乳酸菌合成共轭亚油酸及其应用研究进展
2020-05-08郭小婧张东辉
郭小婧,张东辉
(长江大学生命科学学院,湖北荆州434025)
共轭亚油酸(conjugated linoleic acid,CLA)是亚油酸(linoleic acid,LA)以共轭形式存在的同分异构体的统称。由于CLA 在双键位置和几何构型上存在差异,其异构体十分丰富。在众多异构体中,c9、t11-CLA 和t10、c12-CLA 是最具有生理活性的构型。CLA 是近年来被研究最多的功能性脂肪酸之一,大量研究表明,它具有抗癌、防治动脉粥样硬化、防治糖尿病、降低机体脂肪含量等多种生理功能[1]。CLA 因其重要的生理功能而被广泛应用于保健品、食品、饲料领域。天然的CLA 主要来源于反刍动物的乳肉制品、某些海产品和植物油制品。然而这类食品中,CLA 含量很低,约为0.3 mg/g~7.0 mg/g,远未达到人体体外摄取CLA 的健康推荐量3.2 g/d[2],故而从天然食物中获取CLA 将无法满足人类的需求。因此,开发高含CLA、促进人类健康的功能食品成为近几年的研究热点。
研究表明一些乳酸菌无论在体内还是体外都具有合成CLA 的能力[3],利用乳酸菌的发酵作用提高食品中CLA 含量是获得CLA 功能食品的途径之一,例如植物乳杆菌发酵豆浆后可制得富含CLA 的固态酸豆乳[4]。乳酸菌是能发酵碳水化合物产生大量乳酸的兼性厌氧细菌,广泛存在于自然界中。同时,乳酸菌是人体肠道中的正常菌群,具有帮助消化、维持肠道微生态平衡、增强免疫力等生理功能。乳酸菌良好的益生性和营养功效使其广泛应用于食品加工行业,它既可以改善食品风味,也能提高食品的营养价值和贮藏性。而且,某些乳酸菌具有一定的黏附性,可以在人体肠道内定植并进行增殖,产CLA 的益生性乳酸菌在人肠道内定植后可以持续转化LA,不断地为机体提供CLA 以促进人体的健康。由此可见,利用产CLA 的乳酸菌生产高含CLA 的功能食品具有良好的应用前景。本文从合成共轭亚油酸乳酸菌的研究进展、合成CLA乳酸菌的益生性研究和合成CLA 乳酸菌在食品、饲料中的应用研究现状3 个方面进行阐述,以期为CLA 功能食品的研究开发提供参考。
1 合成共轭亚油酸乳酸菌的研究进展
1.1 合成CLA乳酸菌的筛选及影响CLA合成的因素
乳酸菌合成CLA 的研究源于20 世纪90 年代,研究最多、最广泛的领域是菌种筛选和发酵条件的优化。由于乳酸菌培养条件较瘤胃菌、双歧杆菌易于控制,合成CLA 产量普遍较瘤胃菌高,且安全无毒可用于食品生产,所以产CLA 的乳酸菌筛选工作一直持续至今。
合成CLA 乳酸菌的筛选范围从已知菌种扩展到从泡菜、乳制品、动物肠道等中筛选,不同来源乳酸菌合成CLA 的能力见表1。
目前筛选出可产CLA 的乳酸菌有植物乳杆菌(Lactobacillus plantarum)、嗜酸乳杆菌(Lactobacillus acidophilus)、戊糖乳杆菌(Lactobacillus pentosus)、罗伊氏乳杆菌(Lactobacillus reuteri)、乳酸乳球菌(Lactococcus lactis)、明串珠菌(Leuconostoc mesenteroides)等等,其中筛选出产CLA 乳酸菌以植物乳杆菌最多,嗜酸乳杆菌次之。
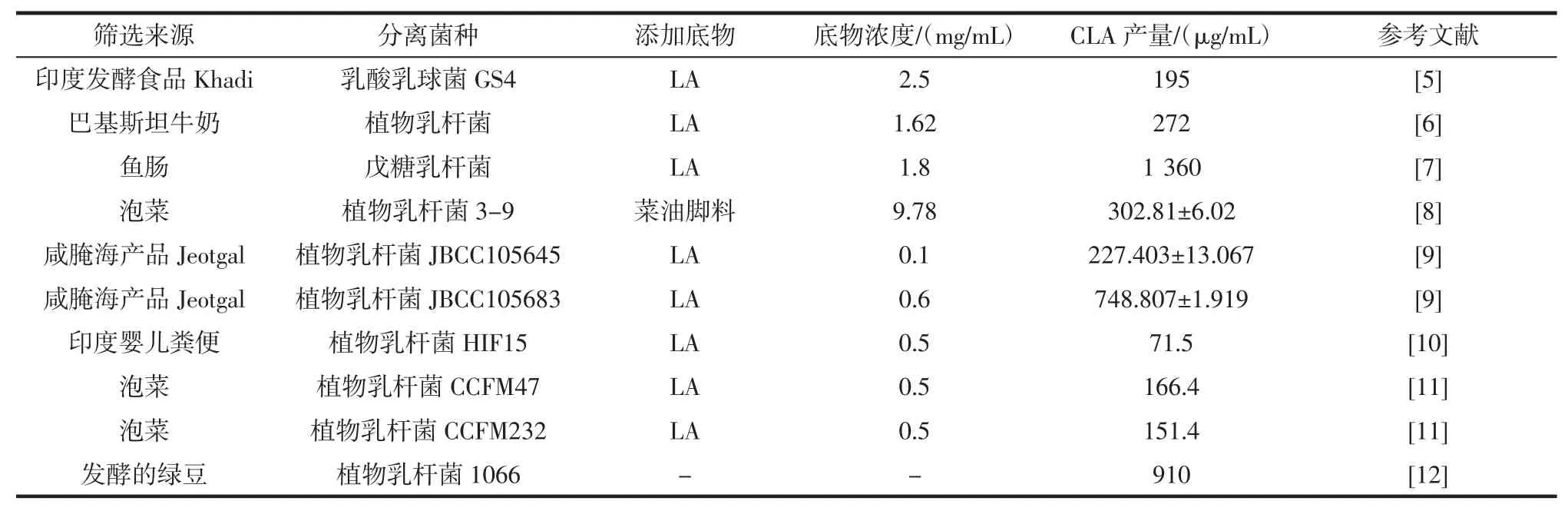
