利用iTRAQ技术和转录组筛选芍药属远缘杂交不亲和基因
2020-04-13贺丹谢栋博张佼蕊何松林李朝梅郑云冰王政刘艺平栗燕逯久幸
贺丹,谢栋博,张佼蕊,何松林,李朝梅,郑云冰,王政,刘艺平,栗燕,逯久幸
利用iTRAQ技术和转录组筛选芍药属远缘杂交不亲和基因
贺丹1,2,谢栋博1,张佼蕊1,何松林1,2,李朝梅1,郑云冰1,王政1,刘艺平1,栗燕1,逯久幸1
(1河南农业大学林学院,郑州 450002;2河南科技学院园艺园林学院,河南新乡 453003)
【目的】远缘杂交育种是目前牡丹、芍药品种改良和育种的主要方法,而远缘杂交不亲和一直是制约其快速发展的主要因素。本研究从牡丹、芍药远缘杂交授粉后不亲和应答相关的柱头差异蛋白与转录组方面深入研究,揭示牡丹、芍药远缘杂交不亲和的分子机理,为杂交育种提供理论依据。【方法】以芍药‘粉玉奴’自交、芍药‘粉玉奴’与牡丹‘凤丹白’杂交为供试材料,在授粉后24 h采取柱头,分别进行同位素标记相对定量(iTRAQ)和转录组技术分析。对所获得的蛋白和转录组数据进行生物信息学分析,并对其中可能与远缘杂交不亲和相关的基因进行定量PCR验证。【结果】利用iTRAQ技术分析牡丹、芍药远缘杂交后柱头中蛋白质的表达差异,共鉴定到685个差异蛋白,富集到了188条通路,其中显著富集的Pathway有18条。与不亲和授粉相关代谢通路有RNA降解、钙信号途径、丝裂原活化蛋白激酶(mitogen-activated protein kinase signaling pathway,MAPK)信号途径、磷脂酰肌醇信号系统。在RNA降解代谢通路中,烯醇酶(Enolase)、热休克蛋白DnaK(HSP70)及病菌抗原(GroEL)均表达下调。在钙信号途径中,钙调蛋白(CALM)表达下调,腺苷酸转运酶(adenine nucleotide translocase,ANT)表达量增加,表达上调。MAPK信号途径中,乙二醛酶Ⅰ(GloI)表达下调。磷脂酰肌醇信号系统中的钙调蛋白(CALM)表达下调。随机选取与差异蛋白相关的6个基因进行qRT-PCR验证,结果显示,6个基因的表达与蛋白质水平趋势相一致,均表达下调。通过转录组测序,共获得了52 998个有注释信息的Unigene,占所有Unigene的40.37%。基于6组样品的RPKM(Reads Per Kilobase per Million)值,共筛选到16 224个差异基因。其中上调基因13 361,下调基因2 863个。对差异基因进行pathway显著富集分析,杂交与自交相比,不亲和差异表达的基因主要富集在氧化磷酸化代谢、ABC转运蛋白、次级代谢产物等通路。与远缘杂交不亲和相关且发生显著变化的基因有、(胼胝质酶)和(squamosapromoter binding protein-like)表达上调,(ABC transporter family protein)表达下调。【结论】在转录组和蛋白数据共注释到6个蛋白、4个基因与植物不亲和性密切相关,这些蛋白与基因可能在远缘杂交不亲和方面发挥着重要作用。
牡丹;芍药;iTRAQ;转录组;远缘杂交
0 引言
【研究意义】牡丹()是芍药科芍药属的落叶亚灌木,迄今已有1 600多年的栽培历史,是我国的传统名花,也是我国特有的名贵观赏兼药用植物[1]。芍药科芍药属的牡丹与芍药,被誉为“花王和花相”,具有很高的观赏和生产价值。牡丹、芍药的新品种培育工作一直是科研和生产的重要内容。杂交育种作为传统的育种方式,也是牡丹、芍药的主要育种方法。利用杂交育种能够培育出一些具有较高观赏价值、抗寒、抗病等特点的新品种,而芍药属远缘杂交在国内的研究还处于初步阶段[2-3]。通过一系列研究发现受精前障碍是牡丹、芍药远缘杂交育种过程中存在的严重问题。在杂交过程中,虽然少量花粉能在柱头上萌发并穿过柱头,但花粉管的伸长却受到阻碍,并且在柱头上产生了大量的胼胝质,从而阻碍花粉管进入子房完成受精[4-5]。