肉品中的寡肽及其呈味机制研究进展
2020-05-08罗玉龙李文博杨致昊赵丽华田建军
窦 露,罗玉龙,刘 畅,李文博,杨致昊,靳 烨,赵丽华,田建军,苏 琳
(内蒙古农业大学食品科学与工程学院,内蒙古呼和浩特 010018)
肉品是人类摄取蛋白质的重要来源,滋味是评价肉品品质的重要指标。研究发现滋味物质的呈现主要通过呈味肽、游离氨基酸、核苷酸和有机酸等。其中蛋白质水解或自溶产生的活性氨基酸和肽是滋味的主要贡献者,赋予食品甜、酸、苦、咸和鲜,这些味道刺激味觉受体细胞产生反应并通过电信号的形式传入大脑最终产生味觉[1-2]。
随着人们生活水平的提高,单一的味觉特性已不能满足人们的需求,因此研究的重点也逐渐转向肽类组分。呈味肽中的寡肽主要包括鲜味肽、甜味肽、苦味肽、酸味肽、咸味肽及浓厚感肽,这六种特征滋味肽共同改善了肉品的滋味。国内外学者已针对各类寡肽的检测、分离鉴定展开研究,并且随着科技的进步,已有学者利用计算机技术模拟了肽类分子与味觉受体之间的相互作用,这为滋味的改良及产品的开发开拓了广阔的研究前景。然而随着细胞生物学、神经科学的发展以及滋味受体的研究和新型受体的完善,国外对于几种寡肽及呈味机制的研究已有最新报道。因此本文通过对肉品中寡肽(鲜味肽、浓厚感肽、苦味肽、甜味肽)的呈味特征、呈味机制以及影响寡肽的呈味因素进行综述,旨在深入探究肉品呈味的机制,为健康、绿色肉制品的研制提供参考。
1 寡肽概述
呈味肽可分为多肽和寡肽,其中寡肽是肽链上氨基酸残基数在10个以内的蛋白质片段。许多特定的寡肽已被鉴定为苦味、咸味、酸味以及鲜味,在发酵和腌制产品的滋味产生中起重要作用,它主要通过内源蛋白水解酶(二肽基肽酶和组织蛋白酶B)分解肉中蛋白质产生[3]。在成熟火腿中,滋味的形成与肽类密切相关,二肽中的肌肽对火腿滋味起到很强的缓冲作用,八肽中的Lys-Gly-Asp-Glu-Glu-Ser-Leu-Ala能形成类似味精的滋味;并且寡肽的含量和比例能决定火腿的滋味,形成鲜味、甜味、酸味和苦味等。Sentandreu等[4]、Aristory等[5]通过对西班牙干熏火腿的研究,发现分子质量小于3000 Da的肽呈现多种滋味,其中分子量在1500~1700 Da的肽具有鲜味,而1800 Da左右的肽呈现苦味。国内王娟[6]在成熟火腿中鉴定出寡肽(二肽、三肽和四肽),其中Ser-Glu、Val-Glu、Leu-Glu、Glu-Glu-Glu和Glu-Gly-Ser呈现出鲜味,而Pro-Leu、Leu-Lys呈苦味。因此寡肽在肉制品的呈味中至关重要,且不同氨基酸组成及序列极大程度地影响肉品的呈味。
2 寡肽的滋味特征
肉制品中的内源蛋白水解能导致寡肽的不断积累,这在一定程度上赋予肉品良好的滋味。寡肽的味感取决于其组成氨基酸的原有味感,如含有谷氨酸(Glu)残基的寡肽能够提高肉的鲜味[7]。氨基酸主要呈现酸、甜、苦、鲜等滋味,其中甘氨酸(Gly)、丙氨酸(Ala)、丝氨酸(Ser)以及脯氨酸(Pro)呈甜味,天冬氨酸(Asp)和Glu则具有鲜甜味,亮氨酸、苯丙氨酸为苦味[8]。呈味肽的味型取决于氨基酸,包括酸、甜、苦、咸和鲜以及浓厚感。其中酸味和咸味是由带电末端或带电侧链的存在而产生。