表1 不同来源乳酸菌合成CLA 的能力Table 1 The ability of CLA biosynthesis by different source of Lactobacillus
除表1 所列菌株之外,朱振元等[13]从7 株乳酸菌中筛选出可产CLA 的罗伊式乳杆菌,Vieira 等[14]从开菲尔粒中筛选出可产CLA 的乳酸乳球菌MRS47。陈丽娟等[15]从青海高寒地区的青贮玉米中分离出产CLA的植物乳杆菌ANCLA2。由表1 可知,不同来源筛选出的乳酸菌合成CLA 的能力相差很大,这与菌株密切相关,同时也受添加底物类型及浓度、发酵方式的影响。合成CLA 的乳酸菌可利用的底物有LA、红花油、葵花籽油、蓖麻油、元宝枫油等。利用植物油替代纯品LA作为底物既可以降低成本,也能减弱高浓度LA 对菌体生长的抑制作用。植物乳杆菌CGMCC8198 利用0.5 mg/mL 元宝枫油为底物发酵 24 h 后,c9、t11-CLA和 t10、c12-CLA 产量分别为 2.11、3.49 mg/mL,是未添加元宝枫油产量的9.16 和2.25 倍,同时一定浓度的元宝枫油(0.5 mg/mL~1 mg/mL)可以上调植物乳杆菌CGMCC8198 亚油酸异构酶(linoleate isomerase)基因的表达[16]。
乳酸菌合成CLA 的发酵方式主要有细胞发酵法、酶法、静息细胞法及固定化细胞法4 种,较传统发酵法而言,静息细胞法生成产物较单一,杂质少,能添加高浓度底物从而获得CLA 的高产量,固定化细胞可重复多批次使用,且能长时间保持细胞的稳定性,因此这两种方法是近年来用于提高CLA 产量使用最多的方法。Khaskheli 等[17]利用植物乳杆菌静息细胞转化米根酶酶解的蓖麻油生成CLA,蓖麻油浓度为8 mg/mL,米根酶浓度0.2 mg/mL,静息细胞浓度12%,pH 6.5,37 ℃反应 20 h 后 CLA 的生成量为 406 μg/mL,其中 c9,t11-CLA 占 56.55%,t10,c12-CLA 占 43.45%。固定化植物乳杆菌3-9 可转化菜油脚料合成CLA:固定化材料为直径1 mm~3 mm 的瓷颗粒,固定时间为1 h,添加菜油脚料浓度9.78 mg/mL,细胞浓度0.6 g/mL,35 ℃连续转化 16 批次,CLA 最高产量为(302.81±6.02)μg/mL[8]。
此外,乳酸菌合成CLA 的产量还受培养基种类、培养时间、接种量、培养温度等多种因素的影响。植物乳杆菌1066 在MRS 培养基中培养18 h 和24 h 后,其CLA 产量分别为 327.67 μg/mL 和 259.33 μg/mL[18],使用奶牛奶培养基、水牛奶培养基和羊奶培养基培养植物乳杆菌 1066 18 h 时,CLA 合成量分别为 510、910、320 μg/mL,培养 18 h 后,CLA 合成量分别为 380、620、190 μg/mL,随着培养时间的延长,CLA 产量下降[12]。
乳酸菌在合成CLA 的过程中生长会受到高浓度底物的抑制,从而限制了高浓度底物的添加,导致最终CLA 的产量较低。对于合成CLA 乳酸菌的筛选后续应着重筛选能耐受高浓度底物的乳酸菌,同时可优化其发酵方式及培养条件提高乳酸菌合成CLA 的能力。此外,还可以通过构建基因工程菌株或诱变技术来提高CLA 产量。
1.2 乳酸菌利用LA合成CLA的机理及相关酶学性质分析
