同株黄连木,进一步研究表明,雌雄同株黄连木的配子体均具有育性,雌雄异株的雌株、雄株和雌雄同株植株间具有杂交亲和性。黄连木是一种重要的木本油料作物,为了提高其果实产量,需要合理的雌雄树配比,如果能开发雌雄同株黄连木资源,将有利于节约土地资源与人力,促进丰产。因此,深入开展雌雄同株黄连木的研究,不仅对其种质资源的鉴定和保存具有重要意义,而且在生产上具有重要应用前景。
SSR(Simple sequence repeats)标记是共显性标记,具有操作简单、稳定性和重复性好、检测结果多态性丰富、遗传信息量大等优点[12],目前已被用于黄连木属植物的遗传多样性和亲缘关系分析、品种鉴定、分类和遗传连锁图谱构建等方面[13-16]。对雌雄同株黄连木的研究主要集中在生物学特性、雌雄配子体育性、花粉活性、结果习性、变异情况以及不同枝营养元素动态差异方面[11],并未涉及雌雄同株黄连木的亲缘关系研究。因此,本研究在河南林州和河北唐县扩大调查范围,寻找更多的雌雄同株树,并探究雌雄同株树上不同性别类型的基因型稳定性。同时采用14个SSR标记对4个地区的黄连木进行了亲缘关系鉴定,为雌雄同株黄连木的起源和演化提供分子水平的理论依据,对科学有效制定雌雄同株黄连木资源保护策略具有重要意义。
1 材料与方法
1.1 材料
在河南马鞍山、河南西齐、河北老姑和河北杨家庵4个地区,实地调查寻找雌雄同株黄连木,通过文本、数据和图像记录4个地区的地理位置、人为干扰程度,同时记录每棵雌雄同株树的花序类型和分布、雌雄同株树的分布比例。
通过调查,在河北和河南4个地区共收集了140个样本,即70棵雌雄同株树,54棵雌树和16棵雄树(表1)。每个地区雌雄同株和雌雄异株树数量基本相等,同时每棵雌雄异株树间隔50 m以上。在河北杨家庵地区,当地居民认为黄连木雄树不能结实,与雌树相比没有价值,因此被大量砍伐。而河南西齐地区雄树太少,因此这两个地区都没有采集雄树样本。基于基因型稳定性研究,在这4个地区的70棵雌雄同株黄连木中随机选取9棵,采集每棵雌雄同株树上的不同性别类型的花序(包括雌花序、雄花序、混合花序和两性花),立即放入干燥的自封袋中带回实验室,液氮速冻后,放在冰箱中-80 ℃保存。
