5年生南方红豆杉生长和分枝性状家系变异与选择
2020-04-16罗芊芊楚秀丽李峰卿徐红兵邓宗付周志春
罗芊芊,楚秀丽,李峰卿,余 明,徐红兵,邓宗付,周志春*
(1. 中国林业科学研究院亚热带林业研究所,浙江省林木育种技术研究重点实验室,浙江 杭州 311400;2. 中国林业科学研究院亚热带林业实验中心,江西 分宜 336600;3. 福建明溪红豆杉产业研究所,福建 明溪 365200;4. 浙江省淳安县富溪林场,浙江 淳安 311700)
南方红豆杉(Taxus wallichiana var. mairei(Lemée H. Léveillé.)L. K. Fu et Nan Li) 为 红豆杉科(Taxaceae)红豆杉属(Taxus L.)喜马拉雅红豆杉(T. wallichiana Zucc.)的变种[1],是集材用、药用和观赏于一体的珍稀濒危树种[2-3],也是我国南方重点推广的珍贵用材造林树种。南方红豆杉在自然条件下生长速度缓慢,再生能力差,很长时间以来,较少有规模的人工栽培。人工栽培的南方红豆杉也因其自然整枝力弱而导致其节子多、易形成多杈干等缺陷影响经济价值[4]。因此,加强其优良品种选育,改良其生长和形质指标,可解决南方红豆杉珍贵用材林生长慢和干材品质差等问题[5]。
对于分布范围广泛的树种而言,长期的地理隔离等会导致天然居群间发生严重的遗传分化,种内不同种源间差异显著[6]。焦月玲等[7]基于南方红豆杉种源幼龄生长测定,发现其树高和地径等生长性状均存在显著种源差异,这为早期选择提供了可能。王艺等[8]在浙江龙泉和浙江安吉两地开展了南方红豆杉种源试验,结果表明,两测试点种源树高、胸径和最大侧枝长等性状变异幅度均超过25%,且方差分析也显示大多数性状指标在种源间呈现显著或极显著差异,说明南方红豆杉种源具有丰富的遗传变异,遗传改良潜力较大,最终从24个南方红豆杉种源中选出 6个适宜于两地的早期速生优良种源。南方红豆杉不仅在种源间存在丰富的遗传变异,且种源内不同个体间也存在显著的遗传差异,在优良种源内开展优树选择能更有效利用种内变异[5]。在优良种源选择的基础上开展优树选择,在利用种源效应的同时,可选择出生长表现突出的优良家系和个体,这也是多数树种遗传改良采用的策略。如辛娜娜等[9]对木荷(Schima superba Gardn et Champ.)、李荣丽等[10]对大叶栎(Quercus griffithii Hook. f. et Thoms ex Miq)及张谦等[11]对樟树(Cinnamomum bodinieri Levl.)等珍贵树种进行优树家系遗传变异研究,结果表明,其家系生长均存在显著遗传变异。因此,为了获得更加优良的珍贵树种种质资源,优树选择及其子代测定必不可少[12]。本文利用设置在浙江淳安、江西分宜和福建明溪3个试验点的5年生南方红豆杉优树家系测定林进行了全林调查,系统研究其幼林生长和分枝性状的家系遗传变异规律及家系×地点的互作等,并初步筛选出一批速生优质家系,为南方红豆杉育种策略制定和实生种子园的留优去劣等提供理论依据。