乳酸菌利用LA 合成CLA 的机理目前主要认为是细胞通过对LA 的氢化作用以消除LA 对细胞的危害,例如植物乳杆菌ATCC8014 等。同时某些乳酸菌具有亚油酸异构酶系,可以将LA 主要异构化为c9,t11-CLA 和 t9,t11-CLA,例如植物乳杆菌 AKU1009a 和 植物乳杆菌ZS2058 等。Ortega-Anaya 等[19-20]研究了植物乳杆菌 ATCC8014 利用 LA 合成 c9,t11-CLA 的机制,LA 首先在亚油酸水合酶(linoleate hydratase,MCRA)的作用下生成10-羟基-顺12-十八碳烯酸(10-Hydroxy-cis-12-octadecenoic acid,10-HOE),10-HOE 在α-烯醇酶(α-enolase)的作用下直接脱水异构化为c9,t11-CLA,植物乳杆菌利用LA 异构化为CLA 途径见图1。
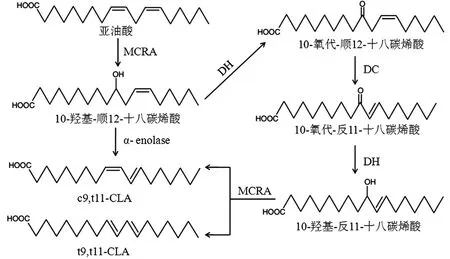
图1 植物乳杆菌利用LA 异构化为CLA 途径Fig.1 Proposed pathway of linoleic acid isomerization to CLA in Lactobacillus plantarum
植物乳杆菌 ATCC8014 的 MCRA 分子量为64.74 kDa,是包含有3 个结构域的外在膜蛋白,酶促反应最适 pH 值为 6.5~7.0,MgCl2(50 mmol/L)、(NH4)2SO4(500 mmol/L)、十二烷基-β-D-麦芽糖苷(2.5%)、聚乙烯乙二醇400(25%)和乙醇(15%)的任一添加均可以增强MCRA 的稳定性。植物乳杆菌ATCC8014 的α-烯醇酶分子量为48.03 kDa,包含有2 个结构域,生成c9,t11-CLA 的最适 pH 值为 7.0,不能直接利用 LA 为底物,任一添加MgCl2、甘油、十二烷基-β-D-麦芽糖苷和乙腈均可以增加α-烯醇酶的稳定性,并能提高酶活使其合成c9,t11-CLA 的产量提高 5 倍~6 倍。Yang等[21-22]发现在植物乳杆菌ZS2058 中,催化生成CLA 涉及多步反应,包括3 种酶:LA 首先经水合作用生成10-HOE,然后经脱水、双键移位、水合、脱水等多步反应生成CLA,如图1 所示。克隆植物乳杆菌ZS2058 的MCRA、短链氢化酶/氧化还原酶(short-chain hydrogenase/oxidoreductase,DH)、乙酰乙酸脱羧酶(acetoacetate decarboxylase,DC)编码基因后转入大肠杆菌中异源表达,重组大肠杆菌中检测到了c9,t11-CLA 和3种中间体,证明了CLA 的多步合成路径,经检测发现MCRA、DH、DC 的分子量分别为 64.76、32.10、30.71 kDa。将植物乳杆菌 ZS2058 的 MCRA、DH、DC 编码基因分别转入耶氏解脂酵母中,酶活测定结果显示有产物10-HOE 和10-氧代-反11-十八碳烯酸的生成,再次证明了植物乳杆菌ZS2058 的CLA 合成路径[23]。分析7 株CLA 合成能力差异的乳酸菌的亚油酸异构酶系各组成基因转录水平,结果显示亚油酸异构酶系中的DH、DC 的转录水平差异是造成乳酸菌合成CLA 能力不同的内在原因[24]。赵彤彤等[25]扩增植物乳杆菌P-8的羟基脂肪酸脱氢酶基因转入大肠杆菌,纯化的羟基脂肪酸脱氢酶可以转化10-HOE 为10-氧代-顺12-十八碳烯酸,植物乳杆菌P-8 的羟基脂肪酸脱氢酶分子量为32.09 kDa,等电点为5.15,属于细胞质蛋白。其实验室同时也证明了植物乳杆菌P-8 的脂肪酸水合酶能将LA 水合为10-HOE,这为揭示植物乳杆菌P-8合成CLA 路径奠定了良好的实验基础。
