中国汉族人群GAD1和GABRB3基因启动子区单核苷酸多态性与精神分裂症的关联研究
2020-04-15张永录丁志杰吴柏林兰小平
李 娜,安 宇,张永录,丁志杰,吴柏林,兰小平
1. 陇东学院生命科学与技术学院,庆阳 745000;2. 复旦大学人类表型组研究院,上海 201203;3. 甘肃省天水市第三人民医院精神科,天水741000;4. 复旦大学生物医学研究院,上海200032;5. 天水师范学院生物工程与技术学院,天水741000;6. 上海市儿童医院,上海交通大学附属儿童医院检验科,上海200040
精神分裂症(schizophrenia,SZ)是一类病因未明的精神类疾病,以妄想、幻想及认知障碍为典型特征,终身患病率为1%[1],遗传度高达79%[2]。目前认为SZ具有高度遗传异质性,不同性别SZ患者的发病年龄和SZ亚型不同[3-4]。
随着分子遗传学技术的发展,已有大量研究发现γ-氨基丁酸(γ-aminobutyric acid,GABA)与 SZ的发病具有相关性。GABA是中枢神经系统中主要的抑制性神经递质,由脑内兴奋性神经递质谷氨酸在谷氨酸脱羧酶(glutamicacid decarboxylase,GAD)的催化下生成。研究[5-7]发现SZ患者死亡后脑组织中GABA活性水平及谷氨酸脱羧酶 67(glutamate decarboxylase 67,GAD67)亚型的mRNA的表达量下降。GAD67由位于2q31.1区的谷氨酸脱羧酶1(glutamate decarboxylase 1,GAD1)基因编码,该基因包含17个外显子。除了合成GABA的限速酶GAD1基因表达量改变引起GABA活性水平下降外,GABA受体也参与SZ的发病。γ-氨基丁酸A受体β3亚基 [γ-aminobutyric acid (GABA)Areceptor β-3,GABRB3]基因、γ-氨基丁酸 A 受体 α5亚基 [γ-aminobutyric acid(GABA)Areceptor α-5,GABRA5] 基因和 γ-氨基丁酸 A受体γ3亚基[γ-aminobutyric acid (GABA)Areceptor γ-3,GABRG3] 基因构成 γ-氨基丁酸 A 受体 [γ-aminobutyric acid (GABA)Areceptor,GABAA]的基因簇,分别编码 GABAA受体的 β3、α5和 γ3亚基。