Protective Effect and Underlying Mechanism of Aplysin on Ethanol-Induced Liver Injury in Rats
2020-04-25CHANGZhishangLIUYingSUAiWANGWenchengXUHongweiJIANGYushanLIANGHui
CHANG Zhishang, LIU Ying, SU Ai, WANG Wencheng, XU Hongwei, JIANG Yushan, LIANG Hui,*
(1. Laboratory of Biomedical Center, Qingdao University, Qingdao 266021, China; 2. School of Basic Medicine,Qingdao University, Qingdao 266071, China; 3. School of Public Health, Qingdao University, Qingdao 266021, China)
Abstract: This study aimed to explore the protective effect of aplysin on ethanol-induced liver injury in rats and to explore the possible underlying mechanism from the perspective of the Toll-like receptor 4 (TLR4) signaling pathway. Male Wistar rats were randomly assigned into three groups: normal control, model control and aplysin-treated groups. The rats in the model control and aplysin-treated groups were given ethanol orally at 8 g/(kg·d) for two weeks, and then at 12 g/(kg·d)for another six weeks. The rats in the aplysin-treated group were administered with aplysin at 150 mg/(kg·d) via gavage one hour before ethanol for 8 weeks. After the last administration, all the animals were fasted with free access to water for 12 h and subsequently sacrificed; liver tissues were collected for histological and biochemical assessments; the levels of serum biomarkers for liver damage and endotoxin levels were detected with biochemical assay kits; primary rat Kupffer cells were cultured for assessment of their phagocytic activity with ink phagocytosis test. The mRNA expression levels of CD14, TLR4 and nuclear factor-kappa B (NF-κB) in Kupffer cells were assessed via reversed transcription polymerase chain reaction; the protein expression levels of TLR4, MyD88, NF-κB p65 and tumor necrosis factor-α (TNF-α) in Kupffer cells were assessed via western blotting; the levels of TNF-α and interleukin-1β (IL-1β) in Kupffer cells were measured by enzyme-linked immunosorbent assay. Aplysin relieved alcohol-induced hepatic histopathological changes, decreased plasma endotoxin levels and suppressed the elevation of liver damage biomarkers. Aplysin treatment effectively restored the phagocytic activity of Kupffer cells. Moreover, aplysin significantly decreased the expression levels of CD14, TLR4,MyD88, NF-κB p65 and TNF-α and the concentrations of TNF-α and IL-1β. These fi ndings showed that aplysin exerted a potent hepatoprotective effect, which might be associated with the inhibition of the TLR4 signaling pathway.
Keywords: aplysin; alcoholic liver disease; endotoxin; Toll-like receptor 4 signaling pathway; Kupffer cells
While recreational consumption of alcoholic beverage is common throughout the world, excessive ethanol consumption brings a series of adverse health and social consequences due to its toxicities. Heavy alcoholism could result in both social and health problems. According to the data of World Health Organization, 5.3% of all global deaths were attributable to alcohol in 2016, in which the number of alcohol-attributable fractions for all-cause deaths in Europe was the highest (10.8%)[1]. In the United States (US), there were approximately 90 000 people dying from alcohol-related causes annually from 2006-2010[2]. It has become the third leading preventable cause of death in the US. As one of the common diseases caused by alcohol, alcoholic liver disease remains a major worldwide health problem. Recent study demonstrated a continuously increasing trend of alcoholic liver disease in western countries such as Finland, Ireland and United Kingdom[3]. However, the exact pathogenesis of alcoholic liver disease is yet to be elucidated.
Liver consists of hepatic parenchymal cells and nonparenchymal cells[4]. As one unique type of non-parenchymal cells, Kupffer cells were innate macrophages, which protected liver with its phagocytosis function[5]. Kupffer cells could engulf exogenous substance, such as endotoxin.On the other hand, patients with alcoholic liver disease were frequently accompanied by endotoxemia in clinical study.Alcohol could be contributing to the endotoximia by two mechanisms: 1) Ethanol could inhibit the intestinal motility,change the species and proportion of intestinal flora, and lead to overproduce of endotoxin due to the translocation of intestinal flora. 2) The consumption of alcohol could lead to increased permeability of the intestinal membrane, which increased the portal concentration of blood endotoxin[6-7].
In response to the increased level of gut-derived endotoxin, the TLR4 signaling pathway may be activated[8].Toll-like receptor 4 (TLR4) could specifically recognize lipopolysaccharide (LPS, also known as endotoxin)[9]. In alcoholic liver disease, alcohol may cause overproduce of LPS and LPS was combined with CD14 on the surface of liver Kupffer cells[10]. TLR4 recognized this compound and hel ped it going into Kupffer cells. After TLR4 signaling pathway was fully activated, downstream nuclear factor-kappa B(NF-κB) was also activated through MyD88-dependent pathway[11-12]. Inflammatory cytokines and chemotactic factors(such as tumor necrosis factor-α (TNF-α), interleukin-1β(IL-1β), monocyte chemotactic protein 1(MCP-1) and macrophage inflammatory protein 1(MIP-1)) were produced and released, which would eventually lead to liver damage[13-14].