通过研究牡丹、芍药远缘杂交不亲和性的机制,对克服远缘杂交受精前障碍,实现牡丹、芍药远缘杂交具有重要意义。【前人研究进展】利用转录组和蛋白质组学筛选相关基因能够提高育种效率,克服常规育种中的困难[6-9]。同位素标记相对和绝对定量(isobaric tags for relative and absolute quantitation,iTRAQ)技术是2004年美国应用生物系统公司(ABI)推出的一种新的蛋白质组学技术。其可靠的结果已经广泛应用在生命科学的多个领域,包括寻找功能蛋白,筛选生物标志物或特殊蛋白,研究抗逆机理等[10]。程云清等[11]以平欧杂交榛‘达维’的正常发育与败育子房为材料,进行蛋白样品技术分析,初步筛选获得可能参与调控榛子子房败育的候选蛋白37个。目前,有关iTRAQ技术应用于植物杂交不亲和性的研究鲜少报道。Chalivendra等[12]采用iTRAQ技术研究番茄的种间生殖障碍,通过对蛋白质组变化的分析发现花粉-柱头互作中的蛋白包括S-RNases、HT-A蛋白、细胞壁疏松以及抗性响应相关蛋白。LI等[13]使用iTRAQ技术揭示了大米花粉-柱头互作的蛋白机制,发现泛素化在授粉的信号转导过程中具有重要作用。利用转录组技术可以挖掘重要的功能基因,揭示优良性状的分子机制,还可以研究不同器官、不同环境胁迫下基因表达的差异。转录组测序技术已被广泛应用于蜡梅、油松、油桐等的研究中[14-16]。ZHOU等[17]对羊草成熟花柱、子房、叶片高通量测序,发现多达1 025个转录本在花柱中特异性表达,这些转录本集中在细胞间交流与信号传导。【本研究切入点】目前,中外学者探讨了一些植物育种中的不亲和机理,但关于牡丹、芍药远缘杂交不亲和机理的研究还很欠缺。本研究在前期芍药属远缘杂交的基础上,发现芍药自交与芍药属远缘杂交亲和性存在显著差异。【拟解决的关键问题】以芍药‘粉玉奴’自交、芍药‘粉玉奴’与牡丹‘凤丹白’杂交授粉后24 h的柱头为供试材料,分别进行转录组和iTRAQ分析及后续分析,以期从中筛选出与杂交不亲和性相关的基因及蛋白,揭示芍药属远缘杂交不亲和性分子机制,为芍药远缘杂交育种提供理论基础。
1 材料与方法
1.1 材料及处理
供试母本芍药品种‘粉玉奴’与父本牡丹品种‘凤丹白’均引自山东菏泽,于2011年栽植于河南农业大学苗圃基地,植株生长良好,可以正常开花结实。于2018年4月初采取‘凤丹白’花粉,进行生活力测定后,储存于4℃冰箱。4月中下旬进行杂交,自交组合为‘粉玉奴’ב粉玉奴’,杂交组合为‘粉玉奴’ב凤丹白’。依据前期荧光显微观察确定杂交障碍发生的关键时间,授粉后24 h取自交与杂交的柱头用液氮速冻并置于-80℃冰箱内储存备用。
1.2 iTRAQ标记
采用丙酮沉淀法提取自交与杂交组柱头的蛋白质。对提取后的蛋白样品进行还原烷基化处理,用考马斯亮兰法测定蛋白浓度,经SDS-PAGE检测后将样品取等量蛋白Trypsin酶解,用iTRAQ 113、114标记未经授粉的柱头中的蛋白,iTRAQ 115、116标记杂交授粉后柱头中的蛋白,TRAQ 117、118标记自交授粉后柱头中的蛋白,最后将标记好的样品分别等量混合,应用强阳离子交换色谱(strong cation ex- change chromatography,cx)方法对混合后的肽段进行预分离,随后再进行液相分离和质谱分析。
1.3 转录组分析
采用天根生物公司多糖多酚植物总RNA提取试剂盒(DP441)提取RNA,进行1%琼脂糖凝胶电泳检测,120 V电泳20 min。取1 μL RNA样品,采用Thermo公司生产的Nano Drop ND-5000型紫外可见分光光度计测定样品的A260/A230和A260/A280的比值及RNA浓度,进行RNA的浓度和质量检测。检测符合建库要求的RNA送至武汉华大进行文库构建,采用BGISEQ-500平台进行高通量测序。