虽然部分乳酸菌合成CLA 的途径已经明确,但因种属区别乳酸菌合成CLA 的过程远不止图1 所列,未来仍需深入研究,力争全面揭示乳酸菌合成CLA 的途径。
1.3 合成CLA乳酸菌的诱变及基因工程菌株
国内学者为了提高乳酸菌生产CLA 的能力,广泛开展对产CLA 乳酸菌的诱变处理工作。通过紫外线、硫酸二乙酯、亚硝基胍、N+离子注入等多种诱变方式对产CLA 乳酸菌进行诱变处理后,突变株合成CLA 的能力有所增强。而且与单一诱变方法相比,采用多种方法对乳酸菌进行多次诱变处理更能提高菌株产CLA 能力。德氏乳杆菌保加利亚亚种(Lactobacillus delbrueckii subsp.bulgaricus)通过紫外和硫酸二乙酯混合诱变后,其 CLA 产量由 55 μg/mL 提高至 292μg/mL[26]。乳杆菌SC-05 经低温等离子体诱变后,CLA 产量由65.16 μg/mL 提高至 196.27 μg/mL[27]。张珂[28]使用常压室温等离子体诱变技术对嗜酸乳杆菌进行诱变处理,筛选出亚油酸异构酶高产突变株,突变株亚油酸异构酶酶活最高为1 214.1 U/mL,是原始菌株的2.84 倍。
除通过诱变外,还可以通过构建基因工程菌株来实现乳酸菌CLA 产量的升高或异源表达乳酸菌亚油酸异构酶来提高酶活。将痤疮丙酸杆菌(Propionibacterium acnes)的亚油酸异构酶PAI 编码基因导入食品级乳酸乳球菌NZ3900 中,得到高产t10,c12-CLA 的重组乳酸乳球菌,重组菌菌体中t10,c12-CLA 含量占总脂肪酸的23.91%,LA 转化率为37.6%,高于乳酸乳球菌自身LA 转化率[29]。干酪乳杆菌(Lactobacillus casei)的亚油酸异构酶LAI 编码基因转入大肠杆菌中,得到能将LA 特异性转化为c9,t11-CLA 的重组菌,LAI 的酶活最大为 206 U/mL[30]。
2 合成CLA乳酸菌的益生性研究
用于合成CLA 功能食品的乳酸菌需具有良好的益生性,既要能耐受胃液(低pH 值),胆汁(高胆盐)和肠液(胰蛋白酶)的消化作用,又要具有较好的黏附性,抗菌性,能抑制病原菌生长。此外,在食品加工和储藏过程中,益生菌需保持较高的存活率。具有益生性的产CLA 乳酸菌无论用于食品加工还是口服定植于人体肠道,都会给人体提供CLA,为人体健康带来福音。而不同的乳酸菌其益生性存在较大差异,因此,研究合成CLA 乳酸菌的益生性将推动产CLA 乳酸菌的应用。Li 等[31]从人类粪便中分离出产 c9,t11-CLA 的嗜酸乳杆菌 F0221 能耐酸(pH 3.0)、耐受高胆盐(0.15 %~0.5 %)、对肠道上皮细胞具有较好的黏附力[(117±26)CFU/100 个细胞],嗜酸乳杆菌 F0221 在添加 0.5 g/L LA 为底物,pH 6.5 的厌氧条件下生成最多的c9,t11-CLA 产量为(102.1±12.6)μg/mL。