融合暗荧光参数的茄子叶片光合速率预测模型构建
2020-04-27陈丹艳荆昊男张海辉
胡 瑾 高 攀 陈丹艳 李 斌 荆昊男 张海辉
(1.西北农林科技大学机械与电子工程学院, 陕西杨凌 712100;2.农业农村部农业物联网重点实验室, 陕西杨凌 712100;3.陕西省农业信息感知与智能服务重点实验室, 陕西杨凌 712100)
0 引言
光合作用是植物物质积累的核心反应,决定植物干物质积累效果,与作物产量和品质密切相关[1]。光合速率不仅受外界环境的影响[2-4],也受内部生理状态影响[5]。在植物光合速率预测模型研究过程中,YE等[6]在传统光合生理模型的基础上,提出不同类型光响应模型,为光合模型研究打下了良好的基础,但模型中大量生理相关参数难以实时测量,因此不能直接应用于设施环境调控。近年来,出现了面向环境调控的光合模型,早期模型一般未考虑多环境因子对植物光合速率的耦合关系[7]。为此,学者们采用多元非线性回归法建立了多环境因子耦合的植物光合速率预测模型,提高了模型的精度和通用性[8-9],但其在完成多维光合数据拟合中仍存在精度不足。采用智能算法进行光合速率建模可有效提高模型精度,已成为新的研究热点[10-12]。但现有基于智能算法的光合速率预测模型大多未考虑叶片内部生理状态不同引起的光合能力差异,难以用于不同生长状态叶片的光合预测。在此基础上,张海辉等[13]建立了融合叶位差异的作物整株光合速率预测模型,虽在一定程度上提高了模型通用性,但其仅考虑叶位对生长状态的影响,未考虑叶龄、健康状况以及胁迫状态等因素对作物内部生理状态的影响,仍不能真正实现对不同生长状态叶片光合速率的精准预测。
相关研究表明,叶片内部生理状态差异会导致其暗荧光参数发生变化[14-15],而暗荧光参数Fv/Fm表示叶绿体PSⅡ反应中心最大光化学量子效率,与光合速率存在正相关性[16-19],已作为一种研究光合作用机理的探针,用来表征植物叶片光合能力[20-21]。因此,综合考虑外界环境因子和内部生理因子对作物光合能力的影响,建立融合作物叶片暗荧光参数与设施环境多因子的光合模型,为面向作物需求的设施环境调控提供理论依据,是实现作物高效生产的关键之一。
为此,本文在传统人工智能光合速率环境响应模型基础上,引入反映植物本身生长状态的叶片暗荧光参数Fv/Fm,建立融合暗荧光参数与环境因子的光合速率预测模型。首先,采用不同试验预处理方式获得内部生理状态具有一定差异的茄子叶片样本;在此基础上,测试不同样本暗荧光参数,并设计多环境因子嵌套试验,得到不同叶片的多维光合速率数据样本;利用遗传支持向量机算法(GA-SVR)建立融合暗荧光参数和环境变量的光合速率预测模型,以实现对不同生长状态的茄子叶片光合速率统一预测,为设施农业环境因子调控奠定基础。
1 试验材料与方法
1.1 试验材料