2.1 鲜味寡肽
鲜味,被认为是第五种基本味道,在肉制品中起重要作用,具有鲜味的氨基酸主要为Glu和Asp。鲜味成分可调节胃肠功能,并通过减少饮食中的钠摄入量来降低成人中风和冠心病的风险。鲜味肽最初是Yamasaki等[9]用木瓜蛋白酶水解牛肉后得到的一种序列为Lys-Gly-Asp-Glu-Glu-Ser-Leu-Ala的八肽,也称为“牛肉风味肽(BMP)”,因肽链中含有Glu和Asp组成的酸性基团,起到了增强肉鲜味的作用。Take等[10]认为包括Glu在内的游离氨基酸可改善鸡肉的鲜味强度[10]。Dang等[11]也从火腿中分离出具有强烈鲜味的六肽(Cys-Cys-Asn-Lys-Ser-Val)和七肽(Ala-His-Ser-Val-Arg-Phe-Tyr)。
鲜味肽的感官特性由组成氨基酸的原有味感决定,一般来说,Glu、Asp、Gln、Asn相互作用或与Thr、Ser、Ala、Gly、Met相互结合形成的肽链呈鲜味,如Glu-Glu、Glu-Asp、Glu-Ser、Glu-Thr、Glu-Gln-Gln、Ser-Glu-Gln等[12]。但Yu等[13]的研究也证明鲜味不仅仅是基于肽链中鲜味氨基酸的存在,由非鲜味氨基酸与肽键结合组成的短肽也可呈现鲜味,例如VPY(Val-Pro-Tyr)、TAY(Thr-Ala-Tyr)、AAPY(Ala-Ala-Pro-Tyr)和GFP(Gly-Phe-Pro),虽具有鲜味增强效果但本身不含鲜味氨基酸,而含有苦味氨基酸。鲜味的产生主要存在3种类型的相互作用,分别为肽与肽的相互作用,肽与核苷酸的相互作用,肽与阳离子的相互作用。Tamura等[14]研究了含有N-末端二肽(Lys-Gly)、酸性肽(Asp-Glu-Glu)、C-末端三肽(Ser-Leu-Ala)和咸味二肽(Orn-β-Ala)的混合物味道,发现所有组合都产生了鲜味,且具有与BMP相同的呈味强度。
协同作用是鲜味的最独特特征,主要通过对自身味道的调控以及与其他物质协调调控来实现。Yamaguchi等[15]和Martin等[16]发现IMP和GMP可以增强Glu和Asp的鲜味,极大程度地增加肉汤的鲜美程度,且二者的增味强度高于MSG。鲜味肽和谷氨酸钠可产生良好的协同作用,究其原因,可能是MSG的加入使得T1R3结合腔的大小从534.125A3扩大到1135.75A3,有利于小肽与T1R3更充分地结合[17]。此外,鲜味还可以和苦味相互作用,从而降低苦感。含有谷氨酰肽的鲜味活性寡肽,包括Glu-Asp、Glu-Glu、Glu-Ser以及Glu-Glu-Glu,可抑制火腿生产过程中蛋白质水解产生的苦味[18]。
2.2 浓厚感肽