Aplysin, a member of the bromine-sesquiterpene compounds family, was mainly found in Laurencia tristicha(red alga)[15]. It had attracted much attention due to its potential anti-bacteria, anti-inflammatory, anti-tumor,immunopotentiation and antioxidant effects[16-18]. Our previous work had shown that aplysin could effectively alleviate alcoholic liver injury by modulating the ethanolmetabolizing pathway, attenuating oxidative stress, and inhibiting mito chondrial damage-mediated hepatocyte apoptosis[19]. The current study focused on the potential role of TLR4 signaling pathway in aplysin mediated hepatic protective effect in ethanol-exposed rats.
1 Materials and Methods
1.1 Materials and reagents
Aplysin was extracted and purified from Laurencia tristicha, which was collected from the coast of Naozhou Island in Zhanjiang, China. The identity of samples was confirmed by Dr. Kuishuang Shao (Institute of Oceanology,Chinese Academy of Sciences) and was utilized to study[18-19].Extraction of the dried plant material, fi ltration and isolation of aplysin were described by Ge Na et al[19]. The aplysin purity in the end-product was 97%.
Forty-five male Wistar rats (180-200 g) were obtained from the Experimental Animal and Animal Experiment Center (Qingdao, China).
A commercial 56% (V/V) liquor (Hong Xing Er Guo Tou,Beijing) was selected as experimental alcohol in this study.
Type IV collagenase was purchased from Sigma-Al drich (St. Louis, MO, US). DMEM/12 culture liquid was purchased from Hyclone (Utah, US). Percoll, MTT, ED2 mAb, Giemsa’s staining and ink phagocytosis test kit were purchased from Solarbio Life Sciences (Beijing, China).Biochemical assay kits (aspartate aminotransferase (AST),alanine aminotransferase (ALT), alkaline phosphatase (ALP),cholinesterase (CHE) and gamma-glutamyl transpeptidase(GGT)) were purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). The chromogenic substrate Limulus reagent kit was purchased from Xiamen Limulus Test Reagent Factory (Xiamen, China). The PureLink RNA Mini Kit and mRNA Reverse Transcription System were purchased from Invitrogen (Grand Island,NY, US). The primary antibody against TLR4, MyD88,NF-κB p65 and TNF-α were purchased from Santa Cruz Biotechnology (California, US). The secondary antibody,β-actin and Histone H3 were purchased from Zhongshan Jinqiao Biological Technology Co., Ltd (Beijing, China).enzyme-linked immunosorbent assay (ELISA) kits and SanPrep column DNA glue recycle kits were purchased from Shanghai Jin Ma Laboratory Equipment Co., Ltd. (Shanghai,China) and Cloud-Clone Corp (Texas, US). All primers were synthesized by Sangon Biotech (Shanghai, China).
1.2 Instruments and equipments
Rotary microtome (LEICA RM2135, Germany); Light microscope (Olympus BX60, Japan); Transmission electron microscope (JEOL, Tokyo, Japan); Plate reader (BioTek ELX808, US); Gel imaging system (UVP BioSpectrum,US); Gel imaging system (UVP BioSpectr um, US);GS-700 imaging densitometer (Bio-Rad, US).
1.3 Methods
1.3.1 Animal grouping and administration
Forty-five male Wistar rats (180-200 g) were housed in a temperature- and humidity- controlled environment with alternating 12 h light-dark cycles. After one week acclimatization period on standard laboratory rat chow, the animals were randomly assigned into three groups of 15 rats per group. In the ethanol-model group, the rats were given ethanol daily via gavage at a dose of 8 g/(kg mb·d) for 2 week,and then the dosage was increased to 12 g/(kg mb·d) for the remaining 6 weeks of the study. In the aplysin-treated group,the rats were administered aplysin via gavage at a dose of 150 mg/(kg mb·d) 1 hour before ethanol treatment as described in ethanol model group throughout the 8-week experiment. The dose of ethanol and the dos e of aplysin were based on median lethal dose studies[20-21]and the results of our previous studies[18-19]. Rats in the control group were treated with identical volume of physiological saline for 8 weeks.All animals were sacrificed 12 hours after the last ethanol treatment. In one set of experiment s (n = 10 for each group),blood was collected by aorta ventralis puncture, serum and plasma were isolated for biochemical and endotoxin assays respectively, while livers were collected for histopathological assays. In a second set of experiments (n = 5 for each group),Kupffer cells were isolated and cultured from the livers of the rats. The experimental protocols and the use of animals followed the criteria outlined in the Guide for the Care and Use of Laboratory Animals with the approval of the Animal Care and Use Committee of the Medical College of Qingdao University. All experimental procedures abided by the guidelines of China for animal care, which conform to the principles that are recognized by international communities for the care and use of experimental animals.
1.3.2 Pathological analysis of liver sections
1.3.2.1 Hematoxylin and eosin sta ining
Liver tissues were fixed in 10% neutral formalin immediately after collection for 24 hours, gradually dehydrated through serial ethanol concentrations, and embedded in paraffin. Tissues were serially sectioned at 5 μm with a rotary microtome and stained with hematoxylin and eosin for histological examination under a light microscope.