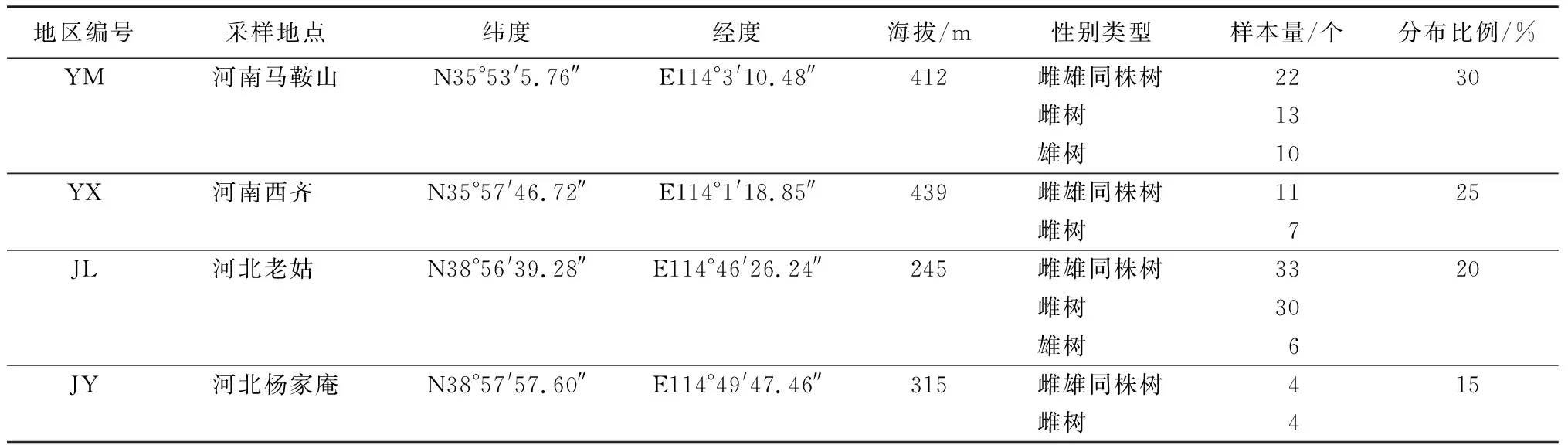
表1 4个黄连木地区基本情况收集点的信息
1.2 方法
1.2.1 基因组DNA的提取
基因组DNA的提取使用新型植物基因组DNA快速提取试剂盒(北京艾德莱生物科技有限公司)。DNA纯度和质量浓度使用超微量紫外分光光度计检测,经1.0%的琼脂糖凝胶电泳检测DNA完整性后,将其稀释至30 mg/L,-20 ℃保存备用。
1.2.2 PCR扩增
本研究基于阿月浑子的SSR原始扩增条件[17],建立了适合中国黄连木的扩增反应体系:15 ng模板DNA,5 μL的PCR MIX混合液(Taq DNA聚合酶、dNTPs、MgCl2、反应缓冲液、PCR反应的增强剂和优化剂以及稳定剂),正反引物各0.25 μL,ddH2O 4 μL,共10 μL的反应体系。
SSR扩增程序分为两个连续步骤:第一步在94 ℃下初始变性3 min,然后进行23个循环,每个循环在94 ℃变性30 s,54~60 ℃退火45 s和72 ℃伸长60 s;第二步包括10个循环,每个循环94 ℃变性30 s,52 ℃退火45 s和72 ℃延伸60 s,最后72 ℃延伸5 min。
1.2.3 引物筛选
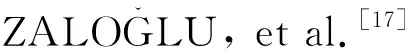
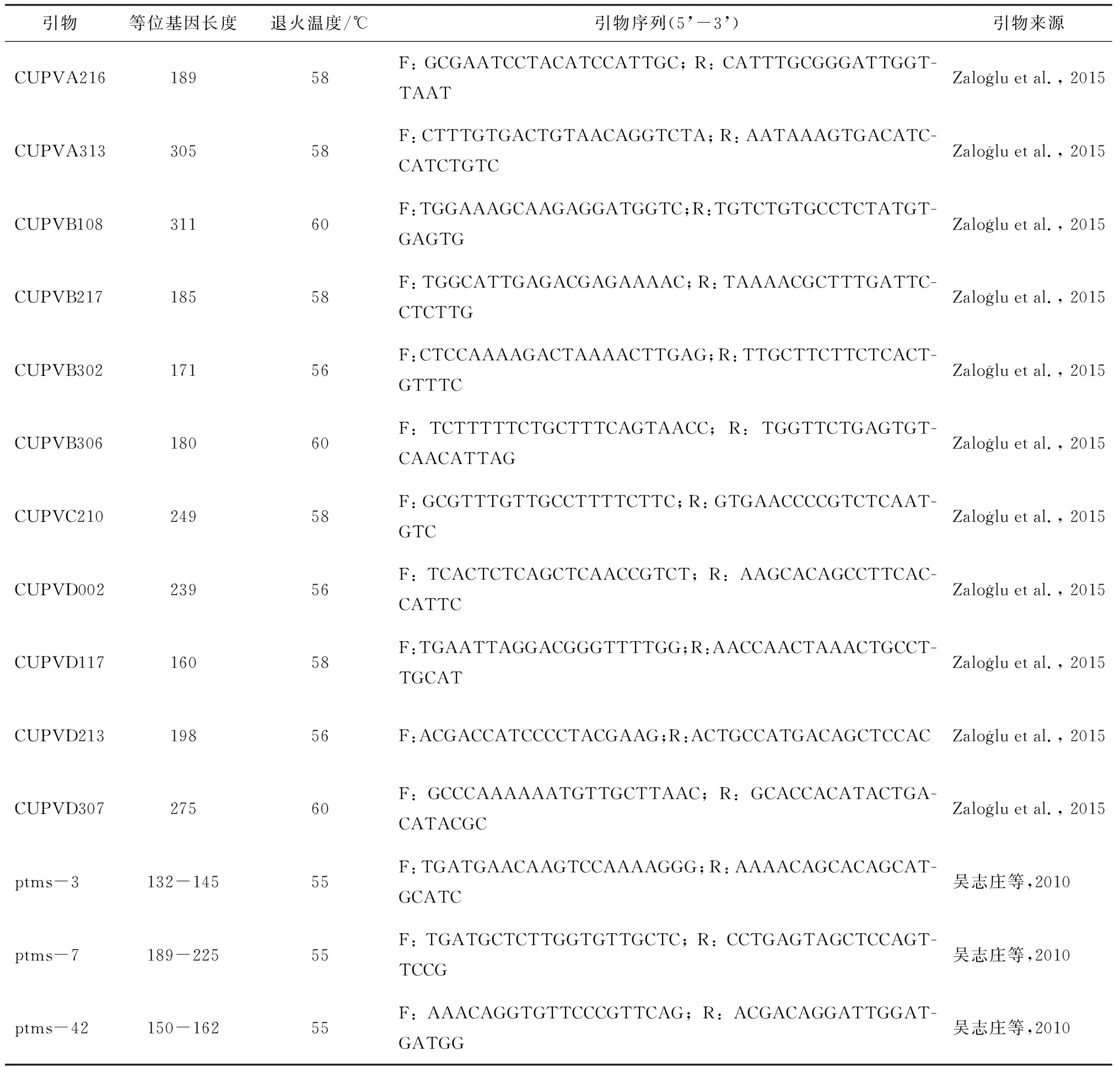
表2 14对SSR引物的序列特征和来源
1.2.4 扩增产物的分离与检测
PCR扩增产物通过8%的非变性聚丙烯酰胺凝胶(5×TBE溶液3.7 mL、30%丙烯酰胺溶液母液8.8 mL、ddH2O 23.1 mL、TEMED 24 μL、10% APS(过硫酸铵)259 μL)电泳分离,150 V恒压。银染后观察、拍照。
1.3 数据分析