1.4 数据分析
首先,计算各个重复样品相对应蛋白质定量值的中位数作为待比较样品的定量值,并据此计算待比较样品间蛋白的最终差异倍数。其次,利用待比较样品间所有重复样品每个蛋白质的定量值进行T-检验,计算出-value,并利用Benjamini-Hochberg多重假设检验的方法校正值,得到校正后的值-value。最后根据差异倍数和-value来筛选出差异蛋白,当差异倍数达到1.5倍及以上(即up regulate≥1.5和down regulate≤0.67),且经过显著性统计检验其-value值≤0.05时,将其作为显著差异蛋白。
使用DEGseq软件对样品间进行差异表达分析,条件为Fold Change≥2且Adjustedvalue≤0.001,将-value矫正为-value。为了提高DEGs的准确性,定义差异倍数为两倍以上并且-value≤0.001的基因,筛选为显著差异表达基因。
1.5 定量PCR验证
使用TRIzol法对各样品进行总RNA提取,用ReverTra Ace qPCR RT Kit反转录试剂盒(东洋纺生物科技有限公司)进行cDNA的合成。以cDNA为模板,选择作为内参基因[18],检测引物为F:5′-TGAGCACCAAAGAAGTGGACGAAC-3′和R:5′-CACACGCCTGAACATCTCCTGAA-3′。使用SYBR Premix Ex TaqTMkit试剂盒(宝日医生物技术有限公司)在ABI PRISM 7900HT Real-Time PCR System(美国应用生物系统中国公司)上进行qPCR检测,每个样品进行3次技术重复。20 μL反应体系包括10 μL SYBR®Premix,1 μL cDNA,正、反引物各取0.5 μL,ddH2O 8 μL。两步法进行扩增,反应程序:预变性95℃,60 s;95℃变性15 s,60℃退火30 s,共40个循环。反应结束后进行溶解曲线分析,按照2-△△CT法计算出基因的相对表达量。引物设计采用Primer 5,基因及检测引物等相关信息详见表1。
2 结果
2.1 iTRAQ分析得到的与杂交不亲和相关蛋白代谢通路
通过对杂交不亲和、自交亲和授粉后的芍药雌蕊柱头差异蛋白进行Pathway富集分析,将 685个差异蛋白富集到188条通路,其中显著富集的Pathway有18条。通过分析,鉴定到与不亲和授粉相关的代谢通路有RNA降解(RNA degradation)、钙信号途径(Calcium signaling pathway)、促分裂原活化蛋白激酶信号途径MAPK及磷脂酰肌醇信号系统(Phosphatidylinositol signaling system)4个通路。4个通路中注释到的差异蛋白共有28个(表2)。

表1 qRT-PCR验证所选基因及对应引物信息

表2 远缘杂交不亲和相关的的差异蛋白通路
2.2 与远缘杂交不亲和相关的差异蛋白
在28个差异蛋白中,与不亲和相关的差异蛋白有6个。在RNA降解代谢通路中,以亲和授粉的雌蕊柱头蛋白为对照,烯醇酶(Enolase)、热休克蛋白DnaK(HSP70)及病菌抗原(GroEL)均表达下调。在钙信号途径中,钙调蛋白(CALM)表达下调,钙信号途径受到影响。当植物处于逆境胁迫时腺苷酸转运酶(ANT)表达量增加,表达上调。磷酸肌醇信号途径通过参与细胞外钙调素的信号转导,从而调控花粉萌发和花粉管伸长的启动。磷脂酰肌醇信号系统中的钙调蛋白(CALM)表达下调。MAPK信号途径参与调控磷酸肌醇信号转导、Ca2+信号途径等,也参与了IAA/ABA的代谢信号途径。乙二醛酶Ⅰ(GloI)是一种能够提高植物抗性的蛋白,该途径中的乙二醛酶Ⅰ(GloI)表达下调(表3)。
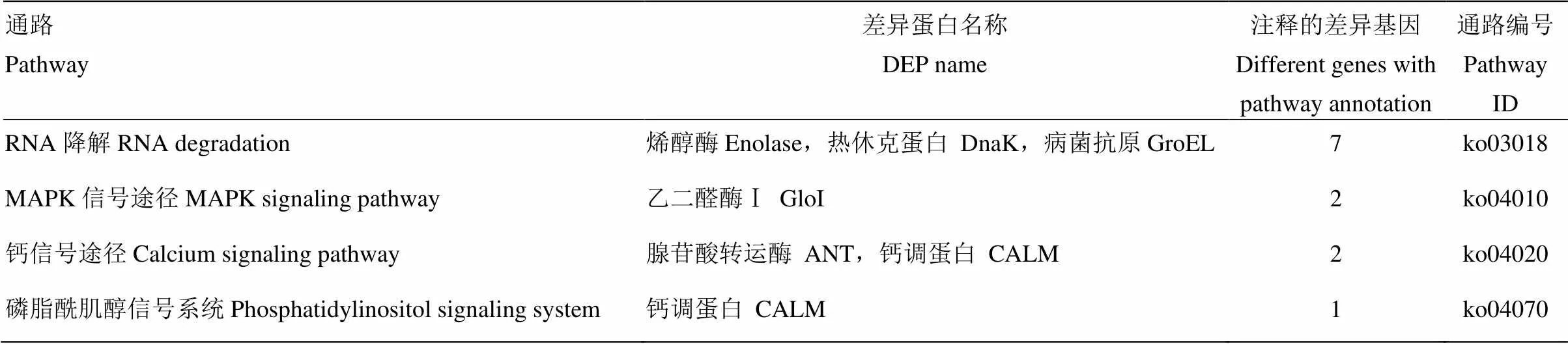