1.3.2.2 Transmission electron microscopy
Liver tissues (1 mm × 1 mm × 1 mm) were fi xed with 2.5% glutaraldehyde in 0.1 mol/L phosphate buffer (pH 7.2)at 4 ℃ for 4 h. Then the sections were post-fixed in 1%osmium tetroxide in 0.1 mol/L cacodylate buffer (pH 7.2)for 2 h at 4 ℃, rinsed in 0.1 mol/L phosphate buffer ed saline (pH 7.4) three times, dehydrated in a gradient series(30%-100%) of acetone, and then in 100% acetone, infiltrated with EPON812, and fi nally polymerized in pure EPON812 at 60 ℃ for 48 h. Processed samples were observed with a transmission electron microscope.
1.3.3 Assessment of liver function
The activities of AST, ALT, ALP, CHE and GGT in serum were determined using commercial kits following the manufacturer’s protocols. All kits were based on colorimetric methods and were read at a plate reader.
1.3.4 Plasma endotoxin assay
The level of plasma endotoxin was determined using a chromogenic substrate Limulus reagent kit according to the manufacturer’s instructions. All consumables and reagents used were endotoxin-free.
1.3.5 Kupffer cell isolation and culture
Kupffer cells were isolated via digestion with type IV collagenase and purified by Percoll density gradient centrifugation[22]. Briefly, the liver was perfused in situ via the portal vein with 350 mL of Hanks’ balanced salt solution at a fl ow rate of 34 mL/min, and then with 0.5 g/L collagenase at the same flow rate for 20 min. Then the liver was excised and transferred to a 60 mm culture dish,and a cell suspension containing hepatic parenchymal cells and hepatic non-parenchymal cells was formed by gentle disruption o f the collagenase-treated liver. The cell suspension was centrifuged three times at 150 × g for 3 min at 4 ℃. All the collected supernatants were further centrifuged at 800 × g for 5 min to pellet hepatic nonparenchymal cells. The collected cells were separated in a 30%/60% Percoll gradient by centrifugation at 2 000 × g for 15 min at 4 ℃. The cells located between the two gradient densities, which included abundant Kupffer cells, were collected. Finally, Kupffer cells were resuspended in DMEM/F12 with 20% fetal bovine serum and viability was evaluated using the trypan blue exclusion test. The survival rates of cell suspensions were more than 85%.
Kupffer cells from different treatment groups were collected and plated onto 6-well plates at a density of 4 × 106cells per well for reverse transcription polymerase chain reaction (RT-PCR) and ELISA analysis. Cells were cultured in a humidified incubator at 37 ℃ and 5% CO2. The non-adherent cells were removed after 2 hours incubation,and the adherent cells were supplied with fresh medium and cultured for another 18 hours before subsequent analysis.The identity of Kupffer cells was confirmed routinely by immunohistochemical staining with ED2, and Giemsa counterstaining was used to observe the morphology of Kupffer cells. Additionally, the phagocytic activity of Kupffer cells was determined with the ink phagocytosis test.
1.3.6 RT-PCR
Total RNA from Kupffer cells was extracted, and then 500 ng RNA for every sample was reverse-transcribed with the PureLink RNA Mini Kit and mRNA Reverse Transcription System according to the manufacturer’s instructions. PCR amplification was then performed with the following primers:
β-actin (Actb) forward, 5’-GGTATGGGTCAGAAGGACTC-3’;
β-actin reverse, 5’-TGATCTTCATGGTGCTAGGAGCC-3’;
Cd14 forward, 5’-GATCTGTCTGACAACCCTGAGT-3’;
Cd14 reverse, 5’-GTGCTCCTGCCCAGTGAAAGAT-3’;
Tlr4 forward, 5’-CTGAAAATGTGCCTTCAAAACA-3’;
Tlr4 reverse, 5’-GTACTTCAGGTTGTTTGTTC-3’;
Nfkb forward, 5’-GTCACCAAAGACCCACCTCAC-3’;
Nfkb reverse, 5’-CCGCATTCAAGTCATAGTCCC-3’.
Every PCR amplification was i mitated by heating at 94 ℃ for 3 min, followed by 30 cycles of 30 s at 94 ℃, 30 s at 55 ℃, and 40 s at 72 ℃; and 5 min at 72 ℃.The PCR products were electrophoresized through 10 g/L agarose gels with 0.5 mg/L ethidium bromide, and visualized on a gel imaging system. Data were analyzed with Quantity One 4.7. The expression levels of CD14, TLR4, and NF-κB were normalized to β-actin. Three independent experiments were performed.
