光周期变化对糜子形态建成及幼穗发育进程的影响
2020-04-13董明降彦苗李海权耿玲玲刘建烨乔志红刘国庆
董明,降彦苗,李海权,耿玲玲,刘建烨,乔志红,刘国庆
光周期变化对糜子形态建成及幼穗发育进程的影响
董明1,降彦苗1,李海权1,耿玲玲1,刘建烨2,乔志红2,刘国庆1
(1河北省农林科学院谷子研究所/河北省杂粮重点实验室,石家庄 050035;2河北省蔚县农业农村局,河北蔚县 075700)
【目的】糜子是典型的短日照作物,对光周期反应极其敏感,造成了糜子育成品种生长适应区域狭窄、不能跨区域应用的问题。研究探讨不同光照长度对糜子植株形态建成及幼穗发育的影响,为明确糜子对光周期的反应机理,培育广适应性糜子新品种提供研究依据。【方法】以河北省糜子地方品种二紫杆为材料,采用盆栽试验,出苗后每天补充光照至18 h;设光照0(对照)、10、15、20和25 d共5个处理,每个处理重复3次,18 h光照结束后转移到每天12 h光照人工气候室;田间调查抽穗期,成熟后室内调查株高、茎粗、穗长、穗干重和穗粒干重等农艺性状;利用体视显微镜室内观察糜子幼穗的发育进程并照相记录,统计分析不同光照条件的变化对糜子农艺性状和经济产量的影响及对糜子幼穗分化进程的影响。【结果】每天18 h长日照条件下生长25 d的处理植株平均株高115.6 cm,单穗粒重0.647 g,分别比对照提高了109.0%和472.6%;茎、叶和穗的干物质积累分别比对照提高了416.7%、142.9%和412.0%,差异均达到极显著水平。延长光照20 d处理与25 d处理相比较,各器官干物质重差异不显著。在18 h长日照条件下,糜子茎尖生长点一直处于未伸长期,持续营养生长,生殖生长滞后;在12 h短日光照条件下5 d后糜子开始从营养生长转入生殖生长,幼穗开始分化,分化过程可分为生长点未伸长期、生长点伸长期、穗枝梗原基分化期、小穗原基分化期、小花原基分化期、雌雄蕊原基分化期和花粉粒形成期等共7个时期。所有5个增加每天光照时间和长日照天数的不同处理,幼穗分化持续时间均为15 d左右,没有受到前期延长光照时间的影响。【结论】短日照条件下,出苗后增加每天光照时间和增加长日照天数能延长糜子的营养生长期,有利于干物质的积累,可以极显著的提高糜子的生物和经济产量;短日照条件能够诱导和促进糜子从营养生长到生殖生长的转变,而长日照条件则抑制糜子生殖生长的开始;糜子的幼穗分化可以划分为7个不同的时期,生殖生长开始后不受前期延长光照时间的影响,持续时间在15 d左右。
糜子;光周期;形态;幼穗发育
0 引言
【研究意义】糜子(L.)是起源于中国最早驯化利用的作物之一[1],栽培历史10 000年以上[2],具有耐干旱[3]、耐贫瘠、较高的水资源利用效率[4]等重要特性,是长城沿线风沙区重要的粮食作物,特别对降水较少地区的种植结构调整和产业发展中起着不可替代的作用。糜子营养丰富[5-6],富含淀粉、蛋白质、维生素、膳食纤维及矿质元素[7]。另外,糜子生长周期短,播期调节能力强,是填闲补种、抗灾救灾的首选作物。但是糜子是比较典型的短日植物,在短日照条件下营养生长周期短,开花早,往往造成植株矮小,穗小粒少,产量较低。也是造成糜子品种适应区域狭窄,生产上难以培育出适合多地区种植的广适应性的品种的主要原因。进行糜子对光周期反应的研究对于解决上述问题,进一步探讨糜子光周期反应的机理都具有重要的现实和理论意义。