1 材料与方法
1.1 材料来源与试验地概况
以来自浙江、江西和福建等3个省份、8个产地的南方红豆杉家系为试验材料,2013年在浙江龙泉育苗[12]。2014年将培育的1年生家系容器苗在浙江淳安(29°37' N,119°01' E)、江西分宜(27°49' N,114°41' E)和福建明溪(26°21' N,117°12' E)3个试验点培育成2年生轻基质容器大苗,于2015年2—3月在3个试验点建立南方红豆杉优树家系测定林。3个试验点均属亚热带季风气候区,其中,浙江淳安年均气温17℃,年均降水量1 430 mm,全年无霜期263 d;江西分宜年均气温 17.2℃,年均降水量1 600 mm,全年无霜期270 d;福建明溪年均气温18℃,年均降水量1 800 mm,全年无霜期261 d。浙江淳安点和福建明溪点是栽植在马尾松(Pinus massoniana Lamb.)林冠下,江西分宜点栽植在冰雪灾后的湿地松(P. elliottii Englem.)和马尾松林冠下,3个地点立地条件中等。
1.2 试验设计与测定方法
浙江淳安点和江西分宜点采用完全随机区组设计,5次重复,8株单列小区,福建明溪点为完全随机设计,单株小区,重复40次,栽植密度为2.0 m×2.5 m,每666.67 m2保留上层松树20株左右,郁闭度0.3左右。浙江淳安、江西分宜和福建明溪 3个地点参试的家系数分别为55、39和32个,浙江淳安和江西分宜、浙江淳安和福建明溪及江西分宜和福建明溪的共有家系数分别为38、14和12个,3个测试点共有的家系数为11个。2018年底对3个试验点的南方红豆杉优树家系试验林进行全林测定,测定指标包括树高、地径、冠幅、当年抽梢长、一级分枝数、最大分枝角、最大分枝长和最大分枝粗等。
1.3 数据处理
在统计分析前,依据拉依达准则[13]去除异常数据。以单株测定值为单元进行性状方差分析[13],最大分枝角经反正弦数据转换,一级分枝数经(X+0.5)1/2数据转换。
浙江淳安点和江西分宜点性状单点方差分析的统计模型为:

式中:Yijk代表单点试验第i区组第j家系第k单株的观测值,u为群体平均效应,Bi为第i区组效应,Fj为第j家系的效应,BFij为第i家系和第j区组的互作效应,eijk为机误。
福建明溪点性状方差分析的统计模型为:
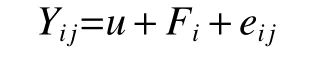