浓厚味道由舌头上的钙通道引起,这些钙通道诱导Ca2+受体细胞的反应,从而增强咸味,甜味和鲜味的强度[19]。浓厚感肽(kokumi)本身没有味道活性,但可以通过调节从味觉受体到大脑的信号转导来增强其他化合物的味觉强度,从而改良风味、增加食品味道的持久性、复杂性和饱满性[20]。kokumi的主要活性成分为含硫组分,包括甲基半胱氨酸亚矾和γ-谷酰基丙烯基半胱氨酸等。图1为几种典型的kokumi分子结构[19]。在浓厚感肽中,谷胱甘肽是第一个被鉴定为具有浓厚感活性的肽类组分,主要应用于肉类、海鲜类食品中,可延长食品的保鲜期,强化令人喜爱的口感[19]。kokumi的呈味效果会受单个氨基酸的影响,但这并不能决定总体呈味效果。许多kokumi肽的末端含有疏水氨基酸,然而这些肽并不呈现苦味,反而呈现出浓厚感、持久性及绵延感等风味特性[21]。浓厚感肽广泛存在于发酵肉制品中,Suzuki等[22]已经从发酵火腿中鉴定出具有浓厚味的二肽和三肽,并且发现可通过细菌γ-谷氨酰转肽酶的转肽反应来降低氨基酸的苦味。

图1 几种典型的Kokumi分子结构
浓厚感肽也具有协同增味的作用,能有效协调5种基本滋味,增强厚度和持久的味觉。来源于鸡肉中的浓厚感肽Glu-Glu、Glu-Val、Ala-Asp-Glu、Ala-Glu-Asp、Asp-Glu-Glu和Ser-Pro-Glu可以增强5′-肌苷一磷酸的呈味作用[23]。Takashi等[24]使用描述性分析研究了添加γ-Glu-Val-Gly对鸡清汤的感官特征影响,在99%的置信水平下,含有γ-Glu-Val-Gly的鸡肉具有比对照样品更强的“口腔涂层”特征。这些数据表明,浓厚感肽γ-Glu-Val-Gly可以增强鸡清汤中的鲜味和浓厚感特征。几种γ-谷氨酰二肽也具有浓厚味道活性,如Glu-Tyr与成熟的火腿味有关。它们的优点在于溶解度高于α-谷氨酰肽,并且能够抵抗肽酶的水解[25]。
γ-Glu-Val-Gly被允许用作食品添加剂并逐渐商业化,有研究者开发出一种利用细菌酶(地衣芽孢杆菌和解淀粉芽孢杆菌GGT的蛋白酶)从蛋白质水解产物中生产kokumi(γ-Glu-Val-Gly)调味料的新方法[26]。Yamasaki等[9]将牛肉进行热水浸提预处理,并用凝胶过滤色谱、离子交换和电泳的方法,分离出序列为Lys-Gly-Asn-Glu-Ser-Leu-Ala的浓厚感肽,并利用爱德曼降解、羧肽酶解技术进行鉴定。陶正清等人采用凝胶色谱过滤从盐水鸭中分离出序列为Gly-Pro-Asp-Pro-Leu-Arg-Tyr-Met、Asp-Pro-Leu-Arg-Tyr-Met、Val-Val-Thr-Asn-Pro-Ser-Arg-Pro-Trp的浓厚感肽[27]。浓厚感肽赋予肉品绵润、回味悠长的口感,然而目前对于浓厚感和鲜味的概念仍容易混淆,因此,是否可以通过确定一个标准参考来辨别二者,还有待进一步研究。
2.3 苦味寡肽
苦味肽以二肽、三肽、六肽和八肽的形式广泛存在于肉制品中,目前已经从成熟的火腿中分离出多种苦味二肽,比如Ile-Val、Leu-Gly、Ile-Asp和Pro-Leu等[4]。党亚丽等[28]发现金华火腿提取物中大部分是相对分子质量<1000 Da的寡肽,包括一些与苦味相关的疏水性小分子肽。肽链中疏水氨基酸的种类、含量、序列、位置及空间结构都与苦味密切相关。Ishibashi等[29]发现C末端中L-构型的疏水性氨基酸的数量增加时,苦味更强烈。研究表明疏水性氨基酸在肽链中间比在肽链两端更苦,这是因为当疏水性氨基酸处于肽链中间时,小肽结构能更充分地与苦味受体结合,从而引发苦感[30]。脯氨酸(Pro)的存在也使得肽呈现苦味,肽结构中含有Pro的肽更有利于与苦味受体结合[31]。此外,圆二色性分析进一步揭示了具有相似苦味强度的肽具有类似的二级结构特征。因此,肽的苦味不仅取决于肽序列,还取决于它们的空间结构。
