外源性睾酮通过诱导自噬致大鼠肾脏损伤
2020-04-16李旭聂黎虹秦凯悦朱亚洲张航李佳鑫赵瑞宁
李旭 聂黎虹 秦凯悦 朱亚洲 张航 李佳鑫 赵瑞宁
(宁夏医科大学 1研究生学院,宁夏 银川 750004;2总医院泌尿外科;3基础医学院;4 2017级本科生)
肾脏是主要的排泄器官,在维持机体内环境稳态中发挥重要作用。睾酮(TP)是机体中最重要的雄激素,其与肾上腺素受体(AR)结合,在男性的生长、发育、生殖及维持正常生理稳态方面发挥重要作用。AR广泛分布于肾小球和肾小管,TP与其结合后可影响肾脏的结构和功能〔1〕。由于雄激素特殊的作用,部分人群选择外源性给予雄激素以达到男性避孕或提高性欲、运动成绩的目的〔2,3〕。研究表明,长期大剂量使用TP可引起肾小球滤过率显著降低,导致肾脏泌尿功能异常〔4〕。 但目前对于外源性TP导致肾脏损伤的机制鲜有报道。自噬是真核生物细胞调节生长、死亡、能量代谢的重要机制,其广泛参与各器官、系统生理及病理生理过程的发生。研究发现,糖尿病肾病、急性肾损伤等肾脏疾病会通过间接诱导细胞自噬而导致肾脏发生损伤〔5〕。新近研究证明,TP可调控细胞自噬水平〔6〕。但关于外源性TP通过调控细胞自噬参与肾脏损伤发生的过程未见报道。本研究旨在探讨外源性TP是否通过调节细胞自噬参与肾脏损伤的发生。
1 材料与方法
1.1材料
1.1.1动物 选用SPF级SD雄性大鼠48只,体重(340±10)g,由宁夏医科大学实验动物中心提供(许可证号:SCXK(宁)2015-0001)。本研究经宁夏医科大学实验动物伦理委员会批准。
1.1.2主要试剂 TP(北京Solarbio公司,中国);氯喹(CQ,sigma公司,美国);甘草甜素(Gly,东京化成工业株式会社,日本);全蛋白提取试剂盒、聚氰基丙烯酸正丁酯(BCA)蛋白浓度测定试剂盒、ECL检测试剂盒(江苏凯基生物,中国);鼠抗β-actin(北京中杉金桥公司,中国);兔抗HMGB1(Abcam公司,英国);兔抗LC3B(sigma公司,美国);兔抗p62、兔抗Beclin1(proteintech公司,美国);兔抗高迁移率族蛋白B1(HMGB1,Abcam公司,英国);辣根酶标记链霉卵白素工作液(HRP)标记的二抗;苏木素-伊红(HE)、染料(北京中杉金桥公司,中国)。