式中:Yij代表单点试验第i家系第j单株的观测值,u为群体平均效应,Fi为第i家系的效应,eij为机误。
浙江淳安点和江西分宜点家系遗传力:

单株遗传力为:
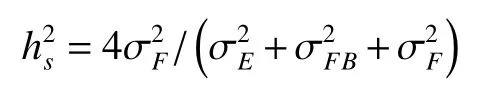
福建明溪点家系遗传力:

单株遗传力为:

采用 SAS8.0 软件的GLM、VARCOMP过程进行性状方差分析,估算性状各变异来源的方差分量、家系遗传力和单株遗传力、性状间表型和遗传相关系数及家系同一性状在不同地点间的简单相关系数。
2 结果与分析
2.1 南方红豆杉生长和分枝性状的家系遗传变异
方差分析结果(表1)表明:各地点5年生南方红豆杉树高、地径、冠幅以及分枝性状都存在极显著的家系变异。在地处较北的浙江淳安点,5年生家系树高和地径变幅分别为1.47~2.06 m和2.07~3.06 cm,平均树高和地径分别为1.73 m和2.57 cm,树高最大家系(闽3)较最小家系(张村)高出40.1%,地径最大家系(三元11)较最小家系(张村)高出47.8%;江西分宜点水热资源更丰富[5],平均树高和地径生长量显著大于浙江淳安点,分别为1.90 m和3.40 cm,家系树高和地径变幅分别为1.44~2.35 m和2.17~4.20 cm,最大家系分别较最小家系高出63.2%和93.5%;福建明溪点分布于南方红豆杉的主产区,但因立地条件与浙江淳安类似,其平均树高和地径生长量也较接近,家系树高和地径变幅分别在1.46~2.29 m和2.26~3.42 cm,最大家系分别较最小家系高出56.8%和51.3%。因南方红豆杉自然整枝能力弱,分枝数少且细的家系选育更具意义。与生长性状一样,南方红豆杉当年抽梢长及一级分枝数、最大分枝角、最大分枝长和最大分枝粗等在家系间的遗传差异也均达到极显著的水平,变异系数为14.9%~43.9%,如浙江淳安、江西分宜和福建明溪3个地点的家系最大分枝粗分别变幅分别为0.70~1.06、0.63~1.29、0.47~0.61 cm。这为早期速生和分枝习性优良的南方红豆杉家系选择提供了很大潜力。
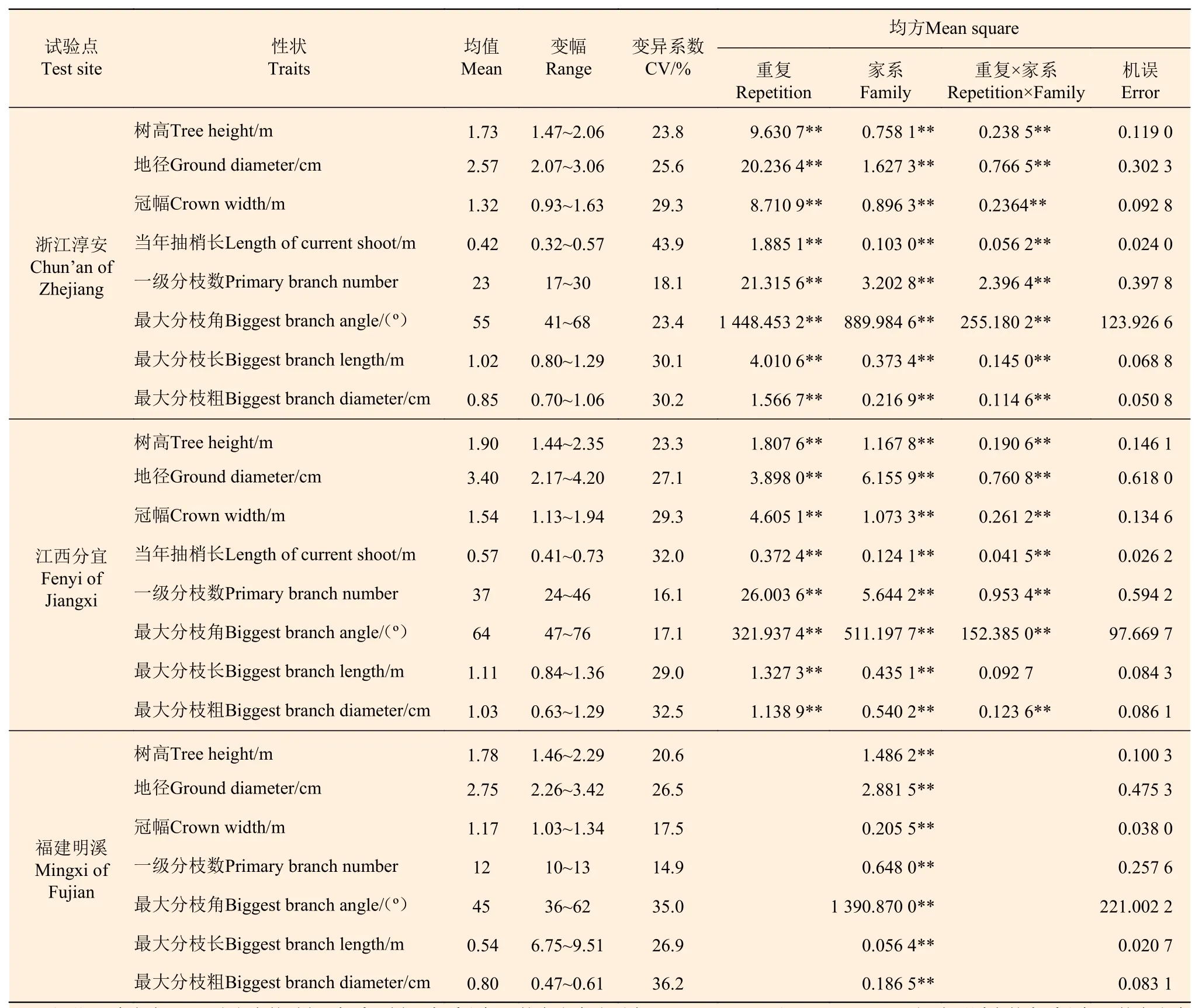
表 1 各试验点南方红豆杉家系生长和分枝性状的方差分析Table 1 Analysis of variance of growth and branching traits of T. wallichiana var. mairei
2.2 南方红豆杉生长和分枝性状的遗传力估算
从表2可看出:5年生南方红豆杉树高、地径、冠幅、最大分枝角、最大分枝长和最大分枝粗在3个地点的家系遗传力均较高(≥0.48),其变幅为0.48~0.88。南方红豆杉生长性状的家系遗传力估算值虽因各地点的测定材料及立地条件不同而有所变化,但总体估算值均较高,如3个地点树高的家系遗传力估算值分别为0.69、0.84和0.87,说明南方红豆杉家系幼林生长受中等至偏强的家系遗传控制。与家系遗传力比较,3个地点南方红豆杉生长和分枝性状的单株遗传力估算值虽然稍低,但仍受中等至偏强的遗传控制。