表3 与远缘杂交不亲和相关的差异蛋白
2.3 转录组分析得到的与授粉不亲和相关的基因
2.3.1 转录组数据的功能注释 为了了解杂交亲和性的基因表达情况,将测序的Unigene进行功能注释。通过选择BLAST参数E-value不大于1e-5和HMMER参数E-value不大于1e-10,与7个公共数据库比对后最终获得52 998个有注释信息的Unigene,占所有Unigene的40.37%。其中45 943条被注释到Nr数据库,35 181条被注释到NT数据库,33 186条被注释到Swissprot数据库,33 828条被注释到Pfam数据库,34 854条被注释到KEGG数据库,35 864条被注释到KOG数据库,被注释到GO数据库的有24 594条。
2.3.2 差异基因分析 基于6组样品的RPKM值,共筛选到16 224个差异基因。其中上调基因13 361,下调基因2 863个。通过KEGG Pathway显著性富集分析发现,这些差异表达的基因涉及多种生物学途径,将基因参与的KEGG代谢通路分为5大类:细胞过程(Cellular Processes)、环境信息处理(Environmental Information Processing)、遗传信息处理(Genetic Information Processing)、代谢(Metabolism)、有机系统(Organismal Systems)。杂交柱头与自交柱头相比,不亲和差异表达的基因主要富集在氧化磷酸化代谢、ABC转运蛋白、次级代谢产物等通路。
在KEGG分析中胼胝质酶、ABC转运蛋白、植物激素信号转导等途径是植物花粉发育,受精的重要途径[19-20],结合相关研究,从以上显著富集的差异表达基因集中筛选得到4个可能与杂交不亲和相关的基因(表4)。CL268_All、Unigene36776_All和CL7449_All在不亲和相对于亲和基因中显著表达上调。CL8637_All在不亲和相对于亲和基因中显著表达下调。

表4 不亲和相关的差异基因
2.3.3 相关差异基因的Blast比对分析 通过BLAST软件将CL268_All编码的氨基酸序列与其他物种对比筛选同源序列(图1),发现该基因与葡萄()(82.70%)、茶()(82.42%)、栓皮栎()(81.76%)、核桃()(81.21%)、桃()(80.87%)的相似度较高。
通过BLAST软件将Unigene36776_All编码的氨基酸序列与其他物种对比筛选同源序列(图2),发现该基因与茶()(81.56%)、葡萄()(80.69%)、樱桃()(80.53%)、桃()(80.75%)、核桃()(80.10%)的相似度较高。
通过BLAST软件将CL8637_All编码的氨基酸序列与其他物种对比筛选同源序列(图3),发现该基因与哥伦比亚锦葵()(82.66%)、橡胶树()(83.05%)、核桃()(82.76%)、栓皮栎()(83.84%)、枣()(82.95%)的相似度较高。

图1 CL268_All与其同源基因编码氨基酸序列的对比
通过BLAST软件将CL7449_All编码的氨基酸序列与其他物种对比筛选同源序列(图4),发现该基因与光皮桦()(73.74%)、榴莲()(76.78%)、麻风树()(73.74%)、核桃()(74.78%)、胡杨()(73.54%)的相似度较高。
2.4 差异表达蛋白相关基因的qRT-PCR验证
为了验证差异蛋白结果,随机选取gi|327424468_1(烯醇酶)、gi|327436891_2(热休克蛋白)、gi|327427673_4(热休克蛋白)、gi|327426876_5(病菌抗原)、gi|327427197_1(钙调蛋白)、gi|327433480_5(乙二醛酶Ⅰ)等差异蛋白对应的基因进行qRT-PCR验证,结果如图5。6个差异蛋白所对应基因在自交与杂交时期芍药雌蕊柱头中表达量和变化趋势有显著差异,表明这些基因在牡丹、芍药杂交过程中的表达具有明显的空间特异性。定量PCR结果与蛋白质水平趋势相一致,均呈现下调表达。