【前人研究进展】光周期是指昼夜周期中光照期和暗期长短的交替变化。在许多植物物种中,开花时间受到光周期的精确调控,确保植物在特定环境下的最佳时机开花以保证产量。延长植物的光照时间,可以促进植物的光合作用,增加产量,同时对幼苗形态也有很大影响[8]。在对甜椒[9]、红皮云杉[10]、番茄[11]、海滨雀稗[12]等进行的增加光照长度的研究中,一般认为植物接收的光照时间越长,进行光合作用的时间相对越长,光合产物的积累量越多,越有利于植物的生长。菊花是短日照植物,在长日照条件下,叶片中会产生TFL(TERMINAL FLOWER)[13]蛋白,可以通过韧皮部从叶片运输至茎尖,直接抑制成花素复合物活性来抑制植株开花,从而保持营养生长[14]。当出现短日照循环条件,TFL蛋白消失,FT蛋白表达,从而促进植株开花。糜子对光照长度的变化反应较为敏感,短光照条件可诱导糜子迅速开始生殖生长,导致营养生长不足,株高降低,叶片数减少,穗小粒少,产量受到严重影响[15]。【本研究切入点】目前,不同日照长度条件对糜子生长发育影响的研究很少,同时国内对于幼穗发育的研究主要集中于小麦[16]等主要农作物,而对糜子幼穗发育研究只在20世纪80年代进行了初步观察[17],但限于当时的科研条件,未能清晰、系统地描绘糜子幼穗发育过程。因此,研究光周期变化对糜子生长发育的影响,观察完善糜子幼穗分化过程,可为研究糜子开花机理,培育广适应性新品种提供帮助。【拟解决的关键问题】本研究分析不同日照长度条件对糜子植株生长发育的影响;明确糜子幼穗分化进程。为今后培育适合多地区种植的广适应性新品种,进一步探讨糜子光周期反应的机理研究奠定基础。
1 材料与方法
1.1 试验材料
以河北省主栽的糜子地方品种二紫杆为供试材料。
1.2 试验方法
试验于2018年3月在河北省农林科学院谷子研究所郄马试验站进行。试验采用盆栽方式进行,盆口直径22 cm、高17cm的白色花盆中,每个处理种3盆。以大田土﹕营养土=2﹕1的混合土作为基质,每盆基质3.5 kg,每盆施纯N 2 g、P2O51.5 g和K2O 1.5 g,一次性作基肥施入。播种前浇透水,使土壤充分吸水达到饱和状态,播种后覆土2 cm,置于温室,光照长度每天18 h(LED灯,增加光照6 h);设置5个处理,播种后0(对照)、10(LD10)、15(LD15)、20(LD20)和25 d(LD25)的植株分别移至人工气候室,每天光照12 h,每个处理3次重复。播种日期为2018年3月23日。
1.2.1 性状调查 按照每个处理条件下抽穗50%以上植株记录抽穗期。在成熟期,每个处理取10株样品测定株高、主茎粗和主穗长。将穗子从穗茎节处剪下,105℃杀青1 h,然后放入80℃烘箱烘至恒重,记录穗重和穗粒重。
1.2.2 幼穗观察 糜子出苗后,选择同一天出苗的植株进行标记。每间隔1—2 d,选取不同处理植株3—5株,剥去样品苞叶露出茎生长点置于体式显微镜(Olympus SZX16,日本)下观察生长发育情况,并拍照记录。
2 结果
2.1 延长光照时间对糜子生长发育的影响