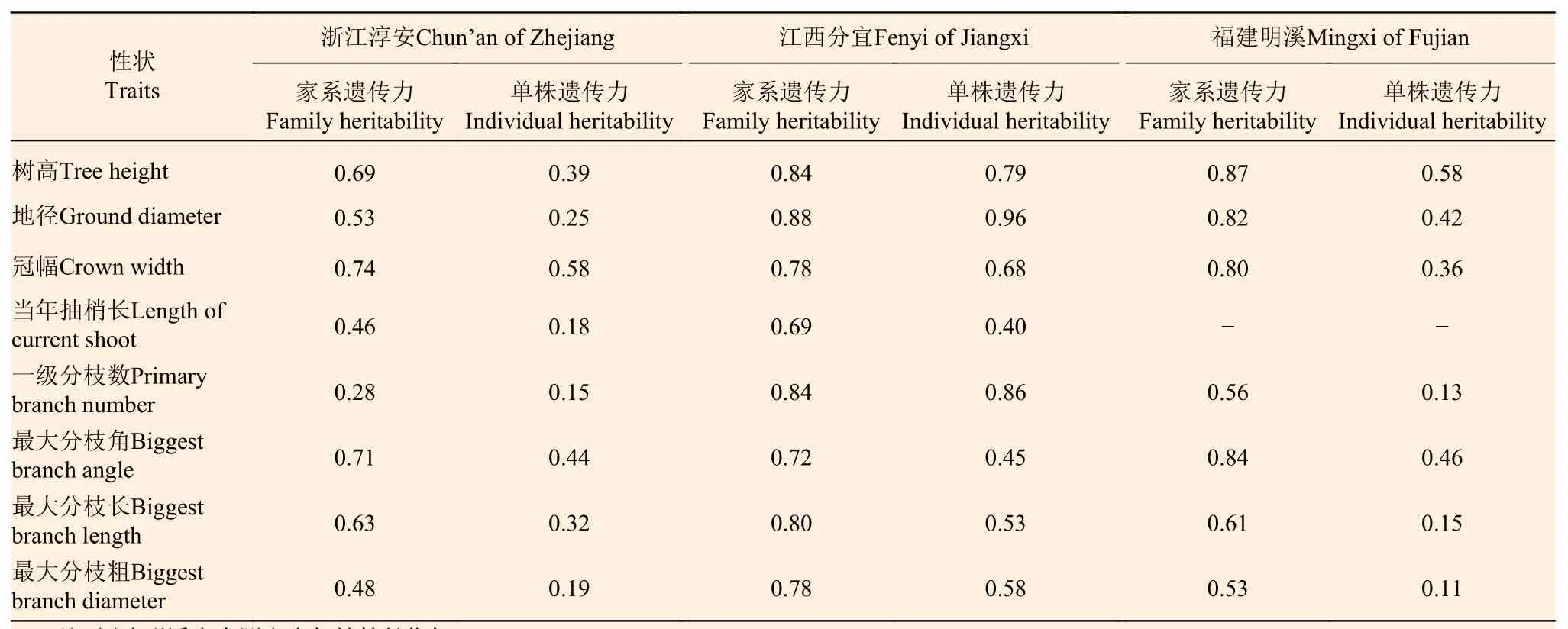
表 2 5年生南方红豆杉生长和分枝性状的遗传力估算值Table 2 Estimation of heritability of growth and branching traits of 5-year-old T. wallichiana var. mairei
2.3 南方红豆杉生长和分枝性状的遗传和表型相关性
表3表明:3个试验点南方红豆杉家系性状间遗传相关系数总体上大于表型相关系数。5年生家系树高、地径和冠幅间呈极显著的正遗传相关(rg=0.799~0.984),各生长性状与一级分枝数和最大分枝粗也呈极显著的正遗传相关(rg=0.495~0.994),这意味着南方红豆杉优树家系树高和地径生长量越大,其分枝越多越粗,冠幅也越大。在江西分宜和福建明溪点,最大分枝角与地径、一级分枝数、最大分枝长及最大分枝粗等性状的遗传相关程度较弱(rg=0.058~0.420),为选择分枝角较大、分枝数少且细的品系提供了可能。
2.4 南方红豆杉生长和分枝性状在地点间的相关性
从图1和表4可以看出:5年生南方红豆杉家系生长和分枝性状在两两地点间的相关性差异较大。在测定的南方红豆杉家系生长与分枝性状中,最大分枝角在3个地点组合间均呈极显著的正相关,表明其家系×地点互作效应不明显,各家系最大分枝角性状稳定,立地条件对其影响较小[14]。而树高和一级分枝数这两个性状在3个地点组合间的相关性都不显著,表明其家系×地点互作效应明显,随着种植生境的改变,家系树高和一级分枝数会发生较大的变化。此外,由表4还可看出:除树高和一级分枝数外,其余生长和分枝性状在浙江淳安和江西分宜两地点间的相关性极显著,其家系与立地互作效应不明显,显示除林冠下光照条件外,这两个地点的立地条件较相似。

表 3 各试验点南方红豆杉家系生长和分枝性状间的遗传和表型相关系数Table 3 Genetic and phenotypic correlation coefficients between growth and branching traits of T. wallichiana var. mairei

图 1 各试验点共有家系树高和最大分枝角在地点间的差异Fig. 1 The differences of tree height and biggest branch angle among all test sites,the traits are from the communal family