根据电泳结果,对清晰的条带进行统计,从大到小依次标记为A、B、C…,纯合基因型用相同字母标记,如AA、BB、CC…;杂合基因型用不同字母标记,如AB、AC、BC…,并建立数据矩阵。用NEI[19]的计算法估计遗传距离(GD)和遗传一致度(GI)。根据遗传距离和遗传一致度,用NTSYS-pc2.1[20]软件对4个地区140个黄连木个体进行UPGMA聚类分析和主成分分析。
2 结果与分析
2.1 雌雄同株样本花序类型和分布
根据种质资源调查,在河南马鞍山、河南西齐、河北老姑和河北杨家庵4个地区分别发现了22、11、33、4棵雌雄同株黄连木。部分雌雄同株树着生4种不同类型的花序,包括雌花序、雄花序、混合花序和两性花。在河北老姑和杨家庵地区,黄连木雌树树龄大约60 a,有几棵树大约20 a或者超过100 a,而雄树大约20 a,雌雄同株树的树龄与雌树的树龄相似[21]。在河南马鞍山地区,雌树、雄树和雌雄同株树的树龄大约10~30 a,有几棵树大约是50 a。在河南西齐地区,雌树、雄树和雌雄同株树的树龄大约是50 a。
河南和河北4个野生黄连木地区雌雄同株树的花序类型和分布分别见表3和表4。可以看出在河南马鞍山地区,着生4、3、2种花序类型的雌雄同株树分别有7、2、13株;在河南西齐地区,着生4、3、2种花序类型的雌雄同株树分别有4、3、4株。在河北老姑地区,着生4、3、2种花序类型的雌雄同株树分别有7、15、11株;在河北杨家庵地区,没有存在4种花序类型的雌雄同株树,存在3、2种花序类型的雌雄同株树分别有1、3株。在河南马鞍山和西齐地区,雌雄同株树上两性花的数量多于混合花序的数量,雌花序和雄花序的数量基本相等;而在河北老姑和杨家庵地区,混合花序的数量多于两性花的数量,雌花序的数量多于雄花序的数量。
2.2 SSR标记的基因型稳定性
SSR标记位点ptms-7呈现的9棵雌雄同株黄连木及每棵雌雄同株树不同性别类型花序的DNA图谱如图1所示。结果表明不同的雌雄同株黄连木的gDNA图谱存在差异,而同一株雌雄同株树不同性别类型花序的gDNA图谱无差异,即雌雄同株树上无特殊标记的DNA条带。
