广州市2018年输入性登革病毒4型全基因组序列特征分析
2020-05-05苏文哲蒋力云曹毅敏李意兰陈宗遒朱娉婷景钦隆胡玉山张周斌
苏文哲,曾 庆,蒋力云,曹毅敏,李意兰,陈宗遒,朱娉婷,景钦隆,狄 飚,胡玉山,张周斌
登革病毒(Dengue virus, DENV)是一种经伊蚊传播的虫媒病毒,感染人体后可导致登革热,目前已有超过100个国家和地区报道过登革热病例[1]。DENV目前已发现有4个血清型(DENV-1~4),每个血清型之间存在一定的交叉反应;异型DENV的二次感染常可引发宿主的严重免疫反应,即所谓的抗体依赖性感染增强现象(antibody dependent enhancement,ADE),进展为重症病例(包括登革出血热和登革休克综合征)[2]。DENV基因组为单股负链RNA,全长约11 000 bps,包含1个开放读码框架,共编码3个结构蛋白(capsid/membrane/envelop, C/M/E)和7个非结构蛋白(NS1/NS2A/NS2B/NS3/NS4A/NS4B/NS5)[2]。全球每年约有3.9亿人感染DENV,其中9 600万感染者具有明显的临床症状,绝大多数的感染者均可自愈,但该疾病所带来的社会负担仍然不可忽视,登革热已经成为全球性的公共卫生问题[3]。目前已有针对4种血清型DENV的疫苗Dengvaxia问世,并已在国外获批投入使用,但由于存在安全问题,未感染过DENV的人群在接种疫苗后感染可能加重临床症状,因此尚未在我国投入使用[4]。
我国建国后首次有记录的登革热暴发疫情始于1978年,波及的省份包括广东、广西、云南、福建和浙江[5]。登革热的主要传播媒介白纹伊蚊和埃及伊蚊均在广东省本地被发现过,在过去30年间,常有输入性的登革热病例引起的本地暴发疫情的报道;最近一次大规模的暴发疫情发生在2014年,由DENV-1型病毒引起,全省报告病例共计45 236例,其中广州市报告病例37 340例,为近10年来最高[6]。东南亚各国气候炎热,卫生条件较差,登革热疫情常年不断,毗邻广东省,是广东省登革热的主要输入地[3]。近年来随着对外贸易和外出旅游频率的不断提高,广州市每年均有输入性登革热病例的报道,但近5年尚未有DENV-4型本地病例的报道。因此,本研究拟通过对广州市2018年报告的输入性DENV-4型登革热病例血清标本进行分离培养,并进一步分析DENV-4其生物学来源,从而为本地登革热的防控提供科学依据并累积基线数据。
1 材料与方法
1.1病例及标本来源 本研究所涉及的输入性登革热病例标本均来自2018年广州市各医院和区疾控中心送检/复核的输入性登革热排查病例血清标本,每份标本均有独立的编号,并有完整的标本信息;标本的使用征得患者知情同意并通过广州市疾病预防控制中心伦理委员会批准;同时通过国家疾控中心传染病监测信息报告系统和广州市疾控中心实验室信息系统收集病例信息。
1.2标本初筛和病毒分离 核酸检测和病毒分离过程参考《登革热诊断标准》(WS 216-2018),使用QIAamp Viral RNA Mini Kit(QIAGEN)试剂盒对血清标本进行核酸提取,进而采用DENV通用型和DENV-1~4型核酸检测试剂盒(江苏硕世生物有限公司)进行实时荧光定量PCR检测。对DENV-4核酸阳性标本在C6/36细胞进行连续传代培养,传代3次无病变者方判定为病毒分离阴性。