1.3.7 Western Blotting analysis
Proteins from Kupffer cells were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred onto a polyvinylidene fluoride membrane(Millipore, Bedford, MA, US) for 1 hour with blotting buffer (39 mmol/L glycine, 48 mmol/L Tris-base, 20%(V/V) methanol, 0.037 g/100 mL SDS). The membranes were blocked for 2 hours with TBST containing 10%non-fat milk at room temperature, and then probed with the primary antibody against TLR4 (1:1 000), MyD88(1:1 000), TNF-α (1:1 000), β-actin (1:8 000), NF-κB p65 (1:1 000) and Histone H3 (1:8 000) overnight at 4 ℃.After three 10 min washes in TBST, the membranes were incubated with horseradish peroxidase-conjugated secondary antibody (1:8 000) at room temperature for 1 h. After the membranes were washed three times with TBST, the protein bands were visualized with a SuperECL puls kit.Densitometric measurements of the bands were determined with GS-700 imaging densitometer and Quantity One software.
1.3.8 ELISA for levels of TNF-α and IL-1β
The levels of TNF-α and IL-1β in the Kupffer cell culture medium were determined with commercial ELISA assay kits following the manufacturers’ instructions.
1.4 Statistical analysis
All experiments were repeated three times, and all data were presented as mean ± standard deviation. Statistical analysis was performed with SPSS 17.0 (IBM, US),Statistical significance was determined by One-way analysis of variance. If statistical significance was detected, post-hoc least significant difference test was performed to compare between groups. Results were considered significant whenP-values were less than 0.05.
2 Results and Analysis
2.1 Effects of aplysin on the serum levels of ALT, AST,ALP, CHE and GGT
After 8-week treatment, the levels of serum ALT, AST,ALP, CHE and GGT activities were significantly increased by 33.9%, 18.0%, 56.5%, 25.9% and 30.9% (P< 0.05),respectively, in the model group relative to the levels in the control group. With aplysin administration, the serum levels were significantly decreased (13.0%, 11.5%, 41%, 31.7%,and 12.5%, respectively,P< 0.05) when compared with those in the model group (P< 0.05) (Table 1).
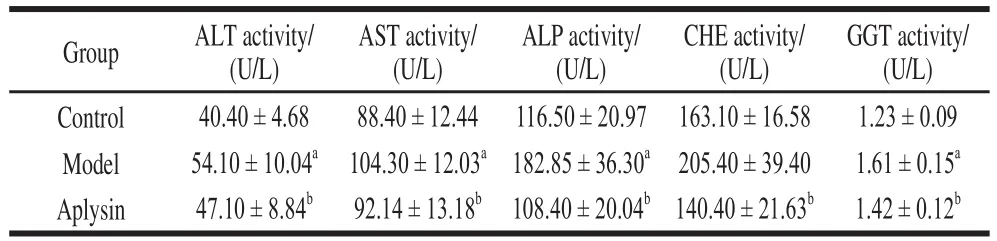
Table 1 Effect of aplysin on ALT, AST, ALP, CHE and GGT activity in serum ( = 10)
2.2 Pathological analysis
Normal lobular architecture and hepatocytes were observed in control rats livers (Fig. 1A). In contrast,remarkable inflammatory response around central vein with vacuolar degeneration and necrosis was observed in ethanol model group (Fig. 1B). Mallary bodies were also present(Fig. 1B). Aplysin treatment significantly alleviated the pathological changes observed in model group (Fig. 1C).

Fig. 1 Effect of aplysin on alcohol-induced histological changes in liver tissues of rats (400 ×)

Fig. 2 Transmission electron microscopic images of liver sections (20 000 ×)
Further assessment of morphology with transmission electron microscopy (TEM) revealed more information:in the control group, smooth nuclear membrane and pores were clearly observed. Morphologically normal cytoplasm,rough endoplasmic reticulum and mitochondria were present(Fig. 2A). On the other hand, dilated endoplasmic reticulum and lysosomes were observed in the model group. Signs of mitochondrial damages such as vague mitochondria ridges were observed as well (Fig. 2B). In the aplysin group, the ultrastructure was generally identical to control group with no significant damage observed (Fig. 2C).
2.3 Effect of aplysin on the plasma level of endotoxin
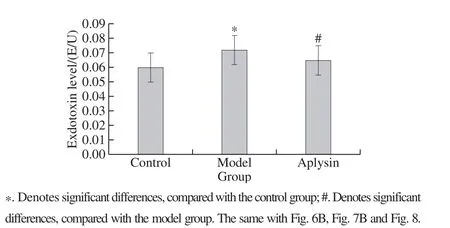
Fig. 3 Effect of aplysin on plasma level of endotoxin (n = 10)
After 8-week treatment, plasma endotoxin in the ethanol model group was significantly increased by 20% relative to that in the control group (P < 0.05). Administration of aplysin significantly abolished the elevation of plasma endotoxin relative to the ethanol group (P < 0.05) (Fig. 3).
2.4 Observation of primary cultured Kupffer cells
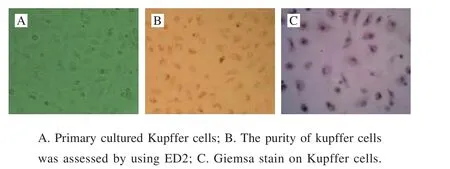
Fig. 4 Observation of primary cultured Kupffer cells (200 ×)
Primary cultured Kupffer cells showed typical shuttle or irregular traiangle shapes (Fig. 4A). The purity was estimated to be more than 97%, as assessed by immunohistochemical staining with ED2 (Fig. 4B). Giemsa staining demonstrated typical “fried egg” appearance (Fig. 4C).