表3 河南马鞍山和西齐地区33个雌雄同株样本花序类型和分布

序号样本编号花序类型和分布1YMMo-12种类型;较多的雌花序和非常少的雄花序2YMMo-23种类型;较多的雄花序和两性花,较少的雌花序3YMMo-32种类型;雄花序∶雌花序=6∶44YMMo-44种类型;较多的雌花序和两性花,较少的雄花序和混合花序5YMMo-52种类型;雄花序∶雌花序=7:36YMMo-62种类型;较多的雌花序和较少的雄花序7YMMo-74种类型;较多的雌花序和雄花序,较少的混合花序和两性花8YMMo-84种类型;较多的雌花序和两性花,较少的雄花序和混合花序9YMMo-94种类型;较多的混合花序和两性花,较少的雌花序和雄花序10YMMo-102种类型;雄花序∶雌花序=2∶811YMMo-112种类型;雄花序∶雌花序=7∶312YMMo-124种类型;较多的混合花序和两性花,较少的雌花序和雄花序13YMMo-132种类型;雄花序∶雌花序=3∶714YMMo-144种类型;较多的雌花序和雄花序,较少的混合花序和两性花15YMMo-152种类型;雄花序∶雌花序=1∶916YMMo-162种类型;较多的雄花序和非常少的雌花序17YMMo-172种类型;雄花序∶雌花序=6∶418YMMo-182种类型;雄花序∶雌花序=6∶419YMMo-192种类型;雄花序∶雌花序=1∶120YMMo-204种类型;较多的雌花序和两性花,较少的雄花序和混合花序21YMMo-212种类型;较多的雄花序和非常少的雌花序22YMMo-223种类型;较多的雌花序和两性花,较少的雄花序23YXMo-13种类型;较多的雄花序和两性花,较少的混合花序24YXMo-22种类型;雄花序∶雌花序=1∶125YXMo-33种类型;雄花序∶雌花序∶两性花=1∶1∶126YXMo-42种类型;雄花序∶雌花序=4∶627YXMo-52种类型;较多的雌花序和非常少的雄花序28YXMo-64种类型;较多的雄花序和