Sosa 等[32]从 13 株乳酸菌中筛出4 株产CLA 且具有益生特性的乳酸菌,即发酵乳杆菌(Lactobacillus fermentum)J20,发酵乳杆菌J23,植物乳杆菌J25,戊糖乳杆菌J26,其合成CLA 产量分别为 (50.9±0.26)、(18.98±1.03)、(23.53±1.94)、(13.44±0.78)μg/mL。同时这4 株乳酸菌在模拟的胃肠道环境中均具有活性且能产生CLA,并可以黏附在小鼠的肠黏膜上。从内蒙古传统发酵奶油中筛得潜在黏附性较高的产CLA 乳酸菌HH7、AS8,其CLA 产量分别为:(7.37±0.41)、(13.22±0.52)μg/mL,HH7、AS8 能耐酸(pH 2.5)、耐胆盐(0.3%)、耐胃蛋白酶(5 mg/mL)、能抑制大肠杆菌和金黄色葡萄球菌的生长,AS8 还能耐受胰蛋白酶(10 mg/mL)。分别用HH7、AS8 菌液灌胃小鼠,其中用HH7 灌胃的小鼠较对照组体重增加,肠道中乳杆菌和双歧杆菌的数量显著增加,有害菌显著减少,表明HH7 能定植肠道并调节肠道菌群。用AS8 灌胃的小鼠体重增长缓慢,肠道中活菌数量较对照组无明显差异[33]。从婴儿粪便中分离出的可产CLA的发酵乳杆菌DDHI27 益生性检测结果为:可耐酸(pH 1.5)、耐高胆盐(2%)、具有抗氧化和抑菌活性、对Caco-2 细胞模型具有较高的黏附力[(25.10±2.1)CFU/细胞][34]。发酵乳杆菌DDHI27 在脱脂奶粉培养基添加LA 0.5 mg/mL,pH 6.0,37 ℃培养 48 h 后合成 c9,t11-CLA:31.97 μg/mL、t10,c12-CLA:5.94 μg/mL 和 t9,t11-CLA:9.99 μg/mL[35]。Palachum 等[36]从印度传统发酵药草木蝴蝶中分离出植物乳杆菌WU-P19 并检测了其益生特性:可产生胆盐水解酶水解胆盐(甘氨胆酸、甘氨脱氧胆酸)、产生β-半乳糖苷酶、具有抵抗抗生素(庆大霉素、卡那霉素、万古霉素、妥布霉素等)特性、能抑制蜡样芽胞杆菌、肠炎沙门氏菌和金黄色葡萄球菌的生长、可在模拟的胃肠道环境中传代。植物乳杆菌WU-P19 合成CLA 最大含量为21 mg/g 菌体,其中CLA 异构体主要为cis9,trans11-CLA 和trans10,cis12-CLA[37]。从奶酪中分离出产CLA 的植物乳杆菌L3C1E8,其 cis9,trans11-CLA 的合成量为 (15.36±0.15)μg/mL,其益生性检测结果显示:耐胆盐(0.3%)和胰液(0.1%)、耐酸(pH 2.5)、对 Caco-2 和 HT-29 细胞模型表现出良好的黏附性、抑制大肠杆菌生长[38]。
上述这些产CLA 乳酸菌均具有良好的益生性,后续通过临床安全性评价后,可以进一步利用这些乳酸菌发酵食品以提高食品中CLA 含量,形成含CLA 的功能食品,或者让这些乳酸菌定植人体肠道,使其长期有效地转化LA 为CLA,为人体提供源源不断的CLA 以促进人体健康。合成CLA 乳酸菌的益生性研究为产CLA 乳酸菌的应用奠定了基础,推动了CLA 功能食品的开发。
3 合成CLA乳酸菌在食品、饲料中的应用研究现状