2.5 The effect of aplysin on the phagocytic activity of Kupffer cells
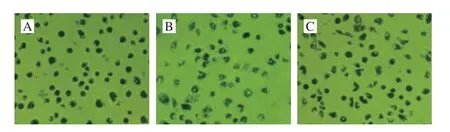
Fig. 5 The effect of aplysin on the phagocytic activity of Kupffer cells (200 ×)
Ink phagocy tosis test revealed that Kupffer cells from control group (Fig. 5A) were enlarged and showed elliptic shape. A large number of Kupffer cells from model group(Fig. 5B) still preserved non-regular shape, such as triangles or diamonds. It revealed that the phagocytic activity of Kupffer cells was decreased compared with control group.Aplysin group (Fig. 5C) effectively restored the phagocytic activity of Kupffer cells relative to model group.
2.6 The effect of aplysin on the mRNA expression levels of CD14, TLR4 and NF-κB in Kuppfer cells
The mRNA expression levels of CD14, TLR4,and NF-κB in Kupffer cells, as measured by RT-PCR,were significantly increased (by 1.1-, 1.7-, and 2.0-fold,respectively, P < 0.05) in the model group relative to the control group (CD14/β-actin, 0.32 ± 0.03; TLR4/β-actin,0.29 ± 0.03; NF-kB/β-actin, 0.22 ± 0.02). The mRNA levels of CD14, TLR4, and NF-κB in the aplysin treatment group were significantly lower (by 38%, 20%, and 23%,respectively, P < 0.05) relative to those in the ethanol model group (P < 0.05)(Fig. 6A, 6B).

Fig. 6 Effect of aplysin on the mRNA expre ssion levels of CD14, TLR4 and NF-κB in Kupffer cells
2.7 The effect of aplysin on the protein expression in Kuppfer cells

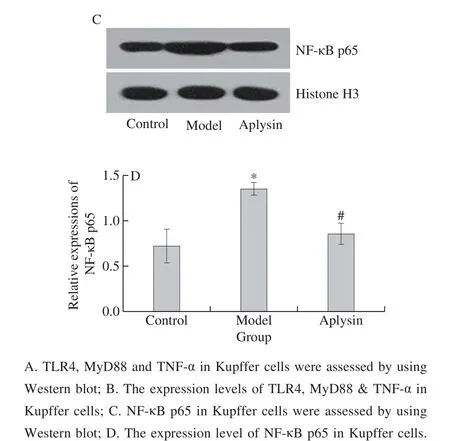
Fig. 7 Effect of aplysin on the protein expression of TLR4 in Kupffer cells
The results of western blotting revealed that the protein expression levels of TLR4, MyD88, NF-κB p65 and TNF-α in ethanol model group were significantly increased(P < 0.05), when compared with the normal control group. The protein expression levels of TLR4, MyD88,NF-κB p65 and TNF-α in aplysin group were significantly decreased (P < 0.05), when compared with those in the ethanol group (Fig. 7).
2.8 The effect of aplysin on the level of TNF-α and IL-1β in Kupper cell culture media
The concentrations of TNF-α and IL-1β in the culture media of Kupffer cells, as measured by ELISA, were significantly increased (by 2.1- and 1.7-fold, respectively,P < 0.05), in the ethanol model group relative to those in the control group (TNF-α (21.86 ± 1.53) ng/L; IL-1β(22.32 ± 1.19) ng/L). The concentrations of those cytokines in the aplysin-treated group were significantly reduced (by 39% and 31%, respectively, P < 0.05) compared with those in the ethanol group (P < 0.05) (Fig. 8).

Fig. 8 Effect of aplysin on the levels of TNF-α and IL-1β in Kupffer cell cultures
3 Discussion
Alcohol is a psychoactive substance with dependenceproperties that has been widely used in many cultures for centuries. Alcohol abuse leads to huge health, social and economic burdens. Of all deaths worldwide, 5.3% are attributable to alcohol consumption; which is greater than many important death causes such as HIV/AIDS (1.8%),violence (0.8%) or tuberculosis (2.3%)[1]. In the US, over 30 000 adults died of cirrhosis in 2016 and half of these deaths were related to alcohol[23]. Though the prevalence of alcoholism among adults in Asia was less than that in western countries, the incidence of alcoholic liver disease, a direct consequence of alcohol consumption, had increased rapidly and also became more severe in China in recent years[1].However, the pathogenesis of alcoholic liver injury has not yet been fully clarified.
Liver histopathology was used to evaluate the therapeutic effect in clinical and animal experiments[24-25].Observation of liver histopathology was the “gold standard”for liver damage evaluation and was widely used in various models of liver damage, such as radiation induced liver injury, chemical induced liver injury, non-alcoholic liver disease and alcoholic liver injury[26-27]. Previous studies had shown that ethanol exposure could lead to histological changes in liver such as the appearance of lipid droplets,hepatocyte swelling and inflammatory cell infiltration into portal duct areas, which reflect morphological and functional damage in alcoholic liver injury[28]. In addition, animals from ethanol model group showed typical characteristics of alcoholic hepatitis like Mallory body. In current study, aplysin attenuated the severity of alcoholic liver injury, as indicated by hematoxylin-eosin staining and transmission electron microscopy. The result was consistent with Xiao Jia[29]and Xu Guangfu[30]et al.’s study.