混合花序,较少的雌花序和两性花29YXMo-73种类型;较多的两性花,较少的雌花序和雄花序30YXMo-82种类型;雄花序∶雌花序=7∶331YXMo-94种类型;较多的雌花序和两性花,较少的雄花序和混合花序32YXMo-104种类型;较多的雄花序和混合花序,较少的雌花序和两性花33YXMo-114种类型;较多的雄花序和两性花,较少的雌花序和混合花序
注:Mo为雌雄同株树。

M.50 bp DNA Ladder;A.雌花序;B.雄花序;C.混合花序;D.两性花。
图1 引物对ptms-7测定的9棵雌雄同株黄连木及每棵雌雄同株树不同性别类型花序的SSR等位基因
2.3 SSR标记的聚类分析
利用基于Nei’s遗传距离数据矩阵的UPGMA方法构建了4个黄连木地区的聚类图。为了更清楚的分析黄连木不同个体之间的亲缘关系,根据聚类结果选择一个较大的遗传相关系数作为阈值。
表4 河北老姑和杨家庵地区37个雌雄同株样本花序类型和分布
注:Mo为雌雄同株树。
图2以遗传相关系数0.86为阈值,河南马鞍山和西齐地区63个黄连木样本的相关统计数据如下:YMMo-6和YMM-7的遗传相似性系数最大,亲缘关系最近,YXMo-9和YMF-6、YMMo-1和YMM-10、YMMo-20和YMMo-22、YMMo-11和YMF-4、YMMo-13和YMMo-16的亲缘关系逐渐变远,而YMMo-14和YMMo-17的遗传相似性系数较小,说明这两个个体在63个黄连木样本中亲缘关系相对较远。在这7个聚类中,雌雄同株树与雌雄同株树、雌树、雄树样本聚类数分别为3、2、2,聚类结果表明在河南马鞍山和西齐地区不同性别类型的黄连木均表现出较近的亲缘关系。图3以遗传相关性系数0.81为阈值,河北老姑和杨家庵地区77个黄连木样本的相关统计数据如下:JLMo-11和JLF-30的遗传相似性系数最大,亲缘关系最近;JLMo-4和JLF-16、JLMo-16和JLMo-30、JLMo-29和JYF-3、JLMo-12和JYF-4、JLMo-32和JLF-13、JLMo-27和JLF-29、JLMo-15和JLMo-26、JLMo-8和JLMo-24、JLMo-10和JLF-2的亲缘关系逐渐变远,而JLMo-20和JLF-8的遗传相似性系数较小,说明这两个个体在77个黄连木样本中亲缘关系相对较远。