一般来说,苦味肽具有两个功能单元,即“结合单元”(BU)和“刺激单元”(SU),如图2[32]。苦味肽的疏水性侧链能够为苦味受体提供BU,而SU作为苦味的决定性部位,由庞大的碱性或疏水性基团组成,二者的存在引发苦味肽与苦味受体的结合,其诱导神经信号级联反应。此外,苦味强度主要取决于苦味受体的疏水识别区域,当两种功能单元(BU和SU)在苦味肽的空间构象中共存时,苦味往往变得更明显。

图2 苦味肽与苦味受体结合
肌肽(β-丙氨酰-1-组氨酸)和鹅肌肽(β-丙氨酰-1-甲基组氨酸)都具有很强的缓冲作用,通过调节肌肉pH进而影响离子成分,最终对滋味产生影响。此外,二肽还具有其他作用,肌肽、鹅肌肽和巴林碱(β-丙氨酰-1-甲基组氨酸),均对食品感官属性和储存特性有一定贡献。鹅肌肽和肌肽通过破坏不饱和醛,可降低肉类酸败程度,在赋予浓郁味道的同时还可抑制脂质氧化[2]。Decker等[33]将不同浓度的肌肽置于绞碎的猪肉中,并与2,6-二叔丁基-4-甲基苯酚(BHT)、三聚磷酸钠以及α-生育酚等抗氧化剂做比较,结果表明肌肽抗氧化效果最好,且具有一定的护色作用。国内已有L-肌肽的合成工艺,技术原理为:β-Ala→Z-β-Ala→Z-β-Ala-Osu→Z-β-Ala-His→β-Ala-His,其纯度可达99%。同时,Jakub等[34]发现肌肽的合成和含量变化可能受到肌肽合成酶和肌肽氮甲基转移酶等基因的影响。
苦味给人一种非愉悦的口腔感觉,因此降低苦味来提高食品的感官品质以及消费者的接受度成为研究热点。研究表明可通过γ-谷氨酰转肽酶(GGT)将苦味氨基酸转化为γ-谷氨酰基衍生物,这种方法不仅可以降低苦味氨基酸Phe、Val、Leu和His的浓度,还产生具有浓厚味的γ-谷氨酰肽。已经在几种大豆蛋白衍生的肽中观察到C-末端的亮氨酸残基导致苦味,用羧肽酶处理可以使苦味强度显著降低[35]。也有研究表明谷氨酸低聚物如Glu-Leu、Pro-Glu和Val-Glu可以作为苦味掩蔽剂来抑制鹅肌肽和肌肽的苦味[36]。2018年Zhang等[37]最新研究发现牛肉蛋白质衍生的肽可以作为苦味受体T2R4的阻滞剂,从而抑制苦味物质的活化,并且已经确定多种苦味阻断化合物的结构特性。目前只发现了13个针对特定T2R并可在受体水平上起作用的苦味阻断剂,然而这些阻断剂不能阻断所有T2R,它们仅与10种T2R亚型相互作用[38]。
2.4 甜味寡肽