已有研究[8-10]表明GABRB3基因是SZ的易感基因。然而,以往的研究所选的相关基因的单核苷酸多态性(single nucleotide polymorphisms,SNPs)位点大多数来自于基因编码区,主要关注因氨基酸序列改变导致蛋白质功能异常对SZ的影响,很少对基因非编码区SNPs位点进行研究。本文选取GAD1和GABRB3基因启动子区SNPs位点为研究对象,对中国汉族人群中GAD1基因rs3791878、rs3749034位点和GABRB3基因rs4906902位点的多态性与SZ的相关性进行分析。统计手册》(第 4 版)(Diagnostic and Statistical Manual of Mental Disorders,fourth edition,DSM-Ⅳ)诊断标准明确诊断,且样本间不存在亲缘关系。624例正常个体样本(对照组)来自天水市健康志愿献血者,其中男性350名,女性274名,平均年龄(52.22±19.24)岁,所有正常个体均由2名以上精神科副主任医师确认无精神类疾病和阳性家族史,样本间不存在亲缘关系。
1 对象与方法
1.1 研究对象
本研究所有对象均来源于中国汉族人群,545例SZ患者样本(病例组)来自2012年10月至2013年10月在天水市第三人民医院就诊的患者,其中男性310例,女性235例,平均年龄(37.35±12.82)岁。所有SZ患者均由2名以上精神科副主任医师参照《美国精神障碍诊断与
1.2 研究方法
1.2.1 基因选择、启动子区预测及Tag-SNPs的选择 根据已有文献报道[11-12]和蛋白质互作(protein-protein interaction,PPI)在线数据库STRING (http://string-db.org/)的分析结果,选择了在GABA通路中存在互作关系的GAD1和GABRB3基因作为目标基因。通过序列保守性分析预测了GAD1和GABRB3基因的启动子区域,结合1000 GENOMES数据库中的中国北京汉族人群(Han Chinese in Beijing,CHB)和中国南方汉族人群(Southern Han Chinese,CHS)(CHB+CHS=197)的 SNPs数据(dbSNP137),利用Haploview 4.2软件,以条件决定系数r2>0.8、最小等位基因频率(minor allele frequency,MAF) >0.05为标准进行选择,共筛选出 3个标签SNPs(Tag-SNPs),分别是位于GAD1基因启动子区的rs3791878和rs3749034位点及位于GABRB3基因启动子区的rs4906902位点。