本试验于2018年10月—2019年3月在西北农林科技大学农业农村部农业物联网重点实验室(34°7′39″N,107°59′50″E,海拔648 m)进行。以茄子(SolanummelongenaL.,长茄305)为试验材料,采用基质栽培(Pindstrup Substrate,丹麦)于CO2人工气候箱中(RGL-P500D-CO2型,达斯卡公司)进行试验。CO2人工气候箱内环境参数设置:昼、夜光周期为14、10 h,昼、夜温度为25、16℃,昼、夜空气相对湿度为60%、50%,CO2浓度为400 μmol/mol。将长势一致的四叶一心茄子苗54株随机平均分为6组,设置6个光合有效光量子通量密度(PPFD)梯度(表1),其余环境因子及栽培管理均一致。由于不同植株摆放位置不同、同一植株不同枝叶的伸展不一,其与光源距离有差异,叶片实际接受有效光量子辐射不同。处理15 d至茄子植株叶片形态产生明显差异,既存在叶片厚而小,亦存在叶片薄而大的样本。选取各培养箱中差异明显的植株为试验样本进行试验数据采集。

表1 CO2人工气候箱光照强度梯度Tab.1 Gradient of light intensity in CO2 artificial climate charmer
1.2 试验方法
试验以Fv/Fm表征植物内部光合能力,在环境因子选取中,选择与植物光合速率显著相关的外部环境因素——温度、CO2浓度及光照强度构造光合速率嵌套试验[2-4,22],获取建模样本集。为得到暗荧光参数差异明显的建模样本集,本文从每个光照处理的培养箱中随机抽取不同叶位的6片茄子叶片组成一个容量为6的试验样本组,共选取5个样本组。其中每个试验样本组对应一个光合试验温度梯度,其梯度设置为35、31、27、23、19℃。并利用MINI-PAM-Ⅱ型调制叶绿素荧光仪(WALZ公司,德国)测取每个样本组中每片茄子叶片暗荧光参数。试验对通过暗适应叶片夹暗适应20 min后的叶片进行叶片暗荧光参数Fv/Fm记录。共获取不同状态叶片暗荧光参数Fv/Fm30个,其分布范围主要在0.61~0.86之间,符合生长规律并覆盖正常生长叶片的Fv/Fm区间,其具体概率分布如图1所示。

图1 样本数据暗荧光参数的概率分布Fig.1 Probabilistic distribution of Fv/Fm for sample data
同时设计试验测量该组样本叶片于不同CO2浓度和光照强度下的光合参数,采用LI-6800型便携式光合速率测试仪(LI-COR公司,美国)获取待测叶片不同CO2浓度与光照强度条件下的净光合速率。其中光照强度梯度为1 500、1 200、1 000、800、600、300、150、60、30、15、0 μmol/(m2·s),叶室CO2浓度梯度为1 300、1 000、700、400 μmol/mol。由于外界环境短期变化不足以改变植物内部生理状态,叶片暗荧光参数的动态性没有光下荧光变化剧烈[23]。基于LI-6800型便携式光合速率测试仪光响应荧光测量原理,每片试验样本采用相同暗荧光参数[24-26]。
试验通过测定不同条件下的光响应曲线完成数据采集,每次更换环境时,均需对叶室内叶片在1 500 μmol/(m2·s)光照条件下进行20~40 min光诱导,当光合数据平稳后开始测量。由于试验在CO2人工气候箱中进行,可以忽略植物午休等现象的影响,于每日07:00—18:30进行试验。通过粗大误差剔除,共获取1 294组样本数据用于建立模型。
2 模型构建