人习惯将蔗糖、果糖和其它糖类引起的味觉称为甜味。随着糖尿病等案例的不断增加,人们对新型低热量甜味剂的需求也逐渐增大。最著名的甜味肽是L-天冬氨酰-L苯丙氨酸甲酯,也就是阿斯巴甜(N-α-L-天冬氨酰-L-苯丙氨酸甲酯),是第一个被发现拥有甜味的肽类组分,也是最常用的无热量甜味剂[39]。目前甜味肽的研究已取得重大的突破,阿斯巴甜、阿力甜等因其甜度高、热量低已在食品领域获得了广泛的应用。Zhang等[40]已经从河豚鱼汤中分离出一种具有甜味和鲜味的八肽。Ming等[41]在研究碱性蛋白酶水解鸡肉期间滋味物质的变化时发现,肽水平在16 h达到最大值,其中对鸡肉的鲜味和甜味有重要作用的氨基酸如Glu、Asp、Ala、Gly、Ser和Thr的含量很高。已有研究证明超甜菜超阿斯巴甜类似物(pCN-C6H4 NHCO-L-Asp-L-(αMe)Phe-OMe)比超级阿斯巴甜更甜[42]。
甜味肽的呈味效果与表面丰富的侧链残基有关。早期阿斯巴甜的发现者试图用天然存在的残基替代阿斯巴甜的两个氨基酸残基,结果发现Asp不能被任何残基取代,而Phe可以被一些非疏水性残基取代[43]。研究表明大多数甜味肽在中性pH时带正电荷,其带正电荷的残基可能是某些甜味受体专一性的靶分子,当带正电荷残基减少时,甜味降低。Mazur等[39]研究发现对于原本呈现甜味的L-L型Asp-Phe-OMe所有可能的手性异构体,即D-L、L-D和D-D都呈现苦味,这一发现说明了“苦味活性位点”在结构上与甜味位点相似,但在其部分表面具有镜像特性。研究者希望通过延长甜味肽末端来增强甜味,结果发现大多数甜味二肽类似物本身具有强烈的甜味,而当向C末端延长它们的肽链时,甜味天冬二肽反而逐渐降低了甜味效价,直至五肽时完全失去甜味[44]。
3 呈味机制
寡肽所呈现的滋味是专门味觉受体细胞(TRCs)的集体感觉,这些感觉主要是由非挥发性或水溶性化合物产生[45]。味觉系统由味觉受体细胞形成,存在于舌头、上颚、咽和喉等部位,能感知不同味觉[46]。这些受体细胞呈梭形,根据功能特性和蛋白质标记物的表达不同分为“I型(暗细胞)”、“Ⅱ型(亮细胞)”、“Ⅲ型(中间细胞)”和“Ⅳ型(基细胞)”,其中Ⅱ型细胞亚群对味觉感知贡献较大[47]。味觉受体家族属于G蛋白偶联受体(GPCR)的C亚家族,包括味觉受体第一家族T1R(T1R1、T1R2和T1R3)和味觉受体第二家族T2R,其中T1R1和T1R3构成了味觉受体异源二聚体[48]。苦味、甜味和鲜味受体均属于G蛋白偶联受体,在味蕾Ⅱ型细胞中表达。且这几种滋味的GPCR都有共同的转导途径,包括异源三聚体G蛋白亚基、Gβ3、Gγ13、磷脂酶C、PLCβ2和瞬时受体电位离子通道TrpM5[49]。
到目前为止,共报道了八种鲜味候选受体,如图3[50],包括异二聚体T1R1/T1R3,代谢型谷氨酸受体Brain-mGluR1、Brain-mGluR4,代谢型谷氨酸受体味道特异性亚型Taste-mGluR1、Taste-mGluR4以及细胞外钙敏感受体(CaSR),它们都属于GPCR的C亚家族(除GPR92)。在不同的受体中,T2R1、T2R4、T2R14和T2R16都能够识别苦味肽,且T2R1表现出针对苦味二肽和三肽的特异性。甜味受体T1R2+T1R3不仅识别甜氨基酸(甘氨酸和D-色氨酸),还识别甜二肽(阿斯巴甜)和甜蛋白。