1.2.2 样本采集及基因组DNA提取 本研究经天水市第三人民医院伦理委员会批准。受试者均由本人或监护人签署知情同意书后,采集EDTA抗凝外周静脉血3~5 mL,采用QIAamp DNA Blood Mini Kit(QIAGEN,德国)试剂盒提取基因组DNA。
1.2.3 SNaPshot基因分型 采用SNaPshot分型技术检测Tag-SNPs的多态性。在T100TMPCR 仪(Bio-Rad,美国)上 利 用 Qiagen multiplex PCR kit(100)(QIAGEN, 德国)试剂构建多重PCR反应体系,50 μL PCR反应物 包含 0.2 μmol/L PCR 引物(表 1)和 1 μL 模板 DNA,PCR反应条件为:97 ℃ 5 min,35个循环,95 ℃ 30 s,58 ℃90 s,72 ℃ 60 s,72℃ 10 min。经纯化和 SNaPshot单碱基延伸反应后,利用ABI 3130 DNA 测序仪(Applied Biosystems,美国)检测等位基因分布。SNaPshot试剂盒为 SNaPshot master mix(Applied Biosystems,美国)。应用Soft Genetics Gene Marker v 2.2.0 软件进行基因型分析。随机抽取 10%的样本使用 ABI 3130 DNA 测序仪进行双向Sanger测序验证,基因分型一致率为100%。

表1 SNaPshot分型引物Tab 1 Primers for SNaPshot genotyping
1.3 统计学分析
使用 R64 3.5.3 软件分析病例组与对照组间年龄和性别的差异,年龄分析采用两独立样本t检验、性别分析采用χ2检验。应用SNPstats(https://www.snpstats.net/snpstats/start.htm)在线软件进行哈迪 - 温伯格平衡 (Hardy-Weinberg Equilibrium,HWE) 检验、等位基因和不同遗传模式下基因型分布分析。采用Kaplan-Meier统计分析检验rs3791878位点风险基因型G/T与男性SZ患者发病年龄的相关性。样本量统计效能检验使用Quanto 1.2.4软件计算。所有检验均为双侧检验,P<0.05 为差异有统计学意义。