功能食品作为保健食品受到广大消费者的青睐,CLA 功能食品具有减肥、抗癌、增强免疫力等多种功效,已引起科学界的广泛关注。目前获取CLA 功能食品的主要方式是在食品或动物饲料中直接添加CLA或LA 或富含LA 的植物油,但纯品的CLA 或LA 价格昂贵,增加了应用成本,富含LA 的植物油的大量摄食会增加脂肪含量,不利于生产实践的推广。利用产CLA 的益生性乳酸菌制备CLA 功能食品或饲料则明显降低生产成本,更有利于推广,并且乳酸菌发酵产生的CLA 与天然食物中CLA 异构体组成最相近,能较好的满足消费者的心理需求。同时,益生性乳酸菌可以改善人体和动物的肠道环境,和CLA 一起共同促进人体和动物的健康。因此,利用产CLA 益生性乳酸菌生产CLA 功能食品或饲料具有深度开发价值。
3.1 合成CLA乳酸菌在食品中的应用
合成CLA 乳酸菌在食品中的应用主要从两个方面展开,一方面利用产CLA 乳酸菌发酵食品从而提高食品中 CLA 含量,即 CLA 功能食品。Jin 等[39]用植物乳杆菌P1201 发酵大豆生产的豆浆中含有CLA(1.3±0.1) mg/g,其中 c9,t11-CLA(1.2± 0.1)mg/g,t10,c12-CLA 0.1 mg/g,而未用该菌株发酵的豆浆中未检测到CLA。用该含CLA 豆奶喂养高脂膳食小鼠后,小鼠体重减少10.7%,脂肪肝减少32.8%[40]。用乳酸乳球菌MRS47 发酵牛奶,40 ℃发酵 8 h 后,牛奶中 CLA 含量为850 μg/mL,提取发酵牛奶的脂肪酸检测CLA 含量为:c9,t11-CLA 66.5 mg/g 脂肪酸,t10,c12-CLA 18.7 mg/g 脂肪酸,c10,c12-CLA 13.4 mg/g 脂肪酸,其中 c9,t11-CLA 含量是对照组的 9.3 倍[14]。Ozer 等[41]用植物乳杆菌AB20-961 发酵生产的土耳其香肠中含有CLA,在制作过程中CLA 含量为4.9 mg/g 脂肪,发酵24 h 后其CLA 含量为5.4 mg/g 脂肪,在储藏期中CLA 含量为4.4 mg/g 脂肪,比对照组CLA 含量增加了0.1 mg/g~0.6 mg/g 脂肪。使用植物乳杆菌R6、嗜热链球菌、德氏乳杆菌保加利亚亚种(1 ∶1 ∶1)发酵脱脂奶,添加 LA 0.11%,脱脂乳浓度 11.5%,37 ℃发酵 40.77 h 后,酸奶中 CLA 含量达到 1 030 μg/mL[42]。
另一方面,可将产CLA 益生性乳酸菌制成菌粉后,作为食品或药品添加成分制成功能食品或保健药物,人体通过口服该制剂后,乳酸菌利用人体内的LA合成CLA。科学研究表明,健康人体每天平均约排泄20 mg LA,这表明人体内有足够的LA 底物供产CLA乳酸菌利用[43]。Lee 等[44]首次报道了可以在人体内产生CLA 的乳酸菌,即植物乳杆菌PL60,志愿者服用植物乳杆菌PL60 冻干粉1 周后,其排泄物中可以检测到植物乳杆菌PL60,连续服用21 d 后,其血清中c9,t11-CLA 和t10,c12-CLA 浓度显著升高,这说明植物乳杆菌PL60 可以在人体肠道内存活并且在人体内合成CLA。这为产CLA 益生性乳酸菌的应用奠定了基础。中国专利“具有减脂瘦身功能的益生菌组合物及其制备方法与应用”中,使用可产CLA 的双歧杆菌、植物乳杆菌和嗜酸乳杆菌益生菌干粉混合金银花、青蒿、黄芪、葛根和黄苓提取物粉制得益生菌组合物,该组合物中益生菌产生的CLA 可降低脂肪,从而达到减脂瘦身功能。用该益生菌组合物喂养肥胖大鼠后可显著降低其体重[45]。从健康幼儿粪便中分离出的罗伊乳杆菌HI120,主要将LA 转化为c9,t11-CLA,其转化效率分别比罗伊乳杆菌标准菌DSM 20016 和长双歧杆菌的NCC2705 高6.1 和9.1 倍。