1.3病毒基因组测定 使用QIAamp Viral RNA Mini Kit核酸提取试剂盒(QIAGEN)对分离株进行核酸提取,提取完毕马上进行基因扩增。全序列测定所用到的引物详见表1[7]。使用SuperScriptTMIII One-Step RT-PCR System with PlatinumTMTaq DNA Polymerase试剂盒(Invitrogen)进行扩增,反应体系参考说明书,反应体系总体积为50 μL。反应条件为:55 ℃ 30 min, 94 ℃ 2 min;94 ℃ 30 s, 58 ℃ 30 s, 68 ℃ 2 min, 40个循环;68 ℃ 10 min, 4 ℃ ∞。使用ABI 9700基因扩增仪进行扩增,在完成基因扩增后,用QIAxcel全自动分析系统对扩增产物进行分析,得到目的片段后进行测序。序列的测定委托天一辉远基因科技公司完成。
1.5序列比对分析、进化树重建和重组分析 参考序列均来自NCBI(https://www.ncbi.nlm.nih.gov),一共下载了84条DENV-4型病毒的基因组序列,所选取的参考序列均是DENV-4型病毒在不同年代和不同地区的代表株,均有明确的分离时间和地点,并涵盖了DENV-4型病毒的4个基因型。用Smart Model Selection(SMS,http://www.atgc-montpellier.fr/sms/)在线工具和贝叶斯信息准则(Bayesian Information Criterion,BIC)寻找最优核酸替换模型和参数,用PhyML 3.1在线工具(http://www.atgc-montpellier.fr/phyml/)重建进化树,进化树的支长计算采用贝叶斯检验(aBayes)的方法[8]。核苷酸和氨基酸序列的差异分析通过MEGA 6.0软件(Molecular Evolutionary Genetics Analysis)完成。
表1 DENV-4病毒基因组全序列测定引物
Tab.1 Primers for DENV-4 full-length sequencing

引物名称引物序列(5′-3′)方向退火温度/℃D4-F1TTAGTCTGTGTGGACCGACAAGGACAG正向68D4-R806AATCCTGGGTTTCTGAGTATCCAACTCTC反向66D4-F683AGTGGGGAACGGAGACGAGAGAAG正向68D4-R1406GGGTTATCGTAGCTGTCACTCCGTG反向68D4-F1088GAAGTGGCTCTGTTAAGAACCTATTGCATCGAAGC正向67D4-R1928CATCTCTTATCTCTATGGGAACTTTGCATGG反向66D4-F1642AATGGTGACGTTTAAGGTCCCTCATGC正向66D4-R2370AGAAACAAAGTAATCCCTCCAACAGCTATGC反向66D4-F2195GATTTTGGCTCTGTTGGTGGACTGCTC正向68D4-R3084ATGTGTGGGTCTTGGGCCATAGACATG反向68D4-F2696GTGAAAGGGGTGTTATCCAAAGGCAAGAG正向67D4-R3571GCACAAAGGGTGGTCACCACAACCAATATC反向