2 结果
2.1 一般人口学资料
病例组共545例,男性310例(56.9%),女性235例(43.1%),年龄14~76岁,平均年龄(37.35±12.82)岁。对照组共624例,男性350例(56.1%),女性274例(43.9%),年龄19~88岁,平均年龄(52.22±19.24)岁。病例组与对照组相比,性别构成差异无统计学意义(P=0.831),年龄差异有统计学意义(P=0.000)。
2.2 统计效能检验
Tag-SNPs中rs3749034 的MAF最小(MAF=0.233)。假设OR=1.5,加性模式,疾病一般人群患病率为1%[1],根据Quanto1.2.4软件计算,结果显示统计效能为0.99。
2.3 Tag-SNPs等位基因分布比较
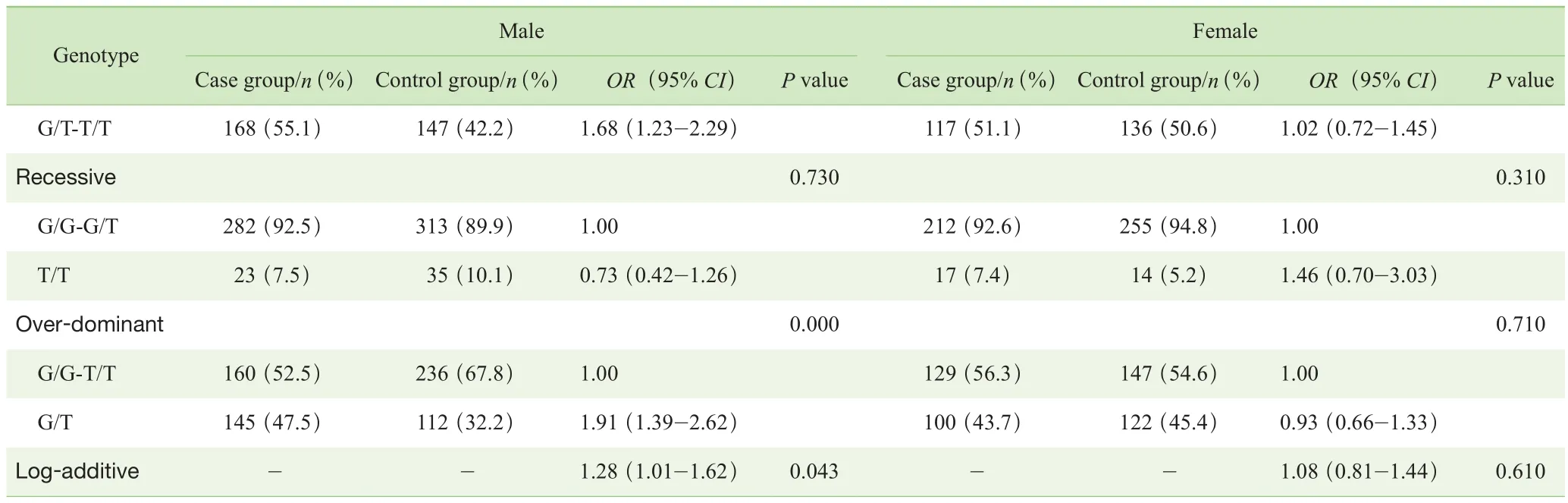
对照组中3个Tag-SNPs位点基因型分布符合HWE检验。由于样本损失,有18例样本的rs3791878位点数据缺失,12例样本的rs3749034位点数据缺失,3例样本的rs4906902位点数据缺失。分别在总体样本和性别分层后样本的病例组与对照组之间对3个Tag-SNPs位点的等位基因分布进行分析。结果显示:总体样本病例组与对照组相比,3个Tag-SNPs位点等位基因分布差异均无统计学意义(P>0.05)(表2);男性病例组与对照组相比,rs3791878位点等位基因分布差异有统计学意义(P=0.039),P值经 Bonferroni校正后,差异无统计学意义(P=0.117);女性病例组与对照组相比,3个Tag-SNPs位点等位基因分布差异均无统计学意义(P>0.05)(表 3)。
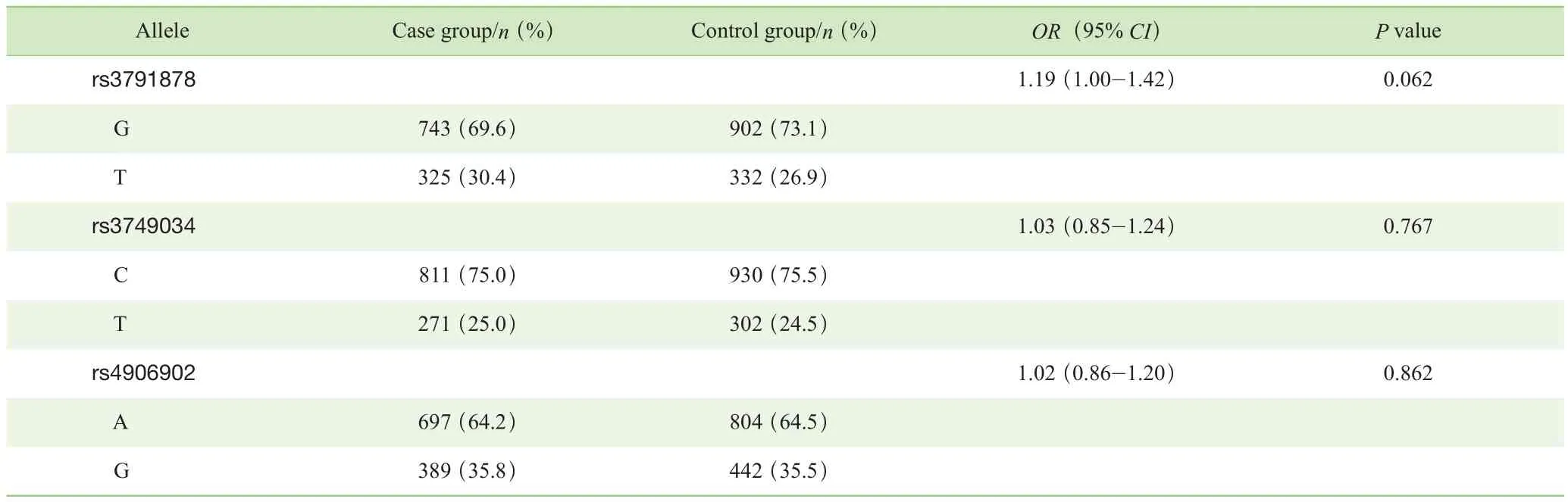
表2 病例组与对照组Tag-SNPs 等位基因分布的比较Tab 2 Comparison of allele distribution of Tag-SNPs between case group and control group
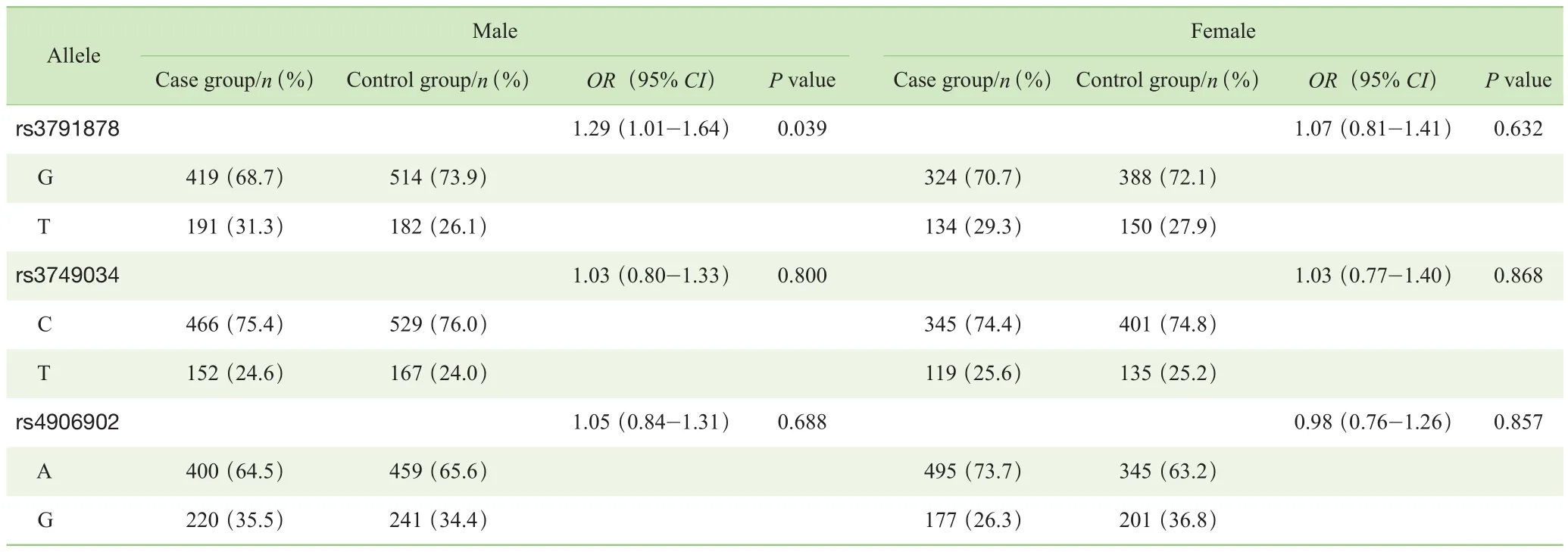