Liver function/damage biomarkers, such as ALT, AST and GGT, were widely used in clinical and animal study[31-32].In addition, Chen Jianwen et al. reported the activity level of ALP was a more sensitive biomarker than AST and ALT among patients with alcoholic liver disease in 2014[33].Takahashi et al. demonstrated that the activity level of CHE in patients with non-alcoholic liver disease was lower than healthy people[34]. However, the activity level of CHE in patients with alcoholic liver disease was higher than healthy people. Therefore, the activity levels of serum AST, ALT,ALP, CHE and GGT were assessed to confirm the degree of liver injury in current study. The results demonstrated that aplysin could decrease activity levels of serum AST,ALT, ALP, CHE and GGT. In conclusion, aplysin induced hepatoprotective effects in ethanol-exposed rats, as indicated by hepatic histopathological observations and hepatic function-enzymatic assays.
Several lines of research have revealed that gutderived endotoxin plays a crucial role in alcohol-induced liver injury[35]. Endotoxin, a toxic lipopolysaccharide (LPS)component of Gram-negative bacteria in the gut, is released into the bloodstream or tissues such as liver after ethanol consumption because of increased intestinal permeability[36].Ethanol may activate Kupffer cells via increased endotoxin release[37]. In current study, the plasma level of endotoxin in ethanol-treated group was significantly higher than that in normal control group. Meanwhile, aplysin could effectively decrease the plasma level of endotoxin, which was relevant to increased phagocytic ability of Kupffer cells. Kupffer cells,the resident macrophages in the liver, are the main endotoxin responsive cells and the major source of inflammatory cytokine in the liver. Studies had shown that the activation of Kupffer cells by endotoxin is the fi rst event in the process of alcoholic liver injury, and animals that were pretreated with selective Kupffer cell toxicants such as gadolinium chloride did not develop alcoholic liver damage[38]. In current study, cell viability of Kupffer cells in ethanol model group was increased, compared with control group. The result was consistent with Sandahl et al.[37]that aplysin could inhibit the activation of Kupffer cells.
The results of Giemsa staining and immunohistochemistry for ED2 confirmed the successful establishment of primary Kupffer cell culture. Additionally, the results of ink phagocytosis test revealed that alcohol could decrease the phagocytic ability of Kupffer cells and cause liver damage,while aplysin could restore such ability. These data are perfectly consistent with the results of endotoxin level assay,in which aplysin decreased the plasma level of endotoxin.These data suggest that aplysin decrease plasma level of endotoxin by enhancing the phagocytic ability of Kupffer cells.
Exposure to endotoxin could lead to Kupffer cells activation and result in liver damage. In this process, TLR4 signaling pathway plays an important role. Excessive endotoxin can up-regulate the expression of the endotoxin receptors CD14 and TLR4 and activate Kupffer cells by the TLR4-dependent signaling pathway[38]. There are two downstream signaling pathways of TLR4: MyD88 dependent and MyD88 independent. Extracellular, endotoxin binds to TLR4, and TLR4 aggregation causes signal transduction into cells. The intracellular TIR region of TLR4 binds to the carboxyl end of MyD88, and the amino end of MyD88 binds to the amino end of IL-21 receptor-associated kinase(IRAK) to activate IRAK. Activated IRAK reactivates TNF-receptor association factor 6 (TRAF-6), and TRAF-6 activates the inhibitor of NF-κB kinases (IKKs) complex.Inhibitor of NF-κB (inhibitor of NF-κB) is phosphorylated and degraded under the action of IKKs complex[14]. The degradation of IKKs activates NF-κB and induces the expression of specific genes in the nucleus, which leads to the increase of the production of inflammatory cytokines IL-1β and TNF-α[39], which leads to inflammatory reaction and ultimately leads to hepatocyte injury. In the current study,ethanol caused increased mRNA expression of CD14, TLR4 and NF-κB, increased protein expression of CD14 and TLR4 and increased concentration of TNF-α and IL-1β. It can be seen that TLR4 signaling pathway could fully activated by ethanol. In aplysin group, mRNA expression levels of CD14,TLR4, NF-κB, protein expression of CD14 and TLR4 and concentration of TNF-α and IL-1β were all significantly decreased. In conclusion, aplysin enhanced the phagocytic capacity of Kupffer cells, increased the elimination of endotoxin; thus inhibited the activation of TLR4 signaling pathway in Kupffer cells, decreasing the expression levels of CD14, TLR4 and NF-κB and the concentration of downstream inflammatory cytokines TNF-α and IL-1β.
In summary, aplysin had significant effect in rats with alcoholic liver disease and its mechanisms were associated with inhibition of TLR4 signaling pathway.
References: ces:
[1] World Health Organization. Global status report on alcohol and health 2018[R]. Geneva: WHO, 2018.