以试验数据作为样本集,将CO2浓度、光照强度、温度和当前叶片Fv/Fm作为输入,以光合速率作为输出,利用GA-SVR算法构建植物光合模型,算法流程如图2所示。首先对样本数据进行归一化处理,随机选取80%样本数据作为训练集进行模型拟合训练, 20%样本数据作为测试集;然后,利用遗传算法,以测试集泛化能力为适应度,完成支持向量机正则化参数c和内核参数g的寻优并建立GA-SVR光合速率预测模型;最后通过异校验分析模型是否满足要求,选择决定系数最大和均方根误差最小的模型作为最终光合速率预测模型。
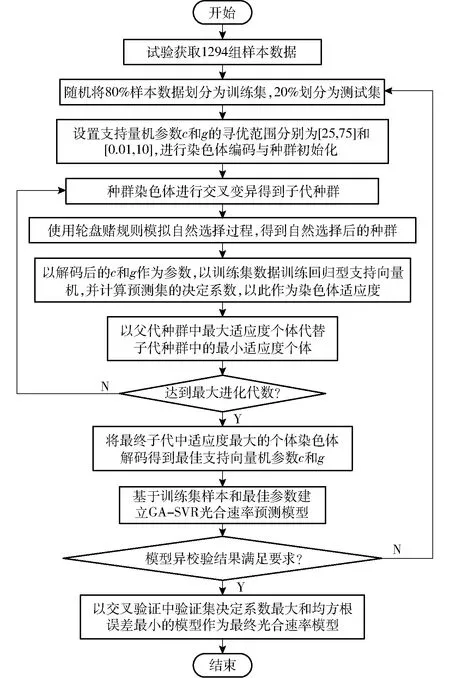
图2 遗传算法优化的回归型支持向量机算法流程图Fig.2 Flowchart of support vector machine based on improved genetic algorithm
2.1 数据预处理
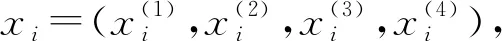
2.2 模型参数选择
2.2.1核函数选择
支持向量机通过Cover定理,利用核函数将低维数据映射到高维空间,用于解决非线性问题。常用核函数有径向基(Rbf)核函数、线性(Linear)核函数、多项式(Poly)核函数及S型(Sigmoid)核函数。为了提高建模精度,选取合适的核函数,比较了不同核函数对模型的影响,结果如表2所示。由表2可知,选用Rbf核函数构建的模型其均方根误差最小、相关系数与决定系数最大,因此选择Rbf核函数作为模型的核函数。

表2 不同类型核函数对模型结果的影响Tab.2 Influence of different types of kernel functions on model results
2.2.2核心参数确定
正则化参数c与核函数参数g对模型拟合结果也有较大影响[27-28]。其中,正则化参数c与模型对误差数据惩罚相关,取值小时模型复杂度小而经验风险大,易造成模型“欠学习”,取值大时模型经验风险小而模型复杂度大,易造成“过学习”现象[29-30]。核函数参数g是支持向量机中Rbf核函数的重要参数,影响核函数形态和支持向量数量。g取值小则核函数选择范围大,支持向量数多,模型的平滑效应小,使得模型无法在训练集得到较高准确率;g取值大则核函数选择范围小,支持向量数少,对未知数据的预测准确率低[31]。
同时,两者对模型精度影响存在耦合效应,为提高模型性能,需精确寻找到参数c和g的最佳值,利用遗传算法进行二维精确寻优操作。为了提升遗传算法全局寻优的精度与速度,首先需利用试参法获得寻优范围。c和g对测试集的均方根误差影响曲线如图3所示。

图3 不同参数对模型归一化均方根误差的影响Fig.3 Influence of different parameters on model
如图3a所示,当参数c取值小于25时,模型均方根误差变化剧烈;在25~75之间时均方根误差变化趋于平稳,且可以取得最小值;而取75以上时模型误差有所增加。因此遗传算法设置c的索引范围为[25,75],同时设置参数c的基因个数为7个。如图3b所示,当参数g在[0.01,10]区间内,模型均方根误差取最小值,因此遗传算法设置g的索引范围为[0.01,10],基因个数为6个。模型采用二进制编码方式,编码公式为
(1)
解码公式为
(2)
式中b——编码后的二进制串
m——染色体所取二进制串字符数量
a——所需编码十进制数