表3 病例组与对照组Tag-SNPs等位基因分布的比较(按性别分层)Tab 3 Comparison of allele distribution of Tag-SNPs between case group and control group (stratified by sex)
2.4 Tag-SNPs基因型分布比较
分别在共显性(co-dominant)、显性(dominant)、隐性(recessive)、超显性(over-dominant)和加性化(logadditive)5种不同的遗传模式下对3个Tag-SNPs位点基因型分布进行分析。结果显示,总体样本的病例组与对照组相比,rs3791878位点在共显性、显性和超显性遗传模式下基因型分布差异均有统计学意义(P=0.022,P=0.011,P=0.006)。挑选赤池信息准则(Akaike Information Criterion,AIC)和贝叶斯信息准则(Bayesian Information Criterion,BIC)数值最小的超显性遗传模式为rs3791878位点可能的遗传模型,该模型下的P值经Bonferroni 校正后,差异仍有统计学意义(P=0.018)。而rs3749034位点和rs4906902位点在5种不同遗传模式下基因型分布差异均无统计学意义(P>0.05)(表4)。在性别分层后样本的病例组与对照组之间对rs3791878位点的基因型分布进行分析,结果显示:男性病例组与对照组相比,rs3791878位点在超显性遗传模式下基因型分布差异有统计学意义(P=0.000),P值经Bonferroni 校正后,差异仍有统计学意义(P=0.000);女性病例组与对照组相比,rs3791878位点在5种不同遗传模式下基因型分布差异均无统计学意义(P>0.05)(表 5)。

表4 病例组与对照组 Tag-SNPs 不同遗传模式下基因型分布的比较Tab 4 Comparison of genotype distribution of Tag-SNPs under different genetic models between case group and control group
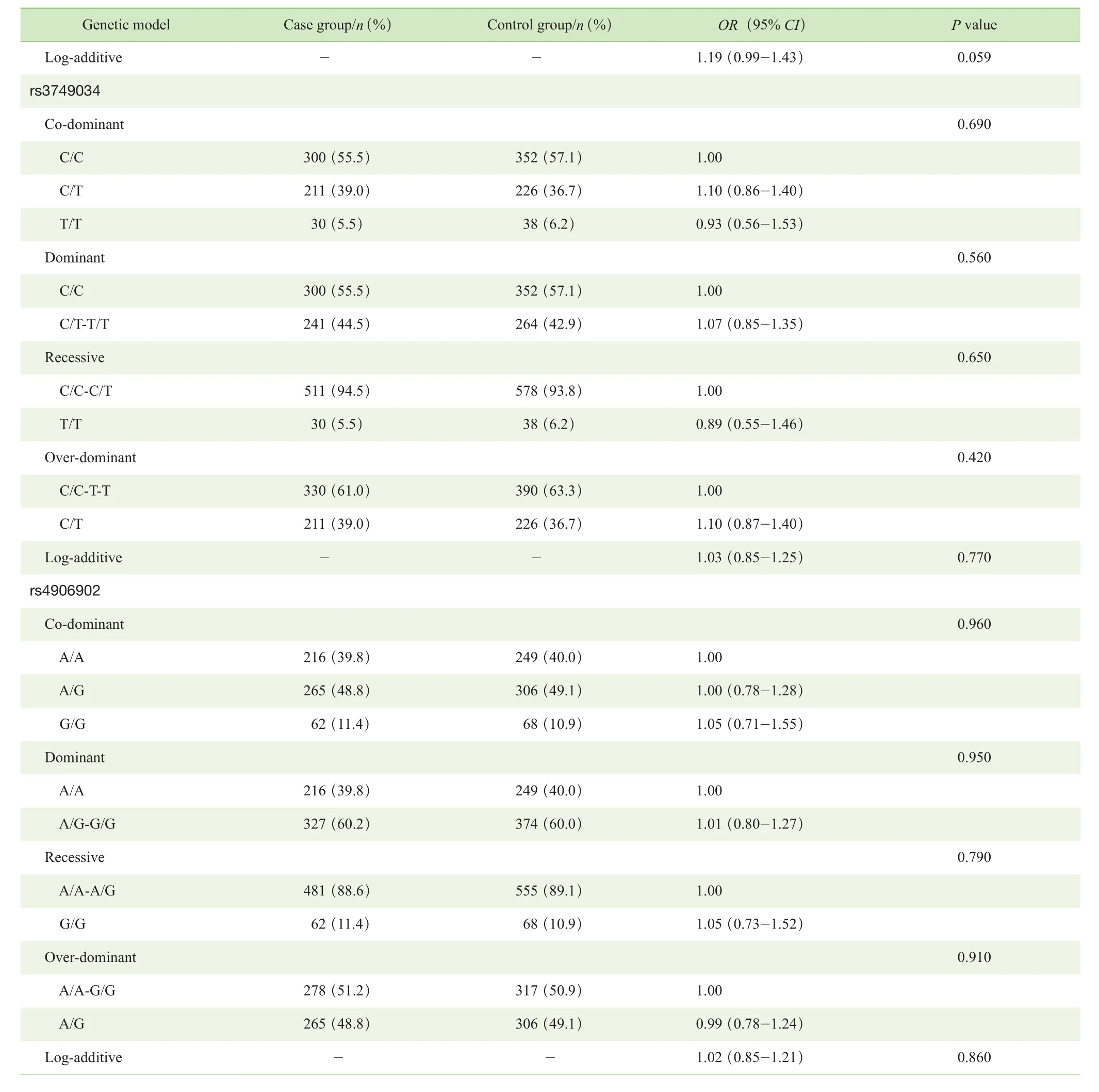
Continued Tab