1.1.3主要仪器 电子天平,显微镜(日本 Olympus公司),高速低温离心机(美国 Thermo公司),切片机(德国SLEE公司,型号CUT5062),全自动酶标仪(美囯Thermo公司)。
1.2方法
1.2.1动物分组与模型建立 将SD雄性大鼠(48只)随机分为6组(每组8只):control组、TP组、TP+CQ组、TP+Gly组、CQ组、Gly组。TP组大鼠皮下注射TP〔3 mg/(kg·d)〕;TP+CQ组大鼠皮下注射 TP〔3 mg/(kg·d)〕加腹腔注射CQ〔50 mg/(kg·d)〕;TP+Gly组大鼠皮下注射TP〔3 mg/(kg·d)〕加腹腔注射Gly〔10 mg/(kg·d)〕;CQ组大鼠仅腹腔注射CQ〔50 mg/(kg·d)〕;Gly组大鼠仅腹腔注射Gly〔10 mg/(kg·d)〕;control组大鼠腹腔注射生理盐水〔2.5 ml/(kg·d)〕。各组大鼠的处理时间均为21 d。
1.2.2标本收集与处理 各组大鼠称重麻醉(10%水合氯醛,0.3 ml/100 g)后,取仰卧位,腹部正中切口,分离并摘取双侧肾脏。一侧肾脏称重后置于4%多聚甲醛中固定;另一侧肾脏冻存于-80℃冰箱。大鼠肾脏指数=肾脏湿重(g)/体重(kg)。
1.2.3HE染色观察肾脏组织形态学改变 取大鼠一侧形态结构完整肾脏用冷生理盐水反复冲洗后,置于4%多聚甲醛中固定,24 h后将其进行修剪后置于包埋盒中脱水过夜;次日进行浸蜡、包埋,冷凝后进行组织切片、烤片;将切片进行脱蜡水化并染色,流水冲洗,梯度酒精脱水,二甲苯透明,中性树胶封片;在光学显微镜下观察肾脏组织病理学变化。
1.2.4组织免疫组化观察自噬指标表达情况 病理切片二甲苯脱蜡,由高到低的梯度乙醇水化,枸橼酸盐缓冲液抗原修复,过氧化氢孵育,山羊血清封闭,各组分别加特异性一抗:抗LC3B-Ⅱ、抗Beclin1、抗P62及抗HMGB1(1∶200稀释)4℃过夜,山羊抗兔二抗免疫球蛋白(Ig)G 37℃水浴锅孵育30 min,HRP 37℃水浴锅孵育30 min,二氨基联苯胺(DAB)显色,苏木素复染,由低到高梯度乙醇脱水,二甲苯透明,中性树脂封固。