短日照条件下,糜子植株营养器官的干物质积累量较低。与对照比较,糜子营养器官在长日照条件下能够积累更多的干物质,且长日照天数越长,糜子营养器官干物质积累量越多(表1)。延长光照25 d的糜子植株营养器官生物量和籽粒干重达到最高,增加幅度分别达到353.97%和490.91%;茎、叶、穗干物质积累分别比对照提高了416.67%、142.86%和412.00%,茎、穗、单穗籽粒的干物质积累量增幅高于叶片。但延长光照20 d各器官干物质重与延长光照25 d各器官干物质重不存在极显著差异(<0.01)。
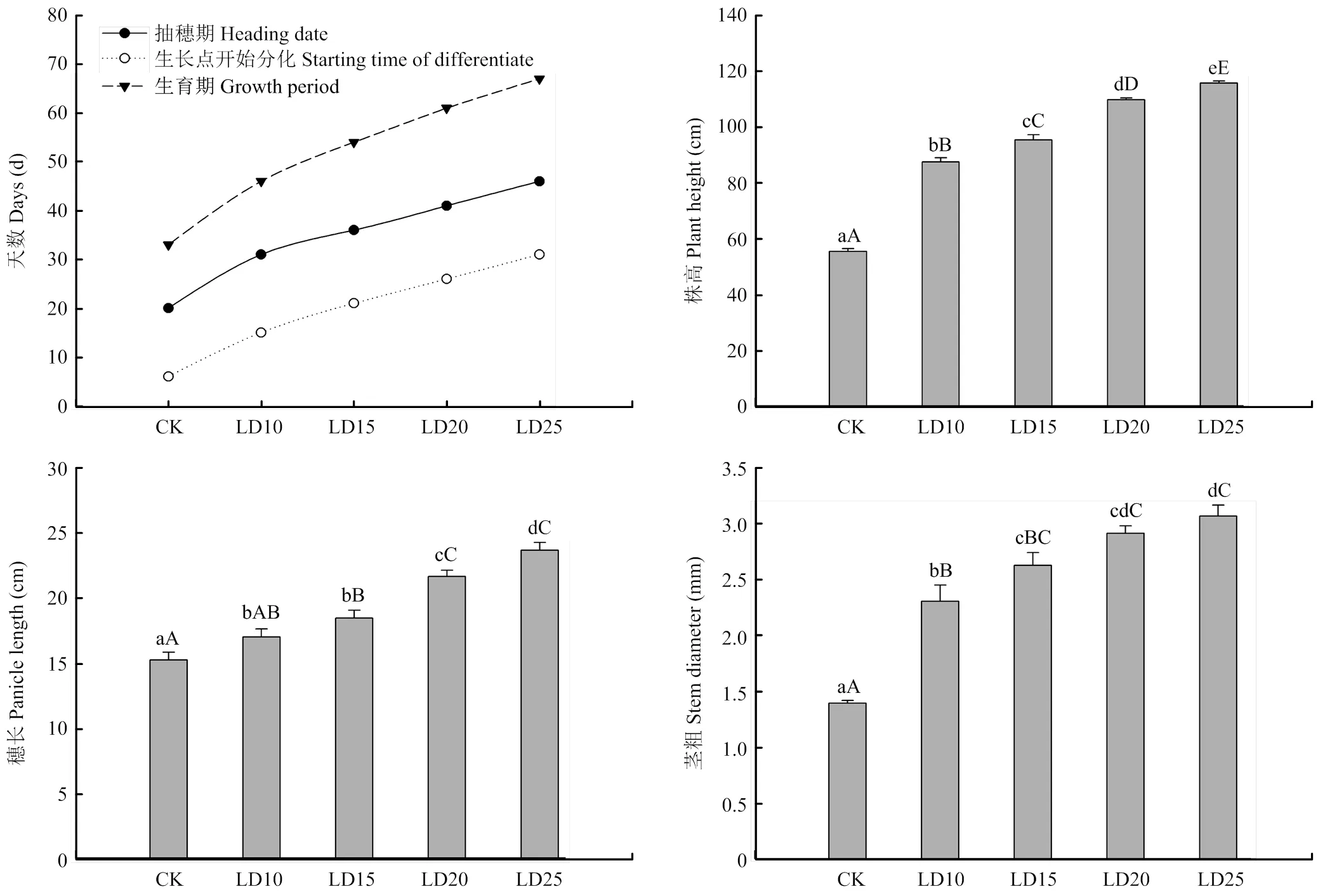
通过调查糜子营养生长期增加光照对糜子植株形态的影响(图1),发现株高、茎粗、穗长在长光照25 d处理后达到最大,茎粗和穗长与长光照20 d处理不存在显著差异(<0.01)。株高、茎粗的增幅较大,穗长的增幅较小。出苗后增加每天光照时间和增加长日照天数条件下,株高和茎粗增幅分别达到57.85%—108.98%和65.59%—120.38%,穗长的增幅达到11.64%—55.43%。分析出苗到抽穗的天数以及长日照天数之间的关系,发现短日照条件下5 d左右糜子即开始幼穗分化,20 d左右进入抽穗期。
2.2 光周期对糜子幼穗发育进程的影响
通过对所有参试的不同处理糜子幼穗发育观察(图2),在长日照条件下糜子一直处于营养生长期,茎尖生长点处于未伸长期;在短日照条件下5 d后茎尖生长点开始伸长,幼穗开始分化;分化开始后,不同处理条件下,糜子幼穗分化持续时间均为15 d左右开始抽穗。即在短日照条件下,糜子出苗(CK)或转移至短日照条件下20 d即开始抽穗,没有受到前期延长光照时间的影响。根据观察,糜子的幼穗分化过程可分为7个时期:(1)未伸长期。生长点处于未伸长期时,植株为营养生长状态。生长锥点在伸长之前为球体,呈光滑透明状,宽度与高度几乎相等,生长点被叶原基紧紧包住。长光照条件下,糜子幼穗保持此时期形态,不发生变化;(2)生长点伸长期(图2-A)。受到短日照条件诱导,生长锥开始伸长,此时营养生长与生殖生长并进。伸长的生长点高度明显大于宽度,生长点逐渐伸长为锥体为生长锥,同时在基部出现半球状突起,为一级枝梗分化;(3)穗枝梗原基分化期(图2-B和图2-C)。一级枝梗分化与生长锥伸长同时进行。生长锥基部长出乳头状突起,此为一级枝梗原基,一级枝梗原基逐渐伸长为锥体。一级枝梗原基分化的同时,生长锥中部即幼穗中部分化出苞叶原基,表现为“二棱期”。苞叶原基出现标志穗轴开始分化。随着分化的进行,苞叶原基被一级枝梗遮盖,只能看到一级枝梗。一级枝梗无序排列,呈向顶式生长。当一级枝梗发育到一定程度之后,二级枝梗原基出现在在一级枝梗锥体两侧,分别排列在锥体两侧。随后在二级枝梗的两侧分化出三、四、五级枝梗,分化方式与二级枝梗相同,都分别在其上一级枝梗锥体两侧生出并伸长;(4)小穗原基分化期(图2-D)。三级枝梗原基出现在穗中部时,早生枝梗的的一侧出现突起,随后在另一侧突起的外侧出现突起,两侧突起逐渐伸长将枝梗包围。此时分化的是小穗的护颖原基。同时,穗枝梗开始伸长。小穗原基分化从顶生生长点开始逐渐向下分化;(5)小花原基分化期(图2-E和图2-F)。糜子每个小穗上发育2朵小花,第一朵小花在发育过程中退化,第二朵小花发育完全。第一朵小花靠近外颖产生,外稃位于外颖一侧,內稃位于外稃对侧,内外稃之间有一个半球状的突起即为第一小花原基。第一朵小花在此及出上停止发育。第二朵小花于第一朵小花上位产生,且较第一朵小花大,由原生长点分化而来。第二朵小花的外稃位于内颖一侧,內稃在其对侧发育;(6)雌雄蕊原基分化期(图2-G和图2-H)。