3 讨论
远缘杂交后芍药柱头中的差异蛋白及差异基因参与较多的通路中也包括RNA降解。在RNA降解代谢通路中,以亲和授粉的雌蕊柱头为对照,烯醇酶、热休克蛋白、病菌抗原均下调表达,其中烯醇酶参与了花粉-柱头相互作用。SHEORAN等[21]在油菜的研究中发现烯醇酶参与了花粉-柱头相互作用,亲和授粉后,烯醇酶在萌发的花粉中上调表达。陶璐与岳训[22]在拟南芥花粉管与柱头的互作研究中发现花粉管的延长触发了柱头组织中的糖酵解途径,激活了糖类物质转化为丙酮酸盐,而丙酮酸盐进一步代谢为被花粉管吸收的营养物质。在丙酮酸盐代谢的过程中,烯醇酶作为关键性的酶参与了丙酮酸盐的转化。因此,烯醇酶与花粉生长及授粉的亲和性密切相关。热休克蛋白是防御类蛋白质,与其他分子物质合作,稳定先前存在的蛋白质以抵抗聚集并介导细胞溶质中以及细胞器内新翻译的多肽折叠,结合延伸的肽片段,具有损伤后应激诱导的作用[23]。花粉萌发穿过柱头可以看作是一种外来物质入侵植物的行为,会引发植物的防御系统,促进防御类蛋白的产生或者上调表达。而在本研究中,热休克蛋白下调表达,其原因有待进一步研究。

图3 CL8637_All与其同源基因编码氨基酸序列的对比

图4 CL7449_All与其同源基因编码氨基酸序列的对比
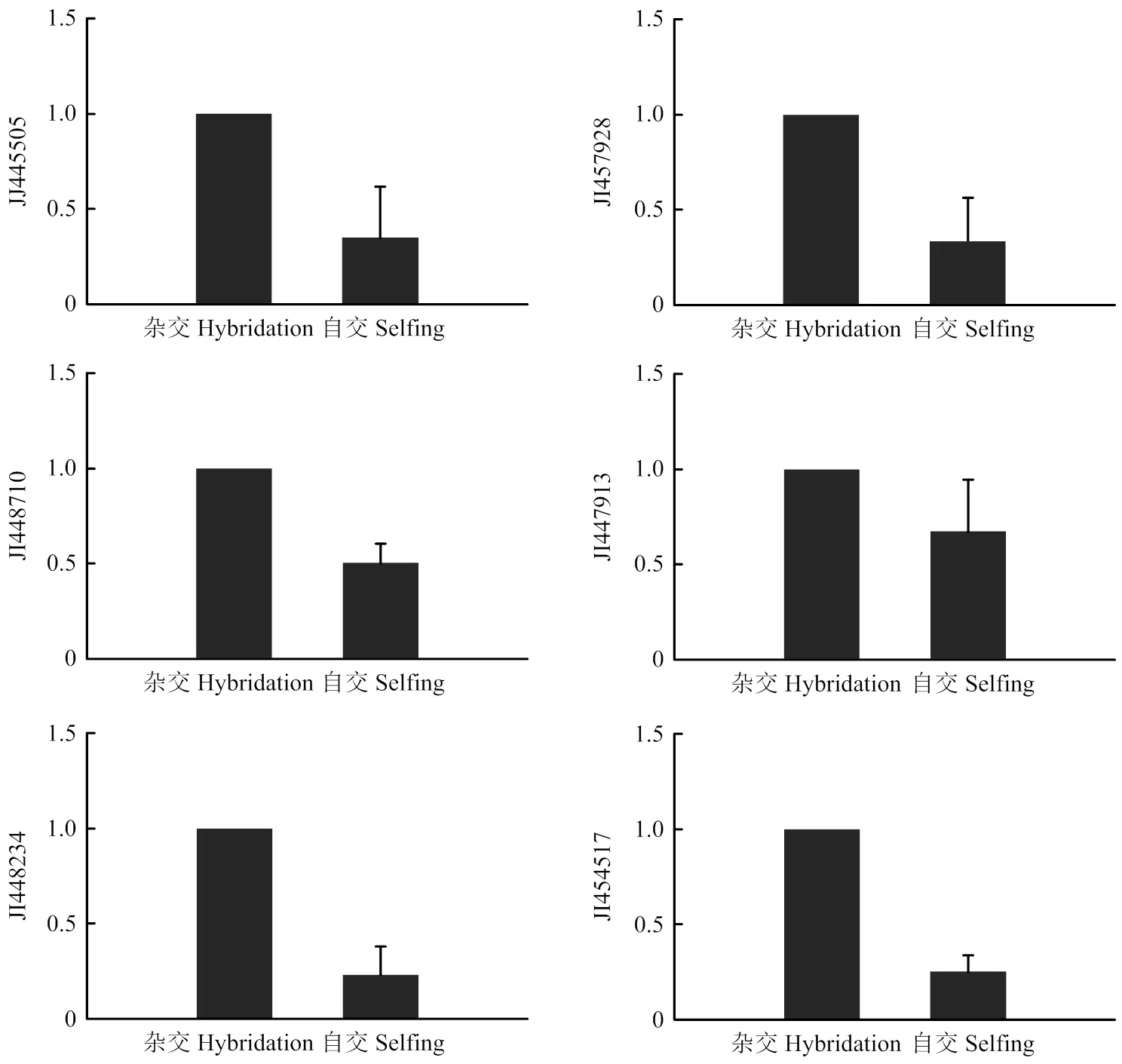
图5 JI445505、JI457928、JI448710、JI447913、JI448234和JI454517等6个基因的qRT-PCR验证
花粉管极性生长受多种信号和代谢途径调控,包括MAPK信号途径、磷脂酰肌醇信号通路等[24]。磷脂酰肌醇信号系统参与花粉萌发和花粉管伸长,磷酸肌醇途径受到抑制之后,会扰乱花粉管顶端Ca2+浓度梯度的产生和维持,还导致胼胝质在顶端的大量沉积[25]。本研究中,牡丹、芍药不亲和授粉后磷脂酰肌醇信号途径中的钙调蛋白下调表达,磷脂酰肌醇途径受到了抑制,从而花粉萌发和花粉管伸长受到了限制。MAPK信号途径中蛋白激酶活性降低抑制花粉管生长,反之则促进[26]。LI等[27]将不亲和的花粉用MAPK抑制剂处理后,花粉恢复活性,验证了MAPK介导花粉失活的假设。乙二醛酶系统存在于植物的叶、花等部位[28],甲基乙二醛(MG)是高等植物在逆境胁迫下产生的一种细胞毒素代谢物,而乙二醛酶Ⅰ(GloI)主要维持甲基乙二醛(MG)的动态平衡,清除过多的MG[28-29]。本研究中,MAPK信号途径中的乙二醛酶Ⅰ(GloI)下调表达,可能导致MG过多,从而引起花粉的死亡。
腺苷酸转运酶(adenine nucleotide translocator,ANT)作为ADP/ATP载体,催化ADP和ATP在线粒体内膜上的交换,为生物机体提供能量[30]。当植物处于逆境胁迫时,ANT表达量减少,在本研究中腺苷酸转运酶ANT表达下调可能是对花粉管穿入柱头的抗逆表达。胼胝质的合成与降解是一个严密的调控过程,受GSL酶的直接调控,与在转录组数据中涉及胼胝质酶合成通路,参与葡聚糖催化合成路径,并作为细胞膜的组成部分。其中在减数分裂后期发挥调控作用,不亲和植株中由其催化合成的胼胝质大量增多,最终导致植株育性降低;则主要参与胞间分子运动和抗逆反应[19]。
目前研究中发现S-Rnase介导自交不亲和,S-Rnase由花柱进入花粉管产生细胞毒性阻碍花粉管生长。自我S-Rnase能降解花粉管内RNA,导致蛋白质合成受阻,引起花粉管发生细胞程序性死亡[31-32]。在苹果中花柱S-Rnase与花粉MdABCF转运蛋白的跨膜区Tran结合,以内吞的方式进入膜内,运输至花粉管液泡中,积累到一定量后,被释放进入花粉管胞质引起花粉管生长停滞,最终引起自交不亲和反应[33]。MdABCF能够非选择性的运送S-Rnase至花粉管内。在植物大、小孢子发生以及雌、雄配子体发育过程中发挥重要作用。主要是与花粉囊发育及花药开裂相关[20]。本研究中上调表达,而花粉囊发育及花药开裂在本研究中均不存在问题,推测可能与花粉的进一步发育有关。