69D4-F3264GAACAACAGTCACCATTCAAGAGGATTGTGACC正向68D4-R4177CATTCTTTAGAAGGGCGCTTCCCAAG反向66D4-F3917GCCCTTTCCTTGACTTTCATAAAATCAACAATGC正向66D4-R4785GCTAGAACTTGAACGTCTTCTTCTTTATCCC反向66D4-F4464TGGCAATTCCAGTCACAATGACCCTATGG正向67D4-R5314GAACTCTGGTTGATGATAAAAGTCTTGTTGTG反向65D4-F5028GAATTGGCGAACCAGATTATGAAGTGGATG正向66D4-R5852TTGCTGGAGTCACTGGAATAGGACC反向66D4-F5498CAGAGCAATAGCCCAATAGAAGATATCGAGAG正向67D4-R6554GAAGATGCCTGCTGTCATAGCTCCTAG反向68D4-F6400AATTGCCAGTTTGCCAACGTACCTTTCCTC正向67D4-R7458ATGTTAGCTGTGGACACGGCTATAGTCG反向68D4-F7079CTTGGTGTGCCGCTATTAGCAATGGGATGC正向70D4-R7877CATAGGAATCGGTTCTTCATGTCCTGGACC反向69D4-F7700GGATCTAAAATCAAGTATGCAGTGTCCAGAG正向66D4-R8708ACAGTCTGGGATTTTTCTTCTTCCCGAGGAG反向69D4-F8486TCCTTCGACAGGCTCGGCATCCTC正向70D4-R9206AGGGGCCATCTGTTCTGTGATCAGCTCTTC反向70D4-F9105GATACATCCTGGAGGACATAGATAAGAAAGATG正向66D4-R9863AGGCTAAGCGCAGGTCCCTTCTATG反向68D4-F9482GAAAGAGTTGAGAAATGGCTGAAAGAGTGTG正向66D4-R10353GCCTCCCTGGGATTTTTACGCCTCCCG反向68D4-F10247GAAGGAGTTCTGTAATTACCAACAACAAACACC正向66D4-R10716CACGAGGAAGCTGTACTCCTGGTG反向68
2 结 果
2.1毒株分离情况 对2018年广州市输入性登革热病例的标本进行检测、鉴定,最后分离培养得到DENV-4型毒株5株,毒株编号和信息如表2所示:
2.2核苷酸与氨基酸同源性比较 将本研究分离到的5株DENV-4型毒株与6株参考株的全基因组序列进行核苷酸和氨基酸的同源性分析。结果显示,4株由泰国和柬埔寨输入的分离株(18XN11706/18XN13312/18XN16107/18XN17096)的核苷酸(氨基酸)同源性为99.2%~99.8%(99.7%~99.9%),与DENV-4型病毒Sylvatic基因型 (EF457906.1)的核苷酸(氨基酸)同源性为85.9%~86.0%(95.4%~95.5%);菲律宾输入的病例分离株(18XN20946)与中南半岛来源的4株分离株核苷酸(氨基酸)同源性为90.8%(96.7%~96.9%),与菲律宾2014年分离株(KU523871.1)的核苷酸(氨基酸)同源性为98.6%(99.1%)与DENV-4型病毒Sylvatic基因型(EF457906.1)的核苷酸(氨基酸)同源性为86.1%(95.1%)。具体结果详见表3。