小花原基分化出內稃之后,第二朵小花原基体积增大,并在其上部分化出4个乳头状突起。中间为雌蕊原基,周边3个为雄蕊原基。随着发育的进行,雌蕊原基上方出现乳头状突起,即为胚珠原基。雄蕊原基上方出现一条横沟和一条纵沟将雄蕊原基分成4个部分,即花粉囊。花粉母细胞形成于花粉囊,花药内容物逐渐充实,花粉母细胞经过2次减数分裂形成四分体,四分体成熟后散开即为初生花粉粒。此过程伴随着花丝伸长,柱头分叉呈羽毛状,子房膨大,小穗基部膨大呈椭球状;(7)花粉粒形成期(图2-I)。进入花粉粒形成期的小穗,内外颖迅速生长,尖端呈现紫色,花丝伸长呈现橘黄色,雌蕊柱头呈现乳白色。
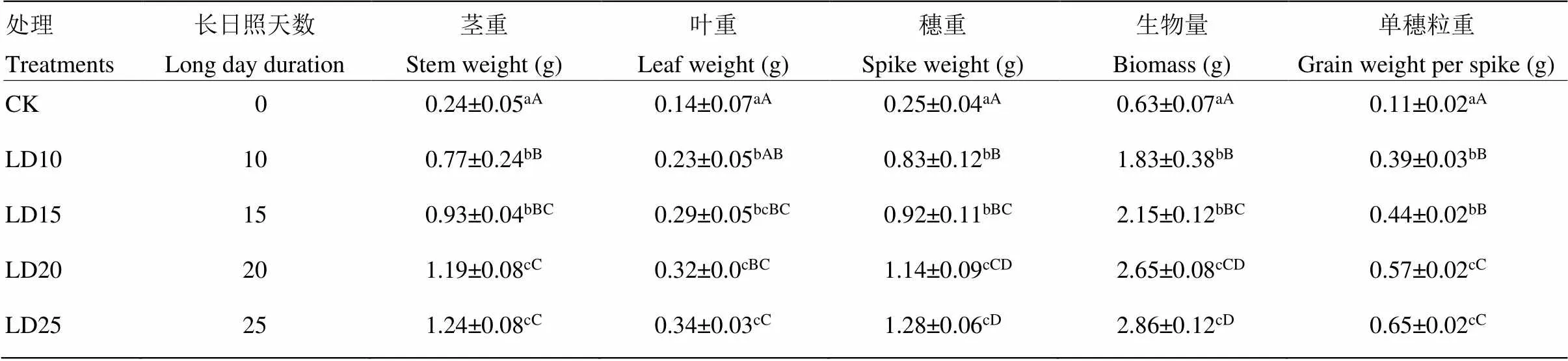
表1 延长光照时间对糜子生物产量的影响
同列数据后不同小写字母表示处理间差异达5%显著水平;同列数据后不同大写字母表示处理间差异达1%显著水平
Different lowercase letters after the same data column indicate a significant difference at a 5% level between different treatments; capital letters indicate a significant difference at a 1% level between different treatments
不同小写字母表示处理间差异达5%显著水平;不同大写字母表示处理间差异达1%显著水平

A:出苗后7 d,生长点伸长期;B:出苗后8 d,穗枝梗原基分化期,一级枝梗原基分化;C:出苗后9 d,穗枝梗原基分化期,二级枝梗原基分化;D:出苗后12 d,小穗原基分化期,出现护颖原基;E:出苗后13 d,小花原基分化期,第一朵小花分化;F:出苗后14 d,小花原基分化期,第二朵小花分化;G:出苗后17 d,雌雄蕊原基分化期,花粉母细胞形成;H:出苗后18 d,雌雄蕊原基分化期,花粉母细胞减数分裂;I:出苗后21 d,花粉粒形成期
3 讨论
光是植物调控其生长与发育的重要环境信号之一,植物通过整合外源信号与内源信号,继而调控准确的开花时间,确保植物在特定环境下的最佳时机开花以保证产量,因此,开花期成为了决定品种地区适应性和季节适应性的关键因素[18],对作物的产量形成有着十分重要的作用。延长光照时间能够增长植物光合作用时间,延迟植株开花时间,促进营养物质在植株体内的积累。本研究在延长光照时间后,抽穗期随着长日照时间的增加而推迟,抽穗期从出苗后21 d(CK)增加至出苗后的46 d。营养生长阶段随之延长,穗重、穗粒重极显著的提高。糜子出苗后在长日照环境中保持营养生长,营养生长的增加为糜子植株的正常生长提供了保障。比如在海南生长期间,增加糜子生长前期的光照长度可以为糜子提供较长营养生长时间,为育种、制种提供良好保障。
植物的开花机制比较复杂,目前,已知的6种不同的开花调控途径包括光周期途径、温度途径、春化途径、赤霉素途径、自主开花起始途径以及年龄途径[19],但这些不同开花调控途径之间的联系尚不明确。本研究是对糜子的光周期反应进行的初步探讨,通过对短日照诱导的叶片和茎尖生长点等关键器官进行不同时间点的转录组测序研究可初步了解一些光周期感应及信号传导等关键基因的表达模式(尚未发表,数据未列出),同时鉴定和筛选分别对长和短光周期反应迟钝的新种质,结合基因组学进行遗传分析,可为探讨糜子开花机制,培育广适应性的糜子新品种奠定基础。