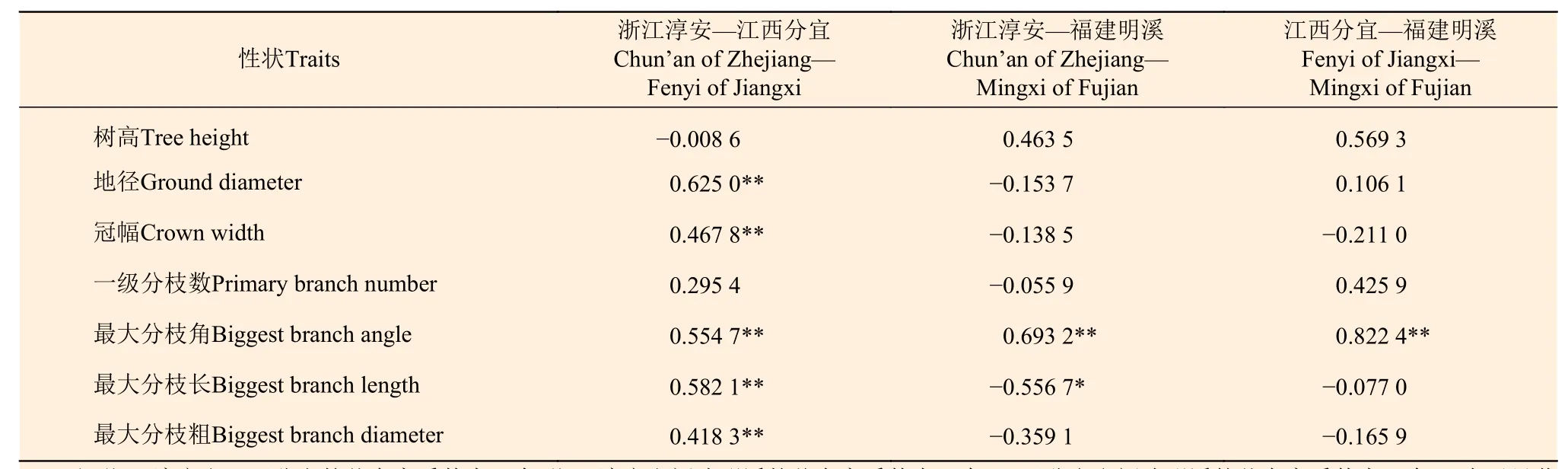
表 4 南方红豆杉家系生长和分枝性状在地点间的相关系数Table 4 Correlation coefficient between growth and branching traits of T. wallichiana var. mairei
2.5 速生优质家系的初选
作为珍贵用材培育目标,南方红豆杉速生优质家系要求速生、节少、分枝细和分枝平展等。树高以高于各地点家系均值为主要选择标准[12],家系最大分枝粗和一级分枝数以低于各地点家系均值为次要选择标准,从浙江淳安、江西分宜和福建明溪3个地点初选出4、5和1个优良家系(表5),其树高均值变幅为1.730~2.100 m。在3个试验点,以SY7、HSX2和SY10 3个家系的综合表现较好,其树高、最大分枝粗和一级分枝数在参试家系中的综合排秩较靠前。在福建明溪点,其入选家系SY10树高1.823 m,高于均值2.53%,最大分枝粗(0.794 cm)及一级分枝数(11枝)分别低于均值1.78%和9.6%,且SY10在江西分宜点也被纳入速生优质家系范围内,可见SY10家系在多个地点皆表现较好且稳定。

表 5 各试验点的优选南方红豆杉家系5年生树高和分枝数值Table 5 Tree height and branch values of preferred families
3 讨论
本文在前期南方红豆杉种源生长和分枝性状变异研究的基础上[12],报道了浙江淳安、江西分宜和福建明溪3个地点5年生南方红豆杉优树家系区域试验测定结果。方差分析结果表明,其生长和分枝性状存在显著的家系遗传变异,且树高和地径的家系变异系数均高达20.0%以上,速生优质家系的选择潜力巨大[15]。3个试验点中,江西分宜点南方红豆杉家系的总体生长表现要明显优于浙江淳安点和福建明溪点,究其原因,可能是由于江西分宜点上层湿地松和马尾松较稀疏,林冠下光照较足,显著促进其分枝生长、冠幅伸展和地径增大。5年生南方红豆杉各性状的家系遗传力普遍较高,受较强遗传控制,类似于欧建德等[16]研究结果;而这3个地点各性状单株遗传力估算值虽然低于其家系遗传力估算值,但仍受中等偏强的遗传控制。同时,江西分宜点的家系遗传力和单株遗传力大体上高于另外两个试验点,这与江西分宜点地处平地,重复间和重复内立地环境相对一致而显著降低了试验的环境效应有关[17]。试验还发现,江西分宜点地径的单株遗传力估算值还出现接近1.00的现象,可能与试验材料不完全符合Stonecypher等[18]的假设,即南方红豆杉自由授粉家系试验群体中包含部分自交或近交后代有关[19]。其实性状遗传力的估算值与所研究的材料、家系多少、年龄、试验地点环境以及试验设计等都有关,其估算值的大小也仅作为参考。