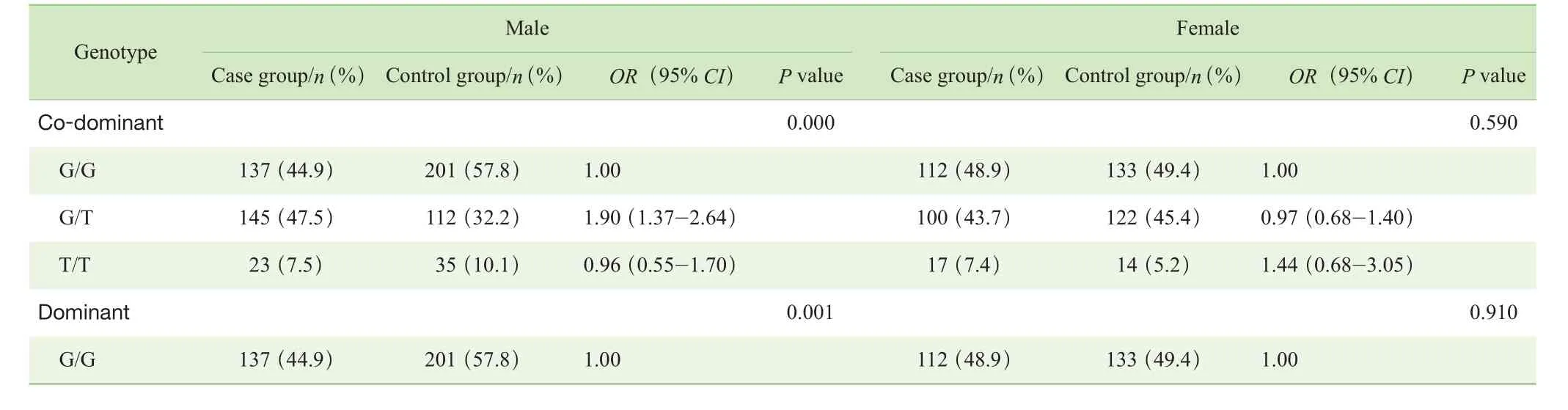
表5 病例组与对照组rs3791878位点不同遗传模式下基因型分布的比较(按性别分层)Tab 5 Comparison of rs3791878 genotype distribution under different genetic models between case group and control group (stratified by sex)
Continued Tab
2.5 rs 3791878 位点风险基因型G/T与男性SZ发病年龄的相关性
310例男性SZ患者共有195例记录了首次发病年龄,应用Kaplan-Meier分析rs3791878位点风险基因型G/T与195例男性患者发病年龄的相关性,结果表明G/T基因型与男性SZ的发病年龄无相关性(χ2=0.271,P=0.603)(图 1)。
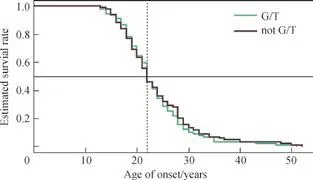
图1 rs3791878位点风险基因型G/T与男性SZ发病年龄的相关性Fig 1 Correlation between the risk genotype G/T of rs3791878 and the age of onset in male SZ
3 讨论
本研究选择GABA通路相关基因启动子区SNPs为研究对象,探索GAD1基因rs3791878和rs3749034位点、GABRB3基因 rs4906902位点多态性与SZ的相关性。PPI分析显示GAD1和GABRB3存在相互作用,且均与溶质载体家族蛋白32成员A1(solute carrier family 32 member 1,SLC32A1)和突触关联蛋白25(synaptosomalassociated protein 25,SNAP25)有直接联系;SLC32A1和SNAP25为GABA通路相关蛋白且与SZ发病相关[13-14]。关联分析结果表明,rs3791878位点在超显性遗传模式下,G/T基因型会增加男性SZ的发病风险,且风险达1.91,但没有得到G/T基因型提前男性SZ患者发病年龄的结果。由于本研究样本量所限,这一结果还需要进一步在更大样本量群体中进行验证。本研究没有发现rs3749034、rs4906902位点与SZ存在相关性。