在禾谷类粮食作物中,糜子既是最抗旱的作物之一,又能用最低的需水量生产其他作物相同的单位重量干物质,同时又具有很强的耐瘠性,可在干旱贫瘠的土壤中收获其他作物所达不到的产量糜子是需水最少的粮食作物,且生长周期较短,这些特性使得糜子成为抗逆生物学研究的最佳材料,引起了众多研究者的注意。近几年相继开展了糜子分子标记的开发[20-21]、糜子群体结构和遗传多样性分析[22-24]、不同基因型对盐碱和干旱胁迫的反应[25]以及通过转录组测序对糜子生长发育及抗逆等生理过程中的关键转录因子进行分析[26],2019年中国科学院上海植物逆境生物学研究中心[27]和中国农业大学[28]等相继完成了糜子全基因组测序工作,并通过比较基因组和转录组分析揭示糜子进化历程和其特殊的C4途径模型。糜子全基因组序列的公布,对进一步研究糜子对光周期变化反应的分子机理将具有极大的推动作用。
4 结论
糜子是典型的短日作物,在18 h长日照条件下糜子茎尖生长点处于未伸长期,持续营养生长,不进行生殖生长,短日照条件下生长5 d即开始幼穗花芽分化;在短日条件下出苗后延长20 d的长日照条件就可以极显著增加糜子株高、茎粗、穗长、穗重和穗粒重等。短日照条件能够诱导和促进糜子从营养生长到生殖生长的转变,而长日照条件则抑制糜子生殖生长的开始;糜子的幼穗分化可以划分为7个不同的时期,生殖生长开始后不受前期延长光照时间的影响,持续时间在15 d左右。
[1] 屈洋, 苏旺, 李翠, 高金锋, 高小丽, 王鹏科, 冯佰利, 柴岩. 陕北半干旱区沟垄覆膜集水模式下糜子边际效应及生理特性. 应用生态学报, 2014, 25(3): 776-782.
QU Y, SU W, LI C, GAO J F, GAO X L, WANG P K, FENG B L, CHAI Y. Marginal effect and physiological characteristics of millet under furrow and ridge mulching catchment mode in semi-arid area of northern Shaanxi., 2014, 25(3): 776-782. (in Chinese)
[2] LU H, ZHANG J, LIU K B, WU N, LI Y, ZHOU K, YE M, ZHANG T, ZHANG H, YANG X, SHEN L, XU D, LI Q. Earliest domestication of common millet (L.) in East Asia extended to 10,000 years ago., 2009, 106(18): 7367-7372.
[3] AMADOUBR I, LE M. Millets: Nutritional composition, some health benefits and processing-A Review., 2013, 25(7): 501.
[4] ANDERSON R. Planting date effect on no-till proso millet., 1994, 7(4): 454-458.