生物不同性状间存在着不同程度的相关,分析性状间的关系可为速生优质家系的选择提供帮助,在选择一个性状的同时,可以间接地知道其它性状的结果。与表型相关比较,遗传相关是在剔除环境效应的影响外,通过方差和协方差统计得到的结果,更能准确地反映由于遗传效应引起的性状间的相关变化[20]。本文性状相关分析表明,江西分宜点和福建明溪点的5年生南方红豆杉优树家系最大分枝角与最大分枝粗及一级分枝数等其它分枝性状相关弱,在遗传上较为相互独立。辛娜娜等[7]在木荷家系研究中也发现其最大分枝角与最大分枝粗不相关或弱相关的规律,这有利于分枝细、分枝角也大即易自然整枝家系的选择。3个试验点家系树高、地径和冠幅三者之间均呈显著的正相关,这些生长性状与一级分枝数和最大分枝粗等也呈极显著的正相关,说明树高和地径生长突出的家系,分枝多且粗、枝叶茂密、冠幅宽[21]。在生产上,可选择速生的家系,再通过合理密植来培育高等级的南方红豆杉优质干材。树高和一级分枝数是2个对立地条件较敏感的性状,其家系×地点互作效应极显著,种植生境对其影响较大,须加强南方红豆杉优树家系的多点测定,估算不同家系的遗传稳定性,才能为不同区域和立地筛选出速生优质和遗传稳定的家系。
在速生优质家系初选过程中,从3个试验点共初选出10个南方红豆杉优良家系,以SY7、HSX2和SY10 3个家系的综合表现较好,且SY10家系在江西分宜点也被纳入速生优质家系范围内,可见SY10家系在多个地点皆表现较好且稳定。在不同地点推广优选家系时,优先选用生长好、稳定性高的家系,可在多个地区造林;对于生长良好而稳定性不高的家系,建议在其生长适宜区域造林;而对于生长较差但稳定性高的家系,其生长适应性较强,对不良环境抵御能力高[22],可用作观景目标来考虑。限于林木生长周期较长,现阶段试验林仍处于幼林阶段,只能对其生长和分枝性状进行初步分析,为后续速生优质品种的多区域推广种植提供一定参考[23]。
4 结论
5年生南方红豆杉生长和分枝性状在家系间的差异均达到极显著水平,具有丰富的遗传变异。3个试验点的南方红豆杉家系遗传力差异较大,且各性状间的相关关系差异也较大,其最大分枝粗、一级分枝数与最大分枝角相关较弱,这有利于易自然整枝家系的选择。种植生境的改变对南方红豆杉家系树高和一级分枝数的影响较大,但对其它生长和分枝性状的影响则较小。从3个试验点初选出的10个南方红豆杉优良家系中,SY10家系在多个地点皆表现较好且稳定。
致谢:参加试验研究的还有中国林业科学研究院亚热带林业研究所金国庆、张蕊,浙江省龙泉市林业科学研究院冯建国、徐肇友、肖纪军、陈焕伟,中国林业科学研究院亚热带林业实验中心曾平生、姚甲宝,福建省明溪红豆杉产业研究所余能健等,谨此谢忱.