表2 广州市2018年输入型DENV-4毒株信息
Tab.2 Information of imported DENV-4 strains in Guangzhou, 2018
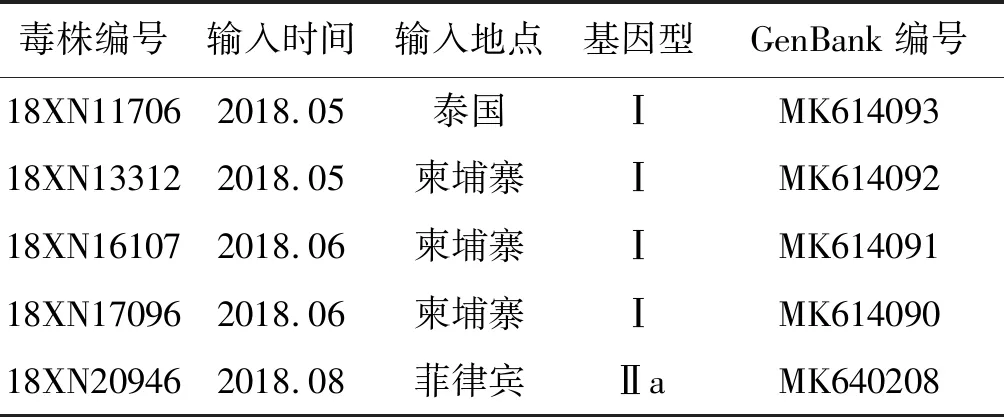
毒株编号输入时间输入地点基因型GenBank编号18XN117062018.05泰国ⅠMK61409318XN133122018.05柬埔寨ⅠMK61409218XN161072018.06柬埔寨ⅠMK61409118XN170962018.06柬埔寨ⅠMK61409018XN209462018.08菲律宾ⅡaMK640208
表3 广州市2018年输入型DENV-4毒株与参考株全序列核苷酸和氨基酸同源性比较
Tab.3 Homology comparisons of full-length genomes among reference DENV-4 strains and imported strains in Guangzhou, 2018

序列编号18XN1170618XN1331218XN1610718XN1709618XN20946AY618988.1AY762085.1EF457906.1JN638572.1KP792537.2KU523871.118XN11706—99.499.299.490.890.091.385.995.898.590.8 18XN1331299.8—99.699.890.890.091.486.095.898.690.8 18XN1610799.799.8—99.790.990.091.386.095.898.590.8 18XN1709699.899.999.9—90.890.091.485.995.898.690.8 18XN2094696.996.896.796.8—90.292.686.191.491.298.6 AY618988.197.197.096.997.097.0—90.786.090.190.290.3 AY762085.197.597.597.397.497.897.3—86.491.791.792.8 EF457906.195.695.595.495.595.195.595.7—86.086.186.0 JN638572.198.498.498.398.396.896.897.295.2—96.291.5 KP792537.299.699.699.499.596.997.297.595.598.5—91.1 KU523871.196.996.896.796.899.197.098.095.096.897.0—
注:表格右上部分为核苷酸同源性比较结果,左下部分为氨基酸同源性比较结果。