[5] BADAU M H, NKAMA I, JIDEANI I A. Phytic acid content and hydrochloric acid extractability of minerals in pearl millet as affected by germination time and cultivar., 2005, 92(3): 425-435.
[6] KALINOVA J, MOUDRY J. Content and quality of protein in proso millet (L.) varieties., 2006, 61(1): 43.
[7] 晁桂梅. 品种及栽培环境对糜子淀粉理化性质影响研究[D]. 杨凌: 西北农林科技大学, 2016.
CHAO G M. Effects of varieties and cultivation conditions on physicochemical properties of proso millet starches[D]. Yangling: Northwest A&F University, 2016. (in Chinese)
[8] 朱静娴. 人工补光对植物生长发育的影响. 作物研究, 2012, 26(1): 74-78.
ZHU J X. Influence of artificial supplement of light on plant growth and development., 2012, 26(1): 74-78. (in Chinese)
[9] 杜洪涛, 刘世琦, 张珍. 光质对彩色甜椒幼苗生长及酶活性影响. 华北农学报, 2005(2): 45-48.
DU H T, LIU S Q, ZHANG Z. Effects of light qualities on growth and activity of enzymes in leaves of color pimientos seedling., 2005(02): 45-48. (in Chinese)
[10] 龙作义, 刘汉平, 吴全德. 光周期对红皮云杉苗木的影响. 牡丹江师范学院学报, 1999(1): 12-13.
LONG Z Y, LIU H P, WU Q D. Effects of photoperiod on seedlings of., 1999(1): 12-13. (in Chinese)
[11] 廖祥儒, 张蕾, 徐景智, 王俊峰, 李同凯. 补充光照对番茄幼苗生长和结果的影响. 河北大学学报, 2003, 23(1): 55-58.
LIAO X R, ZHANG L, XU J Z, WANG J F, LI T K. Effects of supplementary illumination on growth and fruit of tomato seedlings., 2003, 23(1): 55-58. (in Chinese)
[12] 王惠珍, 喻敏, 萧洪东, 蔡凤玲, 邓文杰. 施硅对硅细胞的发育及不同光照时间处理海滨雀稗可溶性糖含量的影响. 华中农业大学学报, 2007(4): 482-485.