amin——编码空间最小十进制数
amax——编码空间最大十进制数
以解码后的c和g作为参数,进行模型训练,并计算决定系数,以此作为染色体适应度。通过赌注转盘进行种群个体淘汰和选择,每个个体被选中的概率为Pi,计算式为
(3)
式中Fi——种群中个体i的适应度
N——种群大小
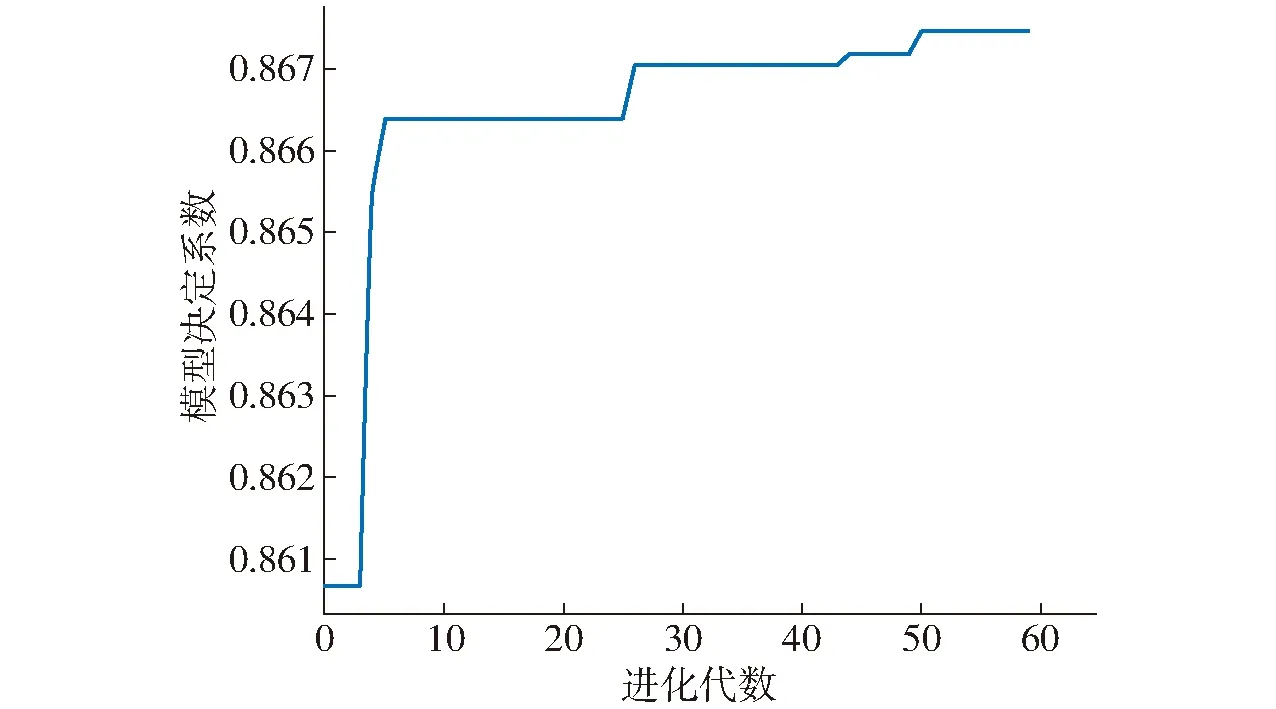
图4 遗传算法进化曲线Fig.4 Improved genetic algorithm evolution curve
在形成的子代中,模拟物种选择过程进行人工干预,以父代的最高适应度个体取代子代最低适应度个体。所形成的子代再通过交叉、变异、选择操作形成新的子代,迭代进化直至最终收敛到最佳解参数c和g。由于人工干预操作降低了种群中的基因多样性,因此应选择适当大的种群变异系数,故选择变异系数为0.2,交叉系数为0.8,遗传算法进化曲线如图4所示。当网络进化到最终收敛时,模型达到最大决定系数,参数c和g最佳解分别为74.61和4.30。由图4可知,模型训练过程中未出现振荡,证明遗传算法对支持向量机参数优化效果明显。
为验证遗传算法对参数的优化效果,采用0.001的寻优步长对参数c和g进行网格遍历寻优,构建了不同c和g条件下的训练集归一化误差曲面,并以遍历最小值点作为误差曲面最低点,如图5所示。可发现曲面最低点坐标(c=74.1,g=4.61,e=0.003 47 μmol/(m2·s))与遗传算法寻优所得误差最低点坐标非常接近(c=74.61,g=4.30,e=0.003 62 μmol/(m2·s)),但其寻优时间是遗传算法寻优的数百倍。遗传算法寻优结果与实际值相比,正则化参数c的相对误差为0.68%,核函数参数g的相对误差为6.72%,所得模型预测数据均方根误差相对误差为4.14%,寻优计算结果与真实最低误差基本吻合,但寻优时间极大缩短,证明在光合速率预测模型构建中使用遗传算法对模型参数进行优化可行。
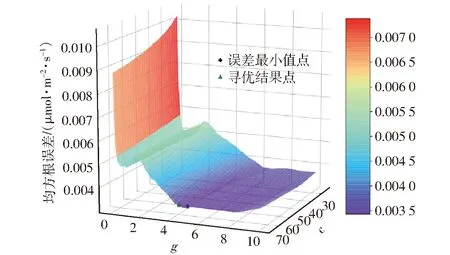
图5 不同c、g参数下的归一化误差曲面Fig.5 Normalized error surface with change of c and g parameters
2.3 预测模型建立
对于给定的训练样本集{(X1,Pn1),(X2,Pn2),…,(Xm,Pnm)},利用GA-SVR进行拟合,得到最终光合速率预测模型为
y=f(X)=ωTX+b′
(4)
式中y——模型输出,即光合速率
ω——模型系数矩阵
X——待变换样本,为(x(1),x(2),x(3),x(4))
b′——模型偏置项
由于样本数据间的非线性关系,GA-SVR通过核技巧将输入向量映射到更高维的空间,使非线性问题转换为线性问题求解,构造回归超平面,实现模型拟合。本文选用Rbf核函数进行输入向量X维度变换,其转换后的输入向量形式为
exp(-g‖X-Xp‖2)
(5)
式中Xp——核函数中心
核函数参数g选用遗传算法进行参数优化后为4.30。
在模型系数矩阵ω的获取中,为使模型输出yi与样本标签Pni尽量接近,引入损失函数L以提高模型精度,定义为
(6)
式中εi、i——松弛变量,为第i样本点的误差
m′——样本总数