在这11个聚类中,雌雄同株树与雌雄同株树、雌树样本聚类数分别为3、8,聚类结果表明在河北老姑和杨家庵地区雌雄同株黄连木与雌雄同株树、雌树表现出较近的亲缘关系。
河南和河北4个地区140个黄连木样本的UPGMA聚类分析见图4。以遗传相关性系数0.86为阈值,140个黄连木样本的相关统计数据如下:JLMo-11和JLF-30的遗传相似性系数最大,亲缘关系最近;JYMo-4和JLF-21、JLMo-4和JLF-16、YMMo-6和YMM-7、YMMo-16和JLMo-22、YXMo-9和YMF-6、YMMo-1和YMM-10、YMMo-13和JLMo-26、JLMo-24和JYMo-2、YMMo-20和YMMo-22、JLMo-16和JLF-24、JLMo-29和JYF-3、YMMo-14和YMMo-17、YMMo-11和YMF-4的亲缘关系逐渐变远,而JLMo-27和JLMo-30的遗传相似性系数较小,说明这两个个体在140个黄连木样本中亲缘关系相对较远。在这15个聚类中,雌雄同株树与雌雄同株树、雌树、雄树样本聚类数分别为6、7、2,聚类结果表明雌雄同株黄连木与雌雄同株树、雌树与雄树均表现出较近的亲缘关系。雌雄同株树不仅可以和当地的黄连木聚类,也可以与其他地区的黄连木聚类。
3 结论与讨论
太行山区是我国黄连木主要分布区[22]。近些年在河南林州和河北唐县分别发现了7棵、23棵雌雄同株黄连木[10-11],改变了黄连木属雌雄异株的观点。中国黄连木亲缘关系的研究主要集中在对黄连木不同居群或不同种源亲缘关系[18,23-24],并未涉及雌雄同株黄连木。