1.2.5Western印迹法检测自噬相关蛋白表达水平 取各组大鼠一侧肾脏,全蛋白提取试剂盒提取蛋白,BCA法定量,然后将样品按照50 μg/每孔的上样量加入既定泳道进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳80 min,并通过湿转转移到聚偏氟乙烯膜上。用5%脱脂乳封闭60 min后,将印迹与特异性一抗:抗LC3B-Ⅱ、抗Beclin1、抗P62及抗HMGB1,4℃孵育过夜。磷酸盐吐温缓冲液(TBST)洗涤膜三次后,将印迹与相应的二抗孵育60 min,PBST洗膜三次并用增强的化学发光显色,用凝胶成像仪拍摄。
1.3统计学处理 采用SPSS22.0软件行方差分析。
2 结 果
2.1各组大鼠体重、肾脏湿重及肾脏指数的比较 各组大鼠体重、肾脏湿重、肾脏指数之间均无明显变化(P>0.05),见表1。
2.2各组大鼠肾脏组织形态学分析 control组肾小球大小、形态基本一致;肾小囊间隙均匀;肾小管上皮细胞形态完整且排列整齐,刷状缘和基底纵纹规整,管腔规则。TP组肾小球毛细血管襻分叶明显,襻内细胞数增多;肾小囊间隙变窄;肾小管上皮细胞变矮、排列松散且不规整,基底纵纹模糊不清,刷状缘不规整,大部分官腔闭塞。TP+CQ组和TP+Gly组可见少量且轻微的肾小球毛细血管襻分叶状改变,肾小囊间隙均匀,与TP组比较,肾小管上皮细胞、刷状缘及基底纵纹完整度及排列状况均明显改善。CQ组和Gly组肾脏病理学情况与control组类似,见图1。

表1 各组大鼠体重、肾脏湿重和肾脏指数比较
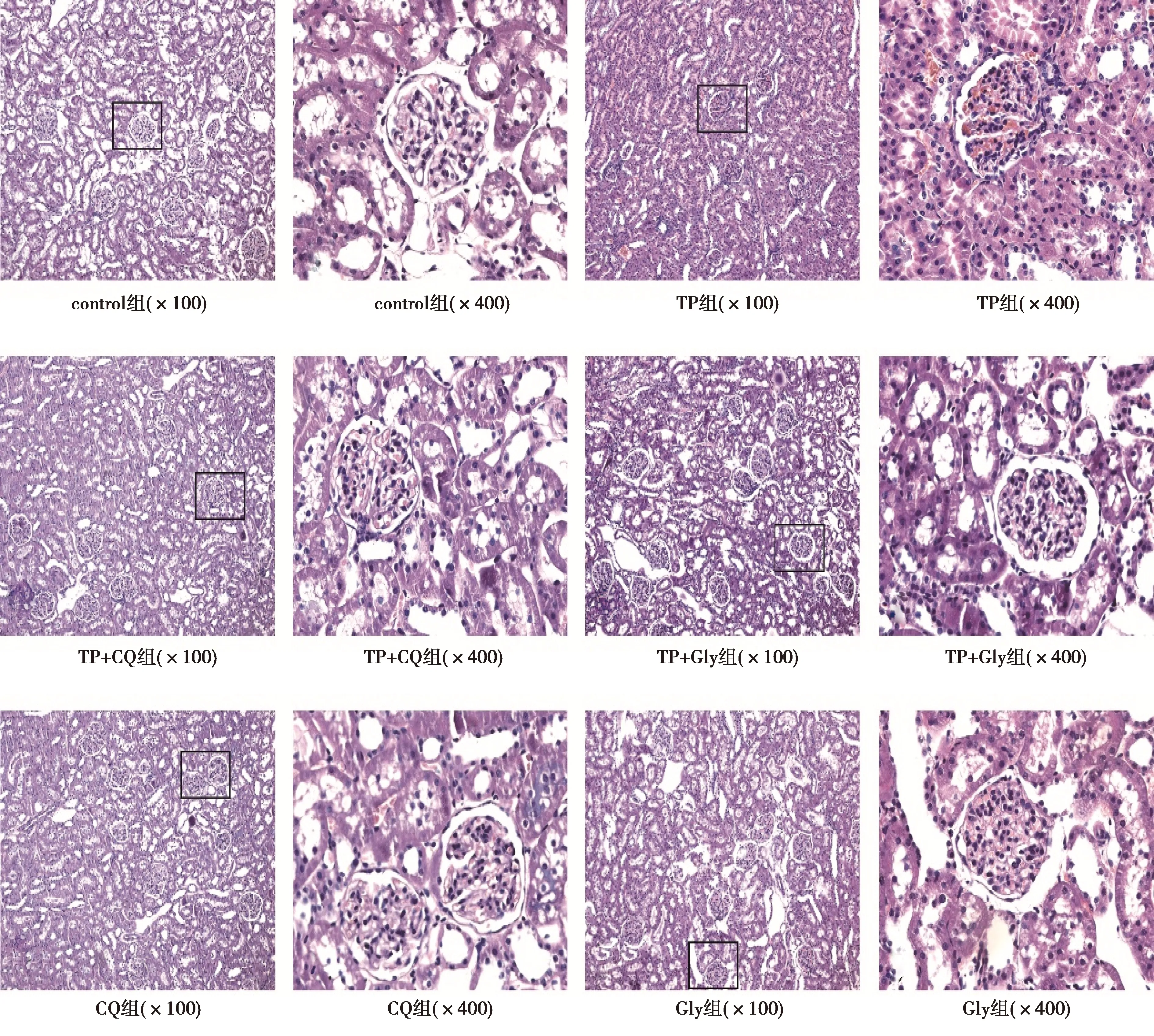
图1 各组大鼠肾脏组织HE染色结果
2.3各组大鼠肾脏组织蛋白免疫组化染色及表达结果 在control组,LC3B-Ⅱ主要表达于肾小管上皮细胞,Beclin1、P62和HMGB1表达于肾小球、肾小管上皮细胞。LC3B-Ⅱ、Beclin1和P62为胞质、包膜阳性,HMGB1为胞核阳性。见图2。与control组比较,TP组大鼠肾脏组织LC3B-Ⅱ、Beclin1、HMGB1蛋白表达水平均明显上调(P<0.05),P62蛋白表达水平无明显变化(P>0.05)。与TP组比较,TP+CQ和TP+Gly组肾脏组织蛋白LC3B-Ⅱ、Beclin1及HMGB1蛋白表达水平均明显下调(P<0.05),P62蛋白表达变化无统计学意义;TP+CQ和TP+Gly组比较各蛋白表达变化均无差异(P>0.05)。CQ和Gly组与control组比较,各蛋白表达变化也无统计学差异(P>0.05),见图3,表2。