4 结论
本研究阐述了牡丹与芍药杂交不亲和性的分子机制,通过iTRAQ分析发现与远缘杂交不亲和性相关的代谢通路主要集中在RNA降解、MAPK信号途径、钙信号途径、磷脂酰肌醇信号系统4个途径。转录组的差异基因则主要是胼胝质合成酶(家族)、及相关基因。
[1] 文书生, 何绒绒, 郑家康, 田如男. 牡丹组织培养技术研究进展. 林业科学, 2018, 54(10): 143-155.
WEN S S, HE R R, ZHENG J K, TIAN R N. Research advances in tissue culture of tree peony., 2018, 54(10): 143-155. (in Chinese)
[2] 景士西. 园艺植物育种学总论. 北京: 中国农业出版社, 2000: 144-173.
JING S X..Beijing: China Agriculture Press, 2000: 144-173. (in Chinese)
[3] 贺丹, 解梦珺, 吕博雅, 王政, 刘艺平, 何松林. 牡丹与芍药的授粉亲和性表现及其生理机制分析. 西北农林科技大学学报(自然科学版), 2017, 46(10): 129-1360.
HE D, XIE M J, LV B Y, WANG Z, LIU Y P, HE S L. Analysis of pollination affinity performance and its physiological mechanism inand., 2017, 46(10): 129-136. (in Chinese)
[4] 何桂梅, 成仿云. 牡丹的杂交育种及其最新进展//中国观赏园艺研究进展. 北京: 中国林业出版社, 2004: 149-155.
HE G M, CHENG F Y. Crossbreeding ofand its latest progress//Beijing: China Forestry Press, 2004: 149-155. (in Chinese)
[5] 贺丹, 王雪玲, 高小峰, 吕博雅, 刘艺平, 何松林. 牡丹芍药远缘杂交亲和性. 东北林业大学学报, 2014, 42(7): 65-68.
HE D, WANG X L, GAO X F, LV B Y, LIU Y P, HE S L. Intergeneric cross-compatibility between peonies., 2014, 42(7): 65-68. (in Chinese)
[6] 张振乾, 肖钢, 春云, 邬贤梦, 熊兴华, 李云昌, 胡琼, 陈社员.利用转录组及iTRAQ技术筛选高油酸油菜抗病相关基因. 华北农学报, 2015, 30(5): 16-24.
ZHANG Z Q, XIAO G, CHUN Y, WU X M, XIONG X H, LI Y C, HU Q, CHEN S Y. The study of high oleic acid rapeseed disease resistance related genes by transcriptome and iTRAQ analysis., 2015, 30(5): 16-24. (in Chinese)
[7] 汪宝卿, 解备涛, 张海燕, 董顺旭, 段文学, 王庆美, 张立明. 基于iTRAQ技术的不同耐旱性甘薯苗期根系差异蛋白分析. 核农学报, 2017, 31(10): 1904-1912.