黄连木属植物的遗传关系与地理位置与环境因素密切相关[18,23,25-27],如郝丽娟[23]基于SSR分子标记对中国黄连木地区进行亲缘关系分析,发现相邻省份的黄连木居群具有较近的亲缘关系。吴志庄等[18]对11个黄连木群体的18个表型性状进行聚类分析,发现来源地较近的群体亲缘关系较为密切,而来源地不同的群体遗传差异相对较大。但在本研究中,黄连木的亲缘关系与来源地无明显的相关性和地域性,如河南和河北4个地区的黄连木均表现出较近的亲缘关系,分析其差异可能源于本研究的4个地区,同属太行山东麓,地域相近,空间距离小(可能属同一起源),较为集中且跨度较小。
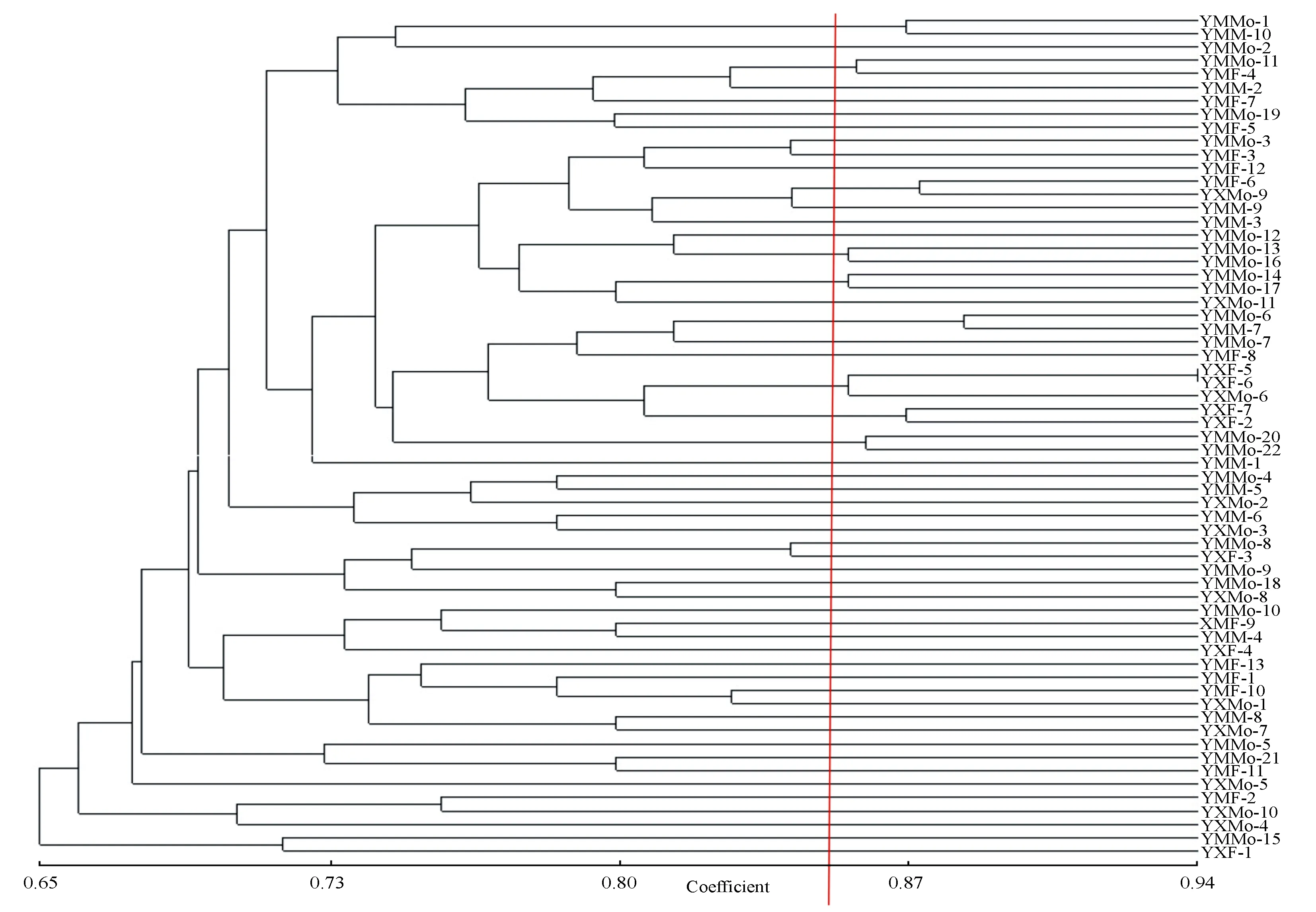
图2 河南马鞍山和西齐地区63个样本的聚类分析树状图
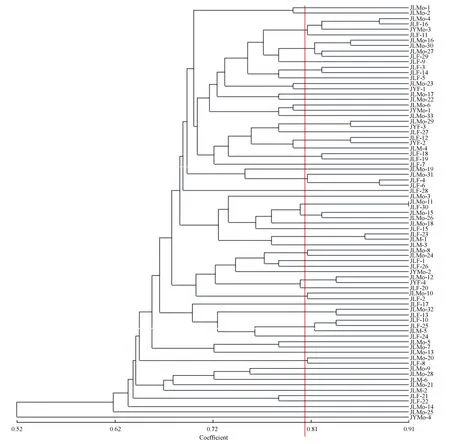
图3 河北老姑和杨家庵地区77个样本的聚类分析树状图
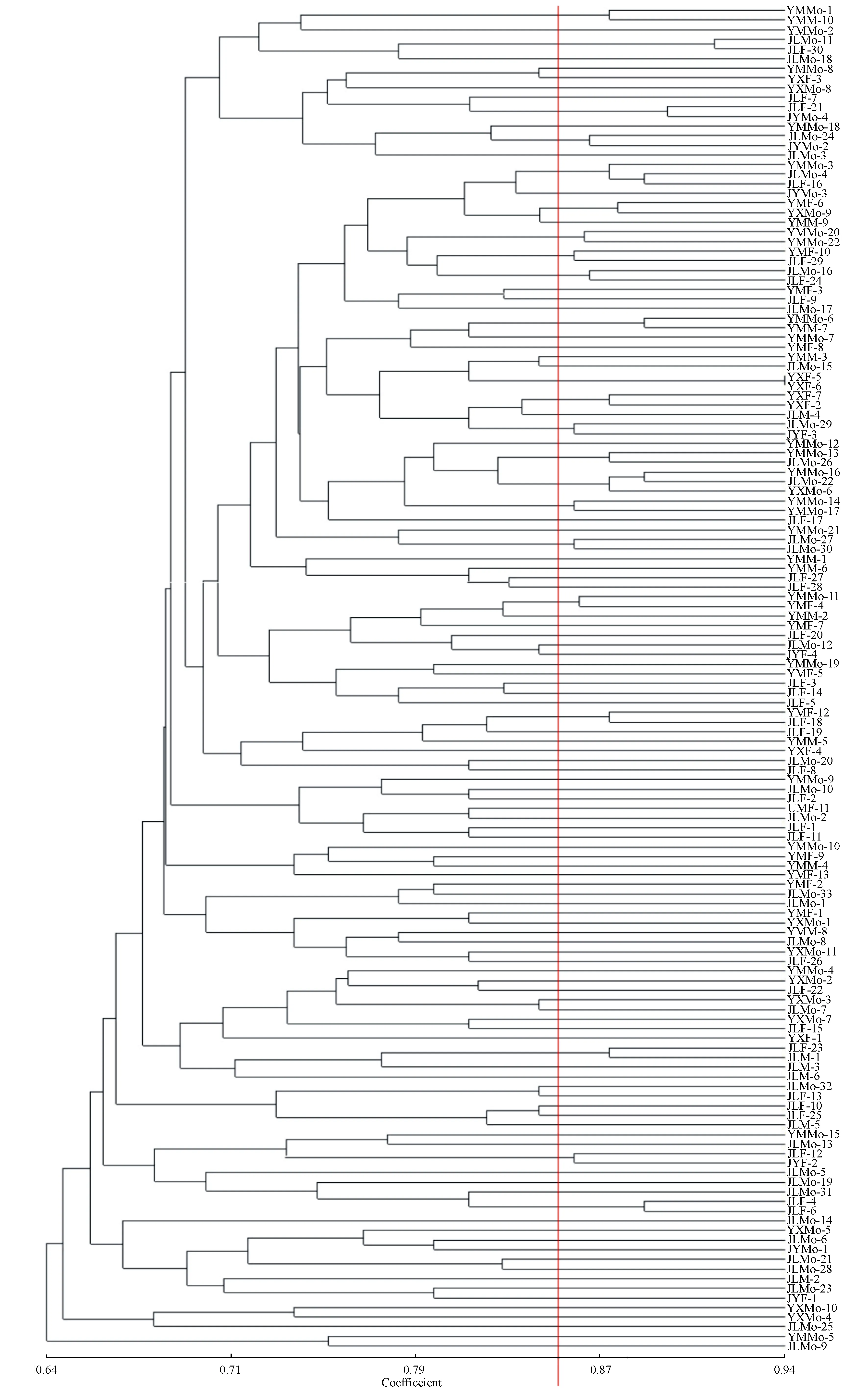
图4 河南和河北四个地区140个样本的聚类分析树状图
在河北两个黄连木地区,雌雄同株与雌树表现出较近的亲缘关系,与雄树的亲缘关系有待进一步确定。在调查种质资源中发现,雄树黄连木的树龄较小,大约为20 a,均为实生树,可能是雄树黄连木被当地人大量砍伐造成。在河南两个黄连木地区,3种性别类型均表现出较近的亲缘关系。这些结果可能是由于河南和河北4个地区黄连木雌雄同株树、雌树和雄树具有共同的起源,也可能是由于雌雄同株黄连木可以与当地的雌树或雄树发生相互转化。
BAI, et al.[21]发现雌雄同株黄连木的性别表现是不稳定的,除了没有雌花变成雄花外,雌花序、雄花序、混合花序或两性花在连续的1 a内可以转变成其他性别类型。但在河南省进行种质资源调查时,许多当地人表示有雌树变成雄树的情况出现,对此还需要进一步证实。本研究发现同一棵雌雄同株树上不同性别类型花序的DNA条带没有差异,说明性别类型在同一株树上的可变表达不是DNA变异造成的,而是基因表达调控引起的发育结果。因此今后的性别稳定性研究工作应从表型水平进行。