林璐[51]对鲜味受体hT1R1/T1R3进行模型构建,确定了BMP与鲜味受体hT1R1/T1R3结合的活性口袋是类似于捕蝇草的结构域(VFTD),同时对比了8BMP与BMP的鲜味强度,发现8BMP与hT1R1/T1R3捕蝇结构域上关键氨基酸结合的氢键个数较多,鲜味更强。Nelson等[52]的最新研究表明T1R1+3是T1R1和T1R3蛋白偶联受体的异构体,且D-氨基酸能引发T1R1+3甜味受体的强烈活化。随着科技进步,Dang等[53]利用计算机辅助方法以及味觉受体模型,得到了T1R1和T1R3的结构模型,发现T1R3中的SER146和Glu277可能在鲜味的协同作用中发挥关键作用,当T1R3受体处于异二聚体开放构象状态时起主导作用,而T1R1受体处于闭合构象状态时发挥辅助作用。
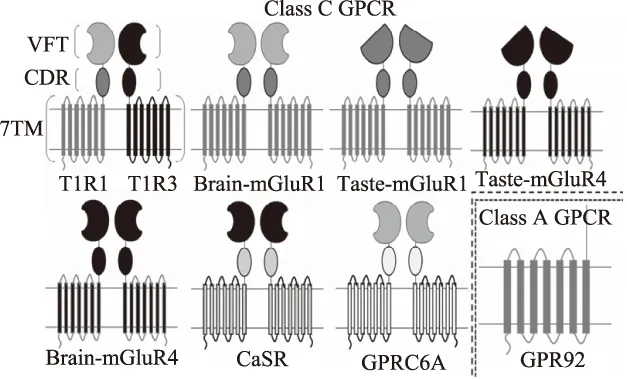
图3 几种鲜味受体的结构示意图
目前的研究认为浓厚感肽的受体是同属于G蛋白偶联受体家族的钙敏受体CaSR。细胞外的钙敏受体(CaSR)可以被γ-谷氨酰肽和芳香族氨基酸强烈激活,且当L-构型的第二个氨基酸残基上包含一至三个碳或两个碳和氧或硫原子的侧链时可促进与受体位点的结合,尤其是具有游离羧基末端且没有侧链(即甘氨酸)的残基显著增强了CaSR活性[54]。研究表明,CaSR在哺乳动物的细胞外钙稳态中起着重要作用,它可以检测到血钙水平的增加,反过来抑制甲状旁腺激素分泌,刺激降钙素分泌,并诱导尿钙排泄以将血钙降低至正常水平[55]。此外,Yue等[56]发现小肠内分泌细胞中Ca2+对中药提取物(小檗碱,黄柏碱和黄连碱)的反应会被磷脂酶C(PLC)抑制剂U-73122抑制,表明苦味受体参与改变肠内分泌细胞的生理状态,由此证明小鼠肠内分泌细胞中的小檗碱介导的胰高血糖素样肽(GLP-1)分泌途径可能与高血糖和苦味受体在小肠功能中的作用有关。以上研究证明受体不仅参与呈味作用,对人体的免疫调节和生理代谢也有一定的积极影响。
为了评估寡肽对不同滋味属性的贡献,研究表明可将蛋白质序列内的味觉活性肽的相对丰度(A)确定为定量参数(A=a/N),其中a是蛋白质序列内的感觉片段的数量,N是蛋白质中的氨基酸残基的数量。已有国外学者的试验结果表明来自猪肌纤维蛋白的肌钙蛋白C可作为基于计算机分析的苦味肽的前体(A值=0.629)[57]。
4 寡肽呈味的影响因素
肉中寡肽的呈味作用受到多种因素的影响,包括pH、加工方式、微生物以及其他因素,其他因素包括畜禽品种、肌肉类型以及宰后肌肉的成熟。此外,这些因素通过影响呈味氨基酸进而影响寡肽的滋味。
4.1 pH
寡肽的呈味作用受到氨肽酶、羧肽酶等水解酶的影响,pH则进而影响这些酶的水解作用,因此,肉品的呈味与pH存在联系。研究者发现绵羊背最长肌和股二头肌的氨基酸释放量存在明显差异,进而导致其滋味存在优劣,原因可能是二者的丙氨酰氨肽酶和氨肽酶B的活性不同[58],而这些酶具有不同的最适pH,因此,pH在一定程度会影响肉品的口感。Spanier等[59]鉴定了pH对肉类滋味的影响,发现pH为6.0、5.5和5.0是促进肽类原料产生的关键pH范围。但也有研究表明pH>5.8引起的DFD(深色,坚硬和干燥)肉会呈现类似发霉的味道。Toelstede等[60]比较浓厚感肽在pH为4.7、5.7、6.7、7.7时的呈味强度,结果发现浓厚感肽在pH为6.7时强度最强,pH为5.7时次之,而pH在4.7和7.7时的强度最低。Nishimura等[61]报道了肌肽和鹅肌肽的含量在火腿长期加工过程中显著增加,且这两种二肽在pH>4时具有重要的缓冲作用。整体上,pH通过影响氨基酸含量以及寡肽的生成进而影响肉品滋味。