WANG B Q, XIE B T, ZHANG H Y, DONG S X, DUAN W X, WANG Q M, ZHANG L M. Analysis of differential proteome in roots during seedling stage of sweetpotato with different drought tolerance based on iTRAQ method., 2017, 31(10): 1904-1912. (in Chinese)
[8] 于涛, 李耕, 刘鹏, 董树亭, 张吉旺, 赵斌. 蛋白质组学分析揭示玉米籽粒发育过程中胁迫相关蛋白的表达特性. 中国农业科学, 2017, 50(11): 2114-2128.
YU T, LI G, LIU P, DONG S T, ZHANG J W, ZHAO B. Proteomic analysis of maize reveals expression characteristics of stress-related proteins during grain development., 2017, 50(11): 2114-2128. (in Chinese)
[9] 赵振利, 童琳琳, 邓敏捷, 董焱鹏, 范国强. 泡桐丛枝病发生过程中的蛋白质组学研究. 西南林业大学学报, 2018, 38(4): 23-28.
ZHAO Z L, TONG L L, DENG M J, DONG Y P, FAN G Q. Proteomic study on the occurrence ofwitches broom disease., 2018, 38(4): 23-28. (in Chinese)
[10] 谢秀枝, 王欣, 刘丽华, 董世雷, 皮雄娥, 刘伟. iTRAQ技术及其在蛋白质组学中的应用. 中国生物化学与分子生物学报, 2011, 20(7): 616-621.
XIE X Z, WANG X, LIU L H, DONG S L, PI X E, LIU W. iTRAQ technology and its application in proteomics., 2011, 20(7): 616-621. (in Chinese)
[11] 程云清, 齐名, 赵永斌, 邢继洋, 刘剑锋. 榛子正常发育与败育子房差异蛋白谱对比分析. 北京林业大学学报, 2018, 40(3): 13-25.
CHENG Y Q, QI M, ZHAO Y B, XING J Y, LIU J F. Comparative studies on differently expressed proteomes of developing and abortive ovary in hazelnut., 2018, 40(3): 13-25. (in Chinese)
[12] CHALIVENDRA S C, LOPEZ-CASADO G, KUMAR A, KASSENBROCK A R, ROYER S, TOVAR-MÈNDEZ A, COVEY P A, DEMPSEY L A, RANDLE A M, STACK S M, ROSE J K C, MCCLURE B, BEDINGER P A. Developmental onset of reproductive barriers and associated proteome changes in stigma/styles of., 2013, 64(1): 265-279.
[13] LI M, WANG K, LI S Q, YANG P F. Exploration of rice pistil responses during early post-pollination through a combined proteomic and transcriptomic analysis., 2016, 131: 214-226.
[14] 杨楠, 赵凯歌, 陈龙清. 蜡梅花转录组数据分析及次生代谢产物合成途径研究. 北京林业大学学报, 2012, 34(1): 104-107.
YANG N, ZHAO K G, CHEN L Q. Deep sequencing-based transcriptome profiling analysis ofreveals insights into secondary metabolites biosynthesis., 2012, 34(1): 104-107. (in Chinese)
[15] 钮世辉, 袁虎威, 陈晓阳, 李伟. 油松雌雄球花高通量基因表达谱芯片分析. 林业科学, 2013, 49(9): 46-51.
NIU S H, YUAN H W, CHEN X Y, LI W. Microarray analysis of large scale gene expression profiles between male and female cones of., 2013, 49(9): 46-51. (in Chinese)
[16] 孙颖, 谭晓风, 罗敏, 李建安. 油桐花芽2个不同发育时期转录组分析. 林业科学, 2014, 50(5): 70-74.
SUN Y, TAN X F, LUO M, LI J A. The sequencing analysis of transcriptome offlower buds at two development stages., 2014, 50(5): 70-74. (in Chinese)
[17] ZHOU Q Y, JIA J T, HUANG X, YAN X Q, CHENG L Q, CHEN Y, LI X X, NG A J, LIU G S. The large-scale investigation of gene expression instigmas provides a valuable resource for understanding the mechanisms of poaceae self-incompatibility., 2014, 15(1): 399.
[18] 贺丹, 李睿, 纪思羽, 吴静, 王政, 刘艺平, 何松林. 牡丹不定根形成相关基因PsARRO-1的克隆及表达分析. 植物生理学报, 2014, 50(8):1151-1158.
HE D, LI R, JI S Y, WU J, WANG Z, LIU Y P, HE S L. Cloning and expression analysis of adventitious rooting related geneof tree peony., 2014, 50(8): 1151-1158. (in Chinese)
[19] 崔海芳, 张凡, 尹俊龙, 郭瑛琪, 岳艳玲. 胼胝质沉积与花粉发育. 云南农业大学学报(自然科学), 2017, 32(3): 551-557.
CUI H F, ZHANG F,YIN J L,GUO Y Q,YUE Y L. Callose deposition and pollen development., 2017, 32(3): 551-557. (in Chinese)
[20] 李明, 李长生, 赵传志, 李爱芹, 王兴军. 植物SPL转录因子研究进展. 植物学报, 2013, 48(1): 107-116.