[2] STAHRE M, ROEBER J, KANNY D, et al. Contribution of excessive alcohol consumption to deaths and years of potential life lost in the United States[J]. Preventing Chronic Disease, 2014, 11: 1-12.DOI:10.5888/pcd11.130293.
[3] MALAGUARNERA G, GIORDANO M, NUNNARI G, et al. Gut microbiota in alcoholic liver disease: pathogenetic role and therapeutic perspectives[J]. World Journal of Gastroenterology, 2014, 20: 16639-16648. DOI:10.3748/wjg.v20.i44.16639.
[4] WERNER M, DRIFTMANN S, KLEINEHR K, et al. All-In-One:advanced preparation of human parenchymal and non-parenchymal liver cells[J]. PLoS ONE, 2015, 10(9): e0138655. DOI:10.1371/journal.pone.0138655.
[5] WEISKIRCHEN R, TACKE F. Relevance of autophagy in parenchymal and non-parenchymal liver cells for health and disease[J].Cells, 2019, 8(1): 16. DOI:10.3390/cells8010016.
[6] PATEL S, BEHARA R, SWANSON G R, et al. Alcohol and the intestine[J].Biomolecules, 2015, 5(4): 2573-2588. DOI:10.3390/biom5042573.
[7] ENGEN P A, GREEN S J, VOIGT R M, et al. The gastrointestinal microbiome: alcohol effects on the composition of intestinal microbiota[J].Alcohol Research: Current Reviews, 2015, 37(2): 223-236.
[8] FRENCH S W, OLIVA J, FRENCH B A, et al. Alcohol, nutrition and liver cancer: role of Toll-like receptor signaling[J]. World Journal of Gastroenterology, 2010, 16(11): 1344-1348. DOI:10.3748/wjg.v16.i11.1344.
[9] MILLER A M, HORIQUCHI N, JEONG W I, et al. Molecular mechanisms of alcoholic liver disease: innate immunity and cytokines[J]. Alcoholism, Clinical and Experimental Research, 2011,35(3): 787-793. DOI:10.1111/j.1530-0277.2010.01399.x.
[10] SURAWEERA D B, WEERATUNGA A N, HU R W, et al. Alcoholic hepatitis: the pivotal role of Kupffer cells[J]. World Journal of Gastrointestinal Pathophysiology, 2015, 6(4): 90-98. DOI:10.4291/wjgp.v6.i4.90.
[11] PETRASEK J, CSAK T, SZABO G. Toll-like receptors in liver disease[J]. Advances in Clinical Chemistry, 2013, 59: 155-201.DOI:10.4291/wjgp.v6.i4.90.
[12] SPENGLER E K, DUNKELBERG J, SCHEV R. Alcoholic hepatitis:current management[J]. Digestive Diseases and Sciences, 2014,59(10): 2357-2366. DOI:10.1007/s10620-014-3173-8.
[13] NEUMAN M G, MAOR Y, NANAU R M, et al. Alcoholic liver disease: role of cytolines[J]. Biomolecules, 2015, 5(3): 2023-2034.DOI:10.3390/biom5032023.
[14] CECCARELLI S, NOBILI V, ALISI A. Toll-like receptor-mediated signaling cascade as a regulator of the inflammation network during alcoholic liver disease[J]. World Journal of Gastroenterology, 2014,20(44): 16443-16451. DOI:10.3748/wjg.v20.i44.16443.
[15] YAMAMURA S, HIRATA Y. Structures of aplysin and aplysinol naturally occurring bromo-compounds[J]. Tetrahedron, 1963, 19(10): 1485-1496.
[16] GONG A J, GE N, YAO W C, et al. Aplysin enhances temozolomide sensitivity in glioma cells by increasing miR-181 level[J]. Cancer Chemotherapy and Pharmacology, 2014, 74: 531-538. DOI:10.1007/s00280-014-2534-5.
[17] GONG A J, GONG L L, YAO W C, et al. Aplysin induces apoptosis in glioma cells through HSP90/AKT pathway[J]. Experimental Biology and Medicine, 2015, 240: 639-644. DOI:10.1177/1535370214555664.
[18] HE J, LIANG H, HAN L, et al. Effect of Aplysin on proliferation and vascular endothelial growth factor expression in human breast cancer cell line[J]. Acta Nutrimenta Sinica, 2009, 31: 482-485. DOI:10.3321/j.issn:0512-7955.2009.05.016.
[19] GE Na, LIANG Hui, LIU Ying, et al. Protective effect of Aplysin on hepatic injury in ethanol-treated rats[J]. Food and Chemical Toxicology, 2013, 62: 362-372. DOI:10.1016/j.fct.2013.08.071.
[20] LIU Y, LIANG H, XU H W, et al. Anticancer activities and immunologic function of Aplysin[J]. Chinese Pharmacological Bulletin, 2006, 22: 1403-1405. DOI:10.3321/j.issn:1001-1978.2006.11.028.
[21] LIANG H, HE J, ZHANG S C, et al. Antitumor activities and its immunologic functions of Laurencia terpenoids[J]. Chinese Journal of Marine Drugs, 2005, 24(1): 6-9.