2.3DENV-4型病毒基因组全序列进化树重建结果 通过SMS模型选择工具计算得出最优的核酸替换模型为GTR(G+I)模型(General Time Reversible Model),参数选择为G=2.069,I=0.535。进化树重建结果如图1所示。从图1中可见,4株由泰国和柬埔寨输入的分离株(18XN11706/ 18XN13312/ 18XN16107/ 18XN17096)在进化上较为接近,属于DENV-4基因Ⅰ型,在GenBank上同源性最高的序列是KP792537.2(新加坡2011年分离株);菲律宾输入的病例分离株(18XN20946)与其他4株亲缘性较远,属于DENV-4基因Ⅱa型,在GenBank上同源性最高的序列是KU523871.1(菲律宾2014年分离株)和KC333651.1(广州市2012年输入病例分离株)。进化树的重建结果详见图1。
2.4氨基酸位点差异分析 经过比对,5株分离株和6株参考株的E蛋白区域有9个位点出现差异,NS1蛋白区域有14个位点出现差异,NS5蛋白区域有25个位点出现差异;4株由泰国和柬埔寨输入的分离株(18XN11706/18XN13312/18XN16107/18XN17096)和菲律宾输入病例的分离株(18XN20946)序列差异较大。具体结果详见表4、表5和表6。

注:红色序列为广州市2018年分离株,黑框内标注为输入地和输入时间。图1 广州市2018年DENV-4分离株基因组全序列进化树重建结果Fig.1 Phylogenetic tree based on full-length genomes of 5 imported DENV-4 strains isolated in Guangzhou, 2018
表4 广州市2018年输入型DENV-4毒株与参考株E蛋白氨基酸差异位点比较
Tab.4 Amino acid substitutions in E gene region among reference DENV-4 strains and 5 imported strains in Guangzhou, 2018
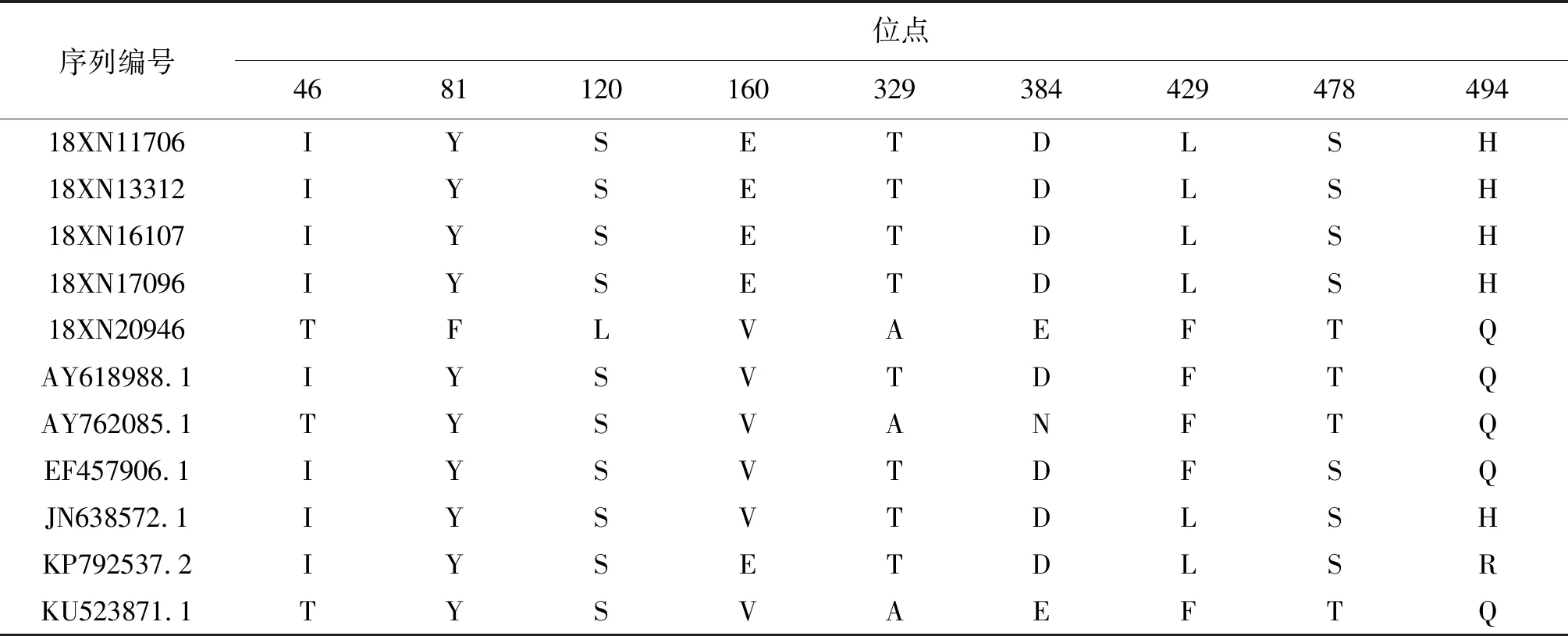
序列编号位点468112016032938442947849418XN11706IYSETDLSH18XN13312IYSETDLSH18XN16107IYSETDLSH18XN17096IYSETDLSH18XN20946TFLVAEFTQAY618988.1IYSVTDFTQAY762085.1TYSVANFTQEF457906.1IYSVTDFSQJN638572.1IYSVTDLSHKP792537.2IYSETDLSRKU523871.1TYSVAEFTQ
表5 广州市2018年输入型DENV-4毒株与参考株NS1蛋白氨基酸差异位点比较
Tab.5 Amino acid substitutions in NS1 gene region among reference DENV-4 strains and 5 imported strains in Guangzhou, 2018
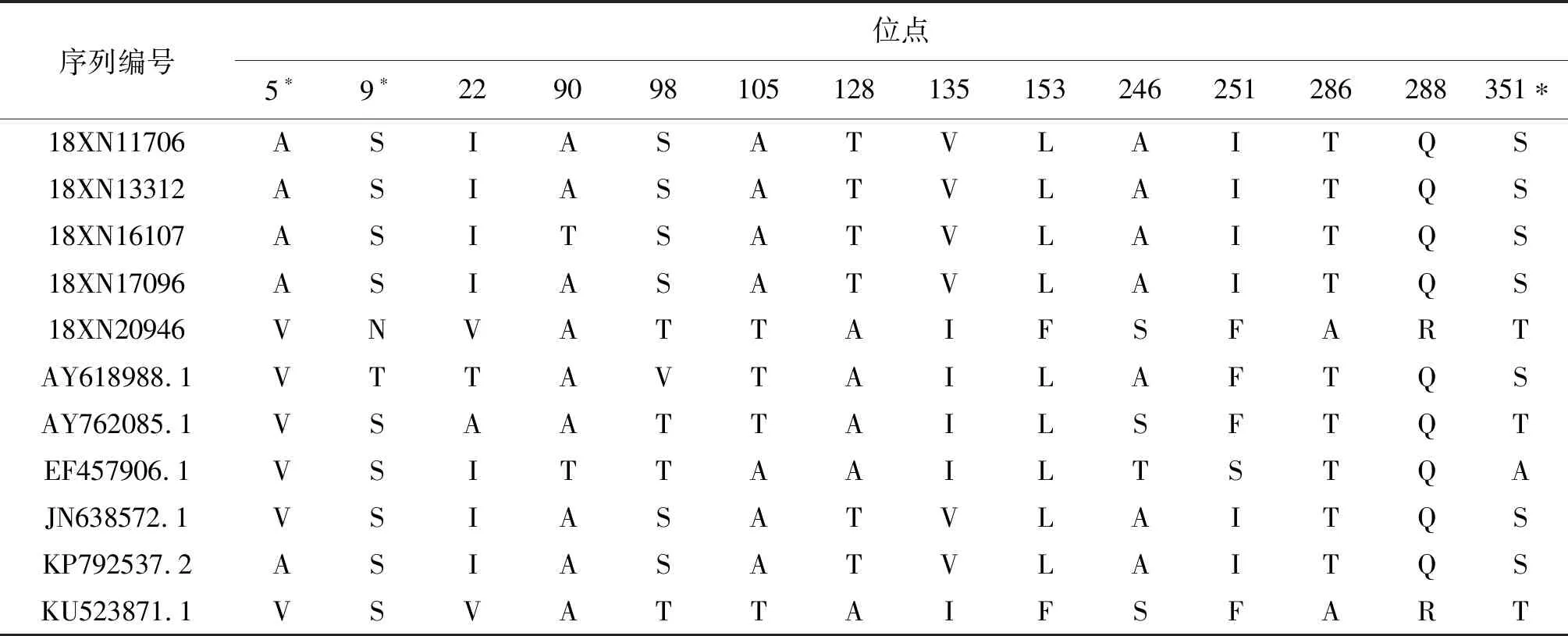
序列编号位点5∗9∗229098105128135153246251286288351∗18XN11706ASIASATVLAITQS18XN13312ASIASATVLAITQS18XN16107ASITSATVLAITQS18XN17096ASIASATVLAITQS18XN20946VNVATTAIFSFARTAY618988.1VTTAVTAILAFTQSAY762085.1VSAATTAILSFTQTEF457906.1VSITTAAILTSTQAJN638572.1VSIASATVLAITQSKP792537.2ASIASATVLAITQSKU523871.1VSVATTAIFSFART