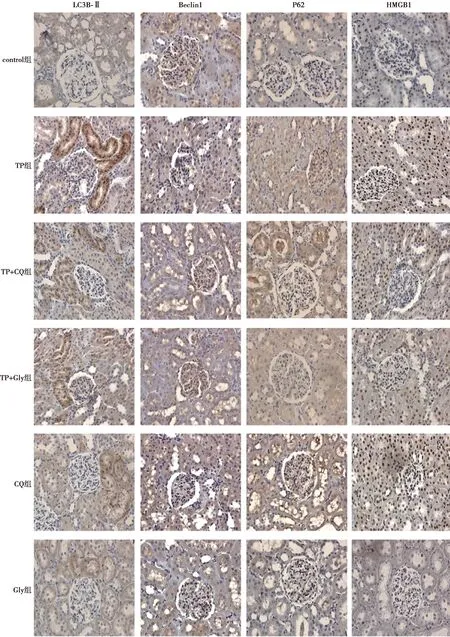
图2 各组大鼠肾脏组织免疫组化染色结果(×400)

图3 各组大鼠肾脏组织LC3B-Ⅱ、Beclin1、P62、HMGB1蛋白的表达结果
表2 各组大鼠肾脏组织LC3B-Ⅱ、Beclin1、HMGB1、P62蛋白表达水平的比较
与control组比较: 1)P<0.05,2)P<0.001;与TP组比较:3)P<0.05,4)P<0.001
3 讨 论
临床上雄激素也可用于治疗再生障碍性贫血等疾病〔7〕。这些对外源性雄激素的给予可能导致肾脏发生慢性损伤,进而影响其正常生理功能。长期应用TP可明显降低肾脏的滤过功能〔4〕;TP也可以增加高血压大鼠肾脏动脉损伤〔8〕。但目前雄激素致肾脏损伤的机制尚未明确。这些现象提示雄激素可能通过调控细胞自噬参与肾脏慢性损伤的病理过程。
本研究提示外源性TP可致大鼠肾脏损伤。同时观察到,TP组大鼠肾脏组织LC3B-Ⅱ、Beclin1表达水平上调。Beclin1是首个被鉴定介导哺乳动物自噬的蛋白,在自噬体形成过程中是不可或缺的条件〔9〕;LC3B-Ⅱ是自噬启动的特异性标记蛋白;两者表达水平增加,提示TP可能通过诱发细胞自噬导致肾脏损伤。随后,联合给予自噬抑制剂,肾脏组织Beclin1、LC3B-Ⅱ蛋白表达下调,肾脏病理损伤部分好转,进一步验证上调细胞自噬水平是TP致肾脏损伤的机制。实验还观察到,TP组HMGB1表达水平升高。HMGB1是重要的自噬调控因子〔10,11〕,不同细胞空间定位的HMGB1均可通过相应机制调控自噬的发生。猜想表达增多的HMGB1可能参与TP致肾脏损伤过程中自噬水平的上调。本研究结果表明在TP致肾脏损伤过程中HMGB1可能通过上调自噬水平参与其中。
综上,外源性TP可能通过上调细胞自噬水平致大鼠肾脏损伤病理过程的发生,HMGB1可能调节了此过程中自噬水平的改变。但在肾脏损伤的病理过程中,睾酮通过何种方式调控自噬,TP和HMGB1在调控自噬过程中是否具有协同作用等问题均需进一步明确。