[22] APARICIO-VERGARA M, TENCEROVA M, MORGANTINI C,et al. isolation of kupffer cells and hepatocytes from a single mouse liver[J]. Methods in Molecular Biology, 2017, 1639: 161-171.DOI:10.1007/978-1-4939-7163-3_16.
[23] TAPPER E B, PARIKH N D. Mortality due to cirrhosis and liver cancer in the United States, 1999-2016: observational study[J]. British Medical Journal, 2018, 362: k2817. DOI:10.1136/bmj.k2817.
[24] HUANG C K, YU T, DE LA MONTE S M, et al. Restoration of Wnt/β-catenin signaling attenuates alcoholic liver disease progression in a rat model[J]. Journal of Hepatology, 2015, 63(1): 191-198.DOI:10.1016/j.jhep.2015.02.030.
[25] ZABALA V, TONG M, YU R, et al. Potential contributions of the tobacco nicotine-derived nitrosamine ketone (NNK) in the pathogenesis of steatohepatitis in a chronic plus binge rat model of alcoholic liver disease[J]. Alcohol and Alcoholism, 2015, 50(2): 118-131. DOI:10.1093/alcalc/agu083.
[26] CHENG W, XIAO L, ANNIWAER A, et al. Molecular responses of radiation-induced liver damage in rats[J]. Molecular Medicine Reports,2015, 11(4): 2592-2600. DOI:10.3892/mmr.2014.3051.
[27] TE J A, ABDULHAMEED M D, WALLQVIST A. Systems toxicology of chemically induced liver and kidney injuries: histopathology-assciated gene co-epression modules[J]. Journal of Applied Toxicology, 2016,36(9): 1137-1149. DOI:10.1002/jat.3278. Epub 2016 Jan 4.
[28] ZHANG M, WU R, JIANG J, et al. The presence of hepatitis B core antibody is associated with more advanced liver disease in alcoholic patients with cirrhosis[J]. Alcohol, 2013, 47: 553-558. DOI:10.1016/j.alcohol.2013.07.003.
[29] XIAO Jia, WANG Jiteng, XING Feiyue, et al. Zeaxanthin dipalmitate therapeutically improves hepatic functions in an alcoholic fatty liver disease through modulating MAPK pathway[J]. PLoS ONE, 2014,9(4): 833-838. DOI:10.1371/journal.pone.0095214.
[30] XU Guangfu, WANG Xinyue, GE Guiling, et al. Dynamic changes of capillarization and peri-sinusoid fibrosis in alcoholic liver disease[J]. World Journal of Gastroenterology, 2004, 10(2): 238-243.DOI:10.3748/wjg.v10.i2.238.
[31] CHIU W C, HUANG Y L, CHEN Y L, et al. Synbiotics reduce ethanol-induced steatosis and inflammation by improving intestinal permeability and microbiota in rats[J]. Food and Function, 2015, 6(5):1692-1700. DOI:10.1039/c5fo00104h.
[32] QU B G, BI W, JIA Y G, et al. Association between circulating inflammatory molecules and alcoholic liver disease in men[J]. Cell Stress Chaperones, 2016, 21(5): 865-872. DOI:10.1007/s12192-016-0711-7.
[33] CHEN Jianwen, LIU Dan, XUE Zhaoyun, et al. Research on therapeutic effect of yangganjiejiu prescription on alcoholic fatty liver and its mechanism[J]. Zhong Yao Cai, 2014, 37(2): 293-298.
[34] TAKAHASHI H, SHIGEFUKU R, YOSHIDA Y, et al. Correlation between hepatic blood flow and liver function in alcoholic liver cirrhosis[J]. World Journal of Gastroenterology, 2014, 20(45): 17065-17074. DOI:10.3748/wjg.v20.i45.17065.
[35] WHEELER M D. Endotoxin and Kupffer cell activation in alcoholic liver disease[J]. Alcohol Research & Health, 2003, 27: 300-306.
[36] TESCHKE R. Alcoholic liver disease: alcohol metabolism, cascade of molecular mechanisms, cellular targets, and clinical aspects[J].Biomedicines, 2018, 6(4): 106. DOI:10.3390/biomedicines6040106.
[37] SANDAHL T D, GRONBAEK H, MOLLER H J, et al. Hepatic macrophage activation and the LPS pathway in patients with alcoholic hepatitis: a prospective cohort study[J]. The American Journal of Gastroenterology, 2014, 109(11): 1749-1756. DOI:10.1038/ajg.2014.262.
[38] GAO B, SEKI E, BRENNER D A, et al. Innate immunity in alcoholic liver disease[J]. Journal of Hepatology, 2019, 70(2): 249-259.DOI:10.1016/j.jhep.2018.10.023.
[39] LI X, WANG Z, ZOU Y, et al. Pretreatment with lipopolysaccharide attenuates diethylnitrosamine-caused liver injury in mice via TLR4-dependent induction of Kupffer cell M2 polarization[J]. Immunologic Research, 2015, 62(2): 137-145. DOI:10.1007/s12026-015-8644-2.