Du等[15]在2008年对235个中国汉族SZ家系的研究结果也表明GAD1基因rs3791878位点与男性SZ的发生相关。结合本研究,我们认为rs3791878位点存在偏从性显性遗传的可能性,其杂合子的表达受到性别影响,从而增加SZ的发病风险。rs3791878位点在超显性遗传模式下与SZ显著相关,其原因一方面可能与G/T基因型对转录起始因子与启动子序列结合能力的改变有关,最终导致该基因表达量的改变;另一方面可能是G/T会启动不同的蛋白质或不同亚基的表达,导致谷氨酸脱羧酶GAD67亚型活性发生改变,促进SZ的发生。
Straub等[16]认为rs3749034位点突变会导致GAD1基因启动子区产生锌指结构转录因子[ATP1A1 (Na+, K+ATPase α-1 subunit) regulatory element binding factor 6,AREB6] 和成肌分化抗原(myogenic differentiation antigen,MYOD)的结合位点,这2种转录因子能够影响GAD67mRNA的表达。但本次研究没有发现该位点与SZ的相关性。这一结果与Addington等[17]发现rs3749034位点参与SZ早期发病的结果不一致。也有研究[18]发现儿茶酚-O-甲基转移酶(catechol-O-methyltransferase,COMT) 基 因 Val158Met(rs4680)位点与rs3749034位点的相互作用会导致左侧海马旁回皮质的厚度减少,且只有COMTVal158Met突变位点携带者才表现出rs3749034位点变异的阳性结果。这说明该位点可能参与SZ的发生,但并非主导因素。
Urak等[19]认为rs4906902位点的等位基因A突变为G会改变神经元特异性转录激活剂N-Oct-3的结合活性。rs4906902位点与rs8179184位点紧密连锁,单倍型G-T频率在SZ患者组的分布显著高于对照组[20],但本研究没有发现rs4906902位点与SZ的相关性,这可能是因为SZ的异质性所导致的。
本研究结果虽然表明rs3791878位点G/T基因型能够显著增加中国汉族男性人群SZ的发病风险,同时提示GAD1基因是中国汉族男性人群SZ的易感基因,但未涉及机制方面的研究工作。今后仍需大样本研究进一步明确GAD1基因与SZ发生的相关性并开展相关的功能研究,以揭示其致病机制。