取参数c=74.61用于惩罚模型损失。
以拉格朗日数乘法求解损失函数L最小时的模型系数矩阵ω,其转换后为
(7)
将各模型参数代入初始模型式(4),最终获得融合核技巧的光合速率预测模型为
(8)
式中b*——使用核技巧后的模型偏置项
3 模型验证和对比
3.1 建模方法对比
为了验证利用GA-SVR所建立模型的性能,建模时间、训练集拟合精度和决定系数对GA-SVR算法与常用的基于梯度下降法的多项式多元非线性回归算法、BP神经网络算法及以网格支持向量机算法进行了算法性能对比,结果如表3所示。
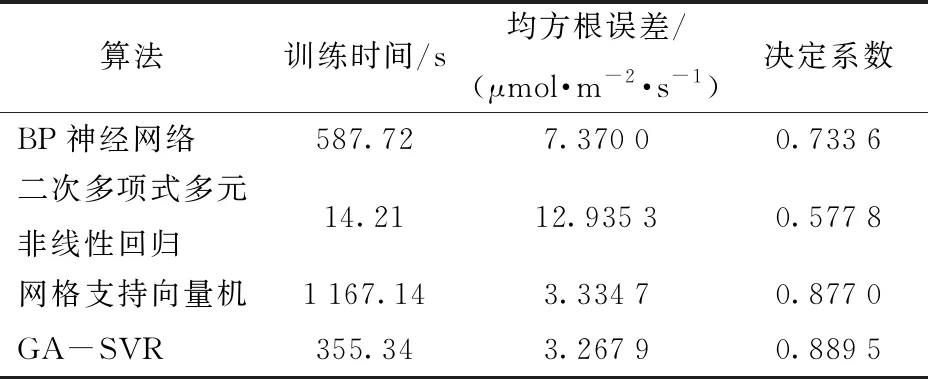
表3 不同算法的性能比较Tab.3 Performance comparison of different algorithms
从表3可发现,二次多项式多元非线性回归算法的模型训练时间远低于BP神经网络算法和网格支持向量机算法,但对于多元大样本数据其拟合精度较低;而网格支持向量机算法在建模精度方面明显优于BP神经网络算法和二次多项式多元非线性回归算法,说明其具有高拟合精度和泛化能力[32],但由于精确建模中模型参数的优化获取过程的影响,其训练速度明显过慢。而GA-SVR算法采用遗传算法对支持向量机参数进行优化,大大缩短了传统SVR模型的训练时间,其训练模型耗时355.34 s。虽然二次多项式多元非线性回归算法耗时最短,但其误差太大,而GA-SVR算法决定系数为0.889 5,均方根误差为3.267 9 μmol/(m2·s),均是上述方法中的最优结果,从而验证了方法的优越性。
3.2 模型验证
为了验证引入荧光参数对模型性能的影响,采用异校验方法,以测试集数据对构建的光合模型和仅考虑环境因子的光合模型进行性能验证。验证结果表明,仅考虑环境因子的模型决定系数和均方根误差分别为0.557 1和13.835 8 μmol/(m2·s);而本文构建的融合暗荧光参数的模型其决定系数与均方根误差分别为0.883 7和2.606 4 μmol/(m2·s),证明引入暗荧光参数极大提高了模型精度。进一步分析包含和不包含荧光参数的光合速率预测模型对未知数据的预测效果,分别获得两模型对测试样本的预测结果如图6所示。
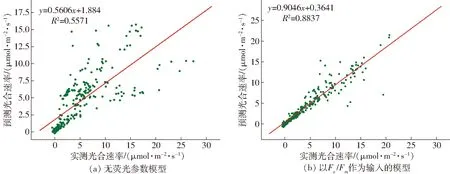
图6 不同输入下的光合速率预测模型拟合效果Fig.6 Fitting effects of photosynthetic rate prediction model under different inputs
从图6a可以发现,若仅以外界光照强度、CO2浓度和大气温度为输入,以光合速率为输出,建立光合速率预测模型,其对测试样本实测值和预测值拟合的斜率为0.560 6,截距为1.884,拟合后决定系数为0.557 1。该模型精度明显不足,难以对不同生长状态叶片进行统一的预测。

图7 不同荧光参数叶片在相同条件下光响应曲线Fig.7 Light response curves of different fluorescence parameters under same conditions