注:*该差异位点位于抗原表位区域
3 讨 论
广州市地处亚热带地区,人口超过1 000万,白纹伊蚊分布广泛;每年的4月份至9月份是广州的雨季,平均气温在20 ℃以上,湿热多雨的气候条件为伊蚊的繁殖生长提供了良好的场所,为登革热的传播创造了理想的条件,因此每年的8月份至10月份通常是广州地区登革热流行的高峰期。卫生死角难以清理,蚊媒密度居高不下,越冬蚊虫无法追踪,是广州市登革热防控的难题[6]。
DENV的4个血清型在东南亚地区常年流行,四季不断,交替循环[9-10];近年来,随着旅游业的发展和国际贸易的日益频繁,广州市与周边地区特别是东南亚地区的往来日益密切,口岸检疫部门每年都有在归国人员和外国入境者中检出DENV阳性者[11]。稍有不慎,输入型病例携带的DENV极易在本地引起暴发甚至流行。以2014年为例,东南亚地区来源的DENV-1型病毒的基因Ⅴ型输入病例在广州市引发了本地大流行,建国以来规模最大的一次城市登革热暴发疫情,病例数达到37 340例[6],DENV-1病毒在广州逐渐呈现本地化的趋势;此外,2016年之后,DENV-2型病毒本地感染病例的比例逐渐升高;此外,近5年来广州市尚未有DENV-3/4引起本地病例的报道。部分暴发街区同时出现了DENV-1和DENV-2型病毒的流行,由于ADE效应的存在,这些街区如果发生2种以上血清型的暴发流行,有极高的概率出现重症登革热病例,使得原本非常严峻的登革热防控变得更加复杂[12]。
表6 广州市2018年输入型DENV-4毒株与参考株NS5蛋白氨基酸差异位点比较
Tab.6 Amino acid substitutions in NS5 gene region among reference DENV-4 strains and 5 imported strains in Guangzhou, 2018

序列编号位点535920122423525626727629633433537237318XN11706YTHVTATIHMVVV18XN13312YTHVTATIHMVVV18XN16107YTHVTATIHMVVV18XN17096YTHVTATIHMVVV18XN20946HSYAIVIVQVIMIAY618988.1HSHATVTIQVIVVAY762085.1HSHATVTIQVIMVEF457906.1HSYAVVTVQVIVVJN638572.1HSYVTATIHVVVVKP792537.2YTHVTATIHVVVVKU523871.1HSHAIVTIQVVMI序列编号位点52552656358663163263564264382689389418XN11706KDIRREHEKIHF18XN13312KDIRREHEKIHF18XN16107KDIRREHGKIHF18XN17096KDIRREHEKIHF18XN20946QETKQDQERTPSAY618988.1KDIKQDQERTLSAY762085.1KDIRQDQERTPSEF457906.1KEIKQDQEKTPLJN638572.1KDVKRDHERTPSKP792537.2KDIKRDHERIHFKU523871.1KDTKQDQERTPS