WANG H Z, YU M, XIAO H D, CAI F L, DENG W J. Influences of Si on Si cells formation and content of soluble sugars in Seashore Paspulum (Swarfz.)under different light duration., 2007(4): 482-485. (in Chinese)
[13] ODA A, NARUMI T, LI T, KANDO T, HIGUCHI Y, SUMITOMO K, FUKAI S, HISAMATSU T. CsFTL3, a chrysanthemum flowering locus T-like gene, is a key regulator of photoperiodic flowering in chrysanthemums., 2012, 63(3): 1461-1477.
[14] HIGUCHI Y, HISAMATSU T. CsTFL1, a constitutive local repressor of flowering, modulates floral initiation by antagonising florigen complex activity in chrysanthemum., 2015, 237: 1-7.
[15] 魏仰浩, 董进亚. 缩短光照对糜子生长发育的影响. 农业科学实验, 1982, 8: 46.
WEI Y H, DONG J Y. Effect of shortening light on the growth and development of proso millet, 1982, 8: 46. (in Chinese)
[16] 杨宗渠, 尹钧, 谷冬艳, 周冉, 任江萍, 李永春, 李金才. 不同发育特性小麦品种叶片与小穗原基分化同步关系的研究. 核农学报, 2007, 21(6): 550-556.
YANG Z Q, YIN J, GU D Y. ZHOU R, REN J P, LI Y C, LI J C. Corresponding relation between leaf spikelet primordium differentiation of different development type of wheat cultivars., 2007, 21(6):550-556. (in Chinese)
[17] 郜吉祥. 固原地区糜子幼穗分化的初步观察. 宁夏农业科技, 1984, 4: 4-6, 57.
GAO J X. Preliminary observation on young panicle differentiation of proso millet in Guyuan area., 1984, 4: 4-6, 57. (in Chinese)
[18] 贾小平, 李剑峰, 赵渊, 全建章, 董志平, 戴凌峰, 张小梅, 张博, 袁玺垒. 谷子抽穗期与农艺性状的相关与回归分析. 植物遗传资源学报, 2019, 20(3): 634-645.
JIA X P, LI J F, ZHAO Y, QUAN J Z, DONG Z P, DAI L F, ZHANG X M, ZHANG B, YUAN X L. Correlation and regression analysis between heading Date and agronomic traits in., 2019, 20(3): 634-645. (in Chinese)
[19] 李莉, 李旭, 刘亚文, 刘宏涛. 光和温度调控开花时间的研究进展. 中国科学: 生命科学, 2016, 46(3): 253-259.
LI L, LI X, LIU Y W, LIU H T. Research progress of flowering time regulation by light and temperature., 2016, 46(3): 253-259. (in Chinese)
[20] JIANG Y, LI H, ZHANG J, XIANG J, CHENG R, LIU G. Whole genomic EST-SSR development based on high-throughput transcript sequencing in proso millet ()., 2018, 20(3): 617-620.
[21] HOU S, SUN Z, LI Y, WANG Y, LING H, XING G, HAN Y, LI H. Transcriptomic analysis, genic SSR development, and genetic diversity of proso millet (Poaceae)., 2017, 5(7): 1600137.
[22] 王瑞云, 季煦, 陆平, 刘敏轩, 许月, 王纶, 王海岗, 乔治军. 利用荧光SSR分析中国糜子遗传多样性. 作物学报, 2017, 43(4): 530-548.
WANG R Y, JI X, LU P, LIU M X, XU Y, WANG L, WANG H G, QIAO Z J. Analysis of genetic diversity in common millet () using fluorescent SSR in China., 2017, 43(4): 530-548. (in Chinese)
[23] MARKO F, NATASA S, KOCJAN A D. Genetic diversity and agronomic performance of Slovenian landraces of proso millet (L.)., 2019, 43(2): 185-195.
[24] JOHNSON M, DESHPANDE S, VETRIVENTHAN M, UPADHYAYA H D, WALLACE J G. Genome-wide population structure analyses of three minor millets: kodo millet, little millet, and proso millet., 2018, 499087.