4.2 加工方式
在肉制品加工过程中,滋味会因加工方式的不同而发生变化,随着加工时间的延长,大分子肽降解生成小分子肽,肉制品的滋味物质会有明显提高。Nodake等[62]用盐和磷酸盐对加拿大猪肉样品进行处理后发现,随着腌制时间的增加,猪肉中的肌肽、鹅肌肽以及酪氨酸、赖氨酸和精氨酸的含量明显提高(P<0.01)。加热温度也会加速滋味物质的分解,在Rotola-Pukkila等[63]的报道中表明,猪肉中的呈味氨基酸和IMP在正常状态下是恒定的,但若对猪肉进行蒸煮会发现随着炖煮时间的延长,二者含量显著增加。
4.3 微生物
微生物对肉类滋味的影响也十分重要。肉类发酵过程中乳酸菌,葡萄球菌和表面霉菌通过氨基酸代谢赋予肉制品味道。特别是,发酵既可以改变现有氨基酸的浓度,又可以在产品中引入新的氨基酸。董杰等[64]以木糖葡萄球菌(Staphylococcusxylosus)C18、植物乳杆菌(Lactobacillusplantarum)L26、德巴利汉逊氏酵母(Debaryomyceshansenula)Y36为发酵剂制作兔肉发酵香肠并进行营养分析,结果表明L26组甘氨酸、C18组丝氨酸的百分含量高,可改善甜味,L26组的苦味氨基酸缬氨酸含量最低,混合发酵组蛋氨酸、异亮氨酸含量最低。林璐等[51]的试验结果表明枯草芽孢杆菌因具有较强的外源蛋白表达能力而有望成为生产BMP的安全重组菌。此外,某些乳酸菌的菌株能有效去除苦味肽,这可能与乳酸菌的蛋白质降解多样性有关。
4.4 其他因素
畜禽的品种及肌肉类型会对肉制品滋味带来影响。Bousset等[65]通过高效液相色谱(HPLC)分析三种猪肌肉(背最长肌,咬肌和斜方肌)中的氨基酸和肽含量,其中咬肌中Asp,谷氨酰胺和牛磺酸含量最高,背最长肌的特征在于Gla和肌肽含量最高,而斜方肌无明显特点。Théron等[66]从干腌火腿的半膜肌和股二头肌中分别得到了43和30种肽,并分析讨论得出在火腿加工过程中不同肌肉类型可能是引起这种差异的主要原因。胡孟等[67]的实验证明,饲料中添加L-组氨酸可显著增加肉鸡肌肉中肌肽和鹅肌肽的含量,之后他们对肌肉中肌肽和鹅肌肽含量进行二次曲线方程拟合,结果显示L-组氨酸添加量为1276 mg/kg时,肌肉咪唑二肽含量最高。
5 结语
肉品中的寡肽不仅具有多种呈味作用,其在参与免疫调节、降胆固醇和抗高血压等方面也具有重要的潜在价值。但目前针对酸味肽和咸味肽的研究相对匮乏,且寡肽的呈味效果不仅只是单一的某种滋味,而是各种滋味的综合效果,因此深入研究酸味肽和咸味肽以及各种寡肽的相互作用具有重要意义。随着科学技术的发展,通过基因功能组学和微胶囊等技术筛选和开发具有特殊生理功能的呈味肽并应用到食品中将具有十分广阔的前景。