将该菌制备成肠溶性活菌菌粉喂养肥胖糖尿病小鼠,结果显示罗伊乳杆菌HI120 可显著降低小鼠空腹血糖、体重、食量及血液中胆固醇和甘油三酯含量、显著减轻脂肪肝,同时罗伊乳杆菌HI120 还可以显著减少小鼠结肠癌肿瘤体的大小和数量[46]。从健康女性的阴道细菌菌群中分离出可产CLA 的副干酪乳杆菌LMG S-26420,主要将LA转化为 c9,t11-CLA 和t9,t11-CLA,其 LA 转化率>30%,冻干形式的副干酪乳杆菌LMG S-26420 菌粉可作为营养、食品或药物添加剂,用于预防和治疗与CLA 缺乏相关病理[47]。
3.2 合成CLA乳酸菌在饲料中的应用
使用产CLA 乳酸菌或其发酵物制备的饲料喂养动物后,不仅可以改善动物的生长和健康状况,还可以提高动物乳、肉或蛋制品中CLA 含量,从而得到富含CLA的食品。Herzallah[48]以乳酸菌饲喂肉鸡和蛋鸡提高了鸡肉和鸡蛋中CLA 含量,其中用1 mL(106CFU/mL)从山羊瘤胃中分离出的罗伊氏乳杆菌连续喂养肉鸡5 周后,鸡肉中 CLA 含量为(1.81±0.113)mg/g 脂肪,是对照组的3.62 倍。用1 mL(106CFU/mL)从骆驼瘤胃中分离出的罗伊氏乳杆菌连续喂养蛋鸡5 周后,鸡蛋中CLA含量为(1.25±0.033)mg/g 脂肪,是对照组的 6.25 倍。中国专利CN 201611019128.8 公开了“一种微生物发酵生产含共轭亚油酸发酵物的制备方法”,该发明使用乳酸杆菌和啤酒酵母发酵米糠、大豆粉和大米粉,经滚筒发酵烘干后制得CLA 含量≥6.0%的发酵物[49]。另一专利“一种饲养雪花牛肉的发酵饲料的制备方法”中,使用植物乳杆菌、枯草芽孢杆菌和酿酒酵母混合发酵富含亚油酸的玉米胚芽粉、小麦粉、大麦芽粉和葛根全粉,制备出一种用于饲养雪花牛肉的富含CLA 和葛根黄酮类物质的发酵饲料,用该饲料等量取代原有饲料,可明显提高牛肉品质[50]。将青贮玉米中分离出的产CLA 植物乳杆菌ANCLA2 回接到全株玉米中,玉米青贮中 CLA 含量达到(13.62±0.94)μg/g,是对照组(未接菌剂)的8.9 倍[15]。该玉米青贮可以作为含CLA 饲料喂养牛羊鱼等,含CLA 的玉米青贮饲料喂养草鱼后可提高草鱼的生长速度及蛋白率。
4 展望
随着人们保健意识的逐渐增强,CLA 因具有重要的生理功能其市场需求量将会持续增加,天然来源的CLA 远远满足不了人们的需求。目前CLA 的生产工艺主要是化学法,化学法合成产物复杂,提纯困难,生产留下的废弃液还会污染环境,不适应时代的发展。微生物法生产CLA 还处于研究阶段,未能实现规模化生产。增加食品中CLA 含量制备CLA 功能食品已经成为众多科学家的研究重点,一方面制备CLA 功能食品操作简单,原材料天然无毒,终产物无需纯化,另一方面产品可以直接食用,既能享受食品的风味,也能获取CLA,并且无污染废弃物。乳酸菌是人体肠道正常菌群,利用乳酸菌生产的CLA 功能食品更能被人们所接受。但是目前产CLA 益生性乳酸菌生产出的食品中CLA 含量并不高。多年来筛选到的产CLA 乳酸菌很多,而用于生产CLA 功能食品的乳酸菌并不多,在后续研究中,应加大可产CLA 乳酸菌的益生性及安全性研究,着重研究如何提高食品中CLA 含量获得高含CLA 的功能食品,并且逐步开展产CLA 益生性乳酸菌在人体肠道定植的研究,为产CLA 益生性乳酸菌直接服务于人类提供理论基础。