DENV-4型病毒根据演化路径的不同,可分为Sylvatic、Ⅰ、Ⅱ和Ⅲ一共4个基因型,其中Ⅱ型又可继续分为Ⅱa和Ⅱb 2个分支。在全球范围内,该型别报道较多的2个地区是东南亚地区和南美洲,这2个地区地处热带,都是登革热流行最为严重的地区,4种血清型的DENV常年交替流行[13]。在我国,DENV-4型病毒仅在云南边境有过本地流行的报道,输入来源均是缅甸[14];此外,在2010年,DENV-4输入病例曾经在广州地区引起过暴发疫情,输入来源是泰国[15]。
本研究中分离到的5株DENV-4型病毒均来自输入病例,通过同源性分析和进化树重建可见4株中南半岛输入的分离株属于DENV-4基因Ⅰ型,菲律宾输入的病例分离株属于DENV-4基因Ⅱa型,在进化上分属于不同的种群,这一结论与上述病例的流行病学资料是相吻合的。
DENV包膜蛋白(Envelop, E)基因共编码495个氨基酸残基,该蛋白包含3个结构域(Ⅰ/Ⅱ/Ⅲ),与病毒的受体结合、免疫应答、亲嗜性和相关病理损伤高度相关[16]。本研究分离到的5株DENV-4型毒株在E蛋白结构上共发现了9处氨基酸位点突变,均不涉及3个结构域,提示DENV-4型毒株的E蛋白结构相对稳定。
DENV NS1基因编码352个氨基酸残基,在宿主体内有2种存在形式,即二聚体的膜型NS1和六聚体的分泌型NS1[17]。NS1蛋白是DENV感染的标志性抗原,在急性期患者的血清中可几乎全程检出,对病例的早期诊断具有重要的指导意义。此外,该蛋白可影响病毒RNA复制、免疫逃逸、宿主凝血功能和血管内皮损伤过程。本研究分离的5株DENV-4型毒株在NS1蛋白结构上共有14处氨基酸位点突变,有3处位于抗原表位区域(A5V,S9N,S351T),其中S9N位于FR-I亚区,该变异可能影响NS1蛋白的二级结构,导致病毒NS1蛋白无法形成膜型的二聚体,从而影响宿主保护性抗体的产生,进一步加速病毒的暴发流行。
DENV NS5基因临近3′端,是病毒编码区最长和最保守的区域,其编码的NS5蛋白是病毒基因组编码的最大蛋白(约900个氨基酸残基),同时具有甲基转移酶(methyltransferase, MTase)、核苷酸末端转移酶(terminal nucleotide transferase, TNTase)和RNA依赖性RNA聚合酶(RNA dependent RNA polymerase, RdRp)活性,是病毒复制过程的关键[18]。本研究分离的5株DENV-4型毒株在NS5蛋白结构上共有25处氨基酸位点突变,其中6处位于该蛋白N端(1-265号氨基酸残基),主要影响MTase活性;18处位于该蛋白C端(272-900号氨基酸残基),主要影响TNTase和RdRp活性。
DENV遍布全球热带亚热带地区,每个地区的DENV都有其独特的演化路径和基因特征。以往的监测活动往往忽略了输入性毒株,在本地病例发生的时候无法追溯到相应的输入来源,从而导致流行病学调查信息与分子流行病学溯源不一致的情况。本研究对广州地区输入性DENV-4型毒株进行全基因组测定,间接掌握周边国家地区DENV-4的基因特征,为本地登革热的防控提供基线数据,具有重要的公共卫生意义。
利益冲突:无