耦合叶片暗荧光参数Fv/Fm和外界环境因子,在相同条件下建立光合速率预测模型,其对测试样本实测值和预测值拟合结果如图6b所示。该拟合直线决定系数为0.883 7,相对于无荧光参数模型有较大提高。且直线拟合系数为0.904 6,接近于1,截距为0.364 1,接近于0。证明融合叶片暗荧光参数的光合模型可显著提升预测精度,对不同状态茄子叶片进行光合速率统一预测。
为进一步验证模型在不同环境的通用性,补充了外界正常环境下生长茄子的模型验证实验,验证样本点共73个,加入荧光参数后,模型预测光合速率与实际光合速率均方根误差为0.547 9 μmol/(m2·s),而未加入荧光参数的模型预测光合速率和实际光合速率均方根误差为1.728 8 μmol/(m2·s)。表明本文模型是各环境下的通用模型。
4 讨论
为分析引入暗荧光参数对模型精度的影响,对比了相同外界环境条件下,不同暗荧光参数叶片的光响应曲线差异,其结果如图7所示。从图7可以发现,在3组不同环境下,暗荧光参数对光响应曲线的影响相似。即在相同外界环境下,Fv/Fm较大叶片的光合速率较大。当未引入叶片Fv/Fm而仅考虑环境因子对光合速率的影响时,对于随机采集的叶片样本,其输入相同而输出不一致,造成所训练光合模型混乱。这类模型对于生长状态不一致的叶片光合速率无法精准预测,其模型泛化能力极差而不能在实际中应用。因此,引入暗荧光参数Fv/Fm对光合速率预测模型有极大影响。分析表明图6a所示模型由于完全忽略了暗荧光参数对光合的影响,对于Fv/Fm差异明显的建模样本,其模型预测精度会显著降低,不能满足对不同生长状态叶片光合速率预测的精度要求。这可能也是过去学者们对光合速率预测模型构建研究过程中,必须选取长势严格一致的植株作为试验材料,并使用同一叶位采集数据的原因[33-34]。
在此基础上,分析暗荧光参数对叶片光合产生影响的原因可以发现,Fv/Fm可指示PSⅡ反应中心处于开放态时光化学的反应效率[35],判断吸收量子中参与PSⅡ反应中心的量子比例,从而间接表征醌循环速度[17-19]。而醌循环的速度决定了光合作用中电子传递速度,影响水裂解速度与光合速率[36]。其值降低表明叶绿体PSⅡ反应中心造成一定损伤[37],使光合能力减弱。因此,植物叶片暗荧光参数的差异会影响其光合速率,将其引入光合速率预测模型能有效地提高模型适用性,与上述结果一致。
5 结论
(1)通过对差异显著茄子叶片进行光合、荧光试验,发现不同生长状态的叶片其暗荧光参数Fv/Fm具有差异,在相同环境下,随着Fv/Fm的增加,作物光合速率呈上升趋势。
(2)基于GA-SVR算法,提出融合暗荧光参数与环境因子的茄子光合速率预测模型。模型训练时间为355.34 s,训练集决定系数为0.889 5,均方根误差为3.267 9 μmol/(m2·s),相较传统网格回归支持向量机算法,优化后模型在训练时间显著降低的前提下,其精度也有所提升。同时,其精度明显优于BP算法和非线性回归算法,说明GA-SVR光合速率预测模型可实现光合速率的精确预测。
(3)采用异校验方式对模型验证,其测试集决定系数为0.883 7,均方根误差为2.606 4 μmol/(m2·s),模型光合速率预测值与实测值拟合斜率为0.904 6,截距为0.364 1,均明显优于仅考虑环境因素的光合速率预测模型,说明本模型可实现对不同生理状态叶片光合速率的精确预测。