[25] CARUSO C, MAUCIERI C, BERRUTI A, BORIN M, BARBERA A. Responses of DifferentL. genotypes to saline and water stress in a marginal mediterranean environment., 2018, 8(1): 8.
[26] YUE H, WANG M, LIU S, DU X, SONG W, NIE X. Transcriptome- wide identification and expression profiles of the WRKY transcription factor family in broomcorn millet (L.)., 2016, 17(1): 343.
[27] SHI J, MA X, ZHANG J, ZHOU Y, LIU M, HUANG L, SUN S, ZHANG X, GAO X, ZHAN W, LI P, WANG L, LU P, ZHAO H, SONG W, LAI J. Chromosome conformation capture resolved near complete genome assembly of broomcorn millet., 2019, 10(1): 464.
[28] ZOU C, LI L, MIKI D, LI D, TANG Q, XIAO L, RAJPUT S, DENG P, PENG L, JIA W, HUANG R, ZHANG M, SUN Y, HU J, FU X, SCHNABLE P S, CHANG Y, LI F, ZHANG H, FENG B, ZHU X, LIU R, SCHNABLE J C, ZHU J K, ZHANG H. The genome of broomcorn millet, 2019, 10(1): 436.
Effects of Photoperiod Changes on Morphological Characters and Young Panicle Development in Proso Millet (L.)
DONG Ming1, JIANG YanMiao1, LI HaiQuan1, GENG LingLing1, LIU JianYe2, QIAO ZhiHong2, LIU GuoQing1
(1Institute of Millet Crops, Hebei Academy of Agriculture & Forestry Sciences/The Key Minor Cereal Crops Laboratory of Hebei Province, Shijiazhuang 050035;2Yuxian Agricultural and Rural Burea of Hebei Province, Yuxian 075700, Hebei)
【Objective】 Proso millet is a typical short-day crop that is extremely sensitive to photoperiod changes which limits the geographical adaptation of proso millet varieties. In the present study, the important agronomic traits have been characterized and young panicle development has been observed under both short-day and long-day conditions, which may help better understand reaction mechanism to photoperiod changes and support widely adaptable variety breeding in proso millet. 【Method】A landrace “Erzigan” was planted in pots under a 18h day light condition, then transferred to a 12h day light condition after 0 day (CK), 10, 15, 20 and 25 days of seedling emergency, each treatment was replicated three times. The main plant characters including heading time, plant height, stem diameter, panicle length, dry panicle weight and dry grain weight per panicle were measured and young panicle development was observed and photographed by using stereo microscope. 【Result】 Plants grown for 25 days under long-day conditions had a plant height of 115.6 cm and a single panicle grain weight of 0.647 g, which was 109.0% and 472.6% higher than the control, respectively. The dry matter accumulation of stem, leaves and ears increased by 416.7%, 142.9% and 412.0%, respectively, comparing with the control. However, the difference in dry matter weight of each organ between the 25d and 20d treatments was not significant. Under the condition of 18 hours day light, the growing point of the stem tip of the proso millet kept in the unstretched period, which meant the plants were at vegetative growth stage without reproductive growing. After 5 days growing under short-day light conditions, the plants began reproductive growth, and the young panicles started to differentiate. The process of young panicle development could be classified into 7 stages including unstretched stage, growing point elongation stage, ear branch differentiation stage, spikelet differentiation stage, floret differentiation stage, pistil and stamen differentiation stage, and pollen grain formation stage. The whole process of young panicle development could last about 15 days before heading, which was not affected by the prolonged light period treatments at early growing stage. 【Conclusion】Artificially prolonging the light period under short day conditions could extend vegetative growth period and was in favor of dry matter accumulation, which can significantly increase the biomass and yielding of proso millet. Short-day conditions promote the transformation of proso millet from vegetative growth to reproductive growth. The whole process of young panicle development could be classified into seven stages, which could last about 15 days before heading and was not affected by all the five treatments at early growing stage.
proso millet (L.); photoperiod response; morphological characters; young panicle development
2019-06-13;
2019-09-03
国家现代农业产业技术体系(CARS-06-13.5-A10)、河北省农业创新工程(2019-4-2-5)
董明,E-mail:dddongming@126.com。通信作者刘国庆,E-mail:guoqingliu@hotmail.com
(责任编辑 李莉)