LI M, LI C S, ZHAO C Z, LI A Q, WANG X J. Research advances in plant SPL transcription factors., 2013, 48(1): 107-116. (in Chinese)
[21] SHEORAN I S, PEDERSEN E J, ROSS A R S, SAWHNEY V K. Dynamics of protein expression during pollen germination in canola ()., 2009, 230(4): 779-793.
[22] 陶璐, 岳训. 拟南芥花粉管与柱头互作的乙醇代谢耦合模型. 生物信息学, 2015, 13(1): 47-53.
TAO L, YUE X. Ethanol metabolism coupling model betweenpollen tube and stigma., 2015, 13(1): 47-53. (in Chinese)
[23] 于晓英, 卢向阳, 陈永华, 李达, 姚觉, 张宏志. 瓜叶菊热胁迫下的基因差异表达.农业生物技术学报, 2007, 15(3): 459-463.
YU X Y, LU X Y, CHEN Y H, LI D, YAO J, ZHANG H Z.Differential gene expression of×under heat stress., 2007, 15(3): 459-463. (in Chinese)
[24] QIN Y, YANG Z B. Rapid tip growth: insights from pollen tubes., 2011, 22(8): 816-824.
[25] 陈坤明. 磷酸肌醇信号途径对青扦花粉萌发和花粉管发育的调控作用及芦苇叶细胞壁结构的生理生态学研究[D]. 北京: 中国科学院, 2006.
CHEN K M. The mechanism of phosphoinositide signaling pathway involved in pollen germination and pollen tube growth ofand the ecophysiology of cell wall in the reed leaves [D]. Beijing: Chinese Academy of Sciences, 2006. (in Chinese)
[26] SALEM T, MAZZELLA A , BARBERINI M L, WENGIER D, MOTILLO V, PARISI G, MUSCHIETTI J. Mutations in two putative phosphorylation motifs in the tomato pollen receptor kinaseshow antagonistic effects on pollen tube length., 2011, 286(6): 4882-4891.
[27] LI S, SAMAJ J, FRANKLIN-TONG V E. A mitogen-activated protein kinase signals to programmed cell death induced by self-incompatibility inpollen., 2007, 145(1): 236-245.
[28] 叶芯妤, 邱雪梅, 王月, 李忠光. 乙二醛酶系统及其在植物响应和适应环境胁迫中的作用. 植物生理学报, 2019, 55(4): 401-410.
YE X Y, QIU X M, WANG Y, LI Z G. Glyoxalase system and its role in response and adaptation of plants to environmental stress., 2019, 55(4): 401-410. (in Chinese)
[29] SINGLA-PAREEK S L, REDDY M K, SOPORY S K. Genetic engineering of the glyoxalase pathway in tobacco leads to enhanced salinity tolerance., 2003, 100(25): 14672-14677.
[30] HAFERKAMP I, HACKSTEIN J H P, VONCKEN F G J, SCHMIT G, TJADEN J. Functional integration of mitochondrial and hydrogenosomal ADP/ATP carriers in themembrane reveals different biochemical characteristics for plants, mammals and anaerobic chytrids., 2010, 269(13): 3172-3181.
[31] 杜玉虎, 张绍铃. 温度对果梅离体花柱S-RNase识别特异性的影响. 中国农业科学, 2008, 41(9): 2734-2740.
DU Y H, ZHANG S L. Effects of temperature on recognition specificity of stylar S-RNase in vitro in., 2008, 41(9): 2734-2740. (in Chinese)
[32] 吴巨友, 李启明, 王鹏, 张绍铃. 梨自交不亲和性反应S-RNase新靶点—微丝骨架. 南京农业大学学报, 2018, 41(5): 7-9.
WU J Y, LI Q M, WANG P, ZHANG S L. Actin cytoskeleton is a new target of S-RNase in self-incompatibility of pear., 2018, 41(5): 7-9. (in Chinese)
[33] 孟冬. 苹果MdABCF转运S-RNase至花粉管影响自交不亲和反应[D]. 北京: 中国农业大学, 2014.
MENG D. Apple MdABCF transport S-RNase into pollen tube effecting self-incompatibility [D]. Beijing: China Agriculture University, 2014. (in Chinese)
Selected Related Genes about Incompatibility of Distant Hybridizationby iTRAQ Analysis and Transcriptome
HE Dan1,2, XIE DongBo1, ZHANG JiaoRui1, HE SongLin1,2, LI ChaoMei1, ZHENG YunBing1, WANG Zheng1, LIU YiPing1, LI Yan1, LU JiuXing1
(1College of Forestry, Henan Agricultural University, Zhengzhou 450002;2Colleg of Horticulture Landscape Architecture, Henan Institute of Science and Technology, Xinxiang 453000, Henan)
【Objective】Distant hybrid breeding is the main method of cultivar improvement and breeding in tree peony and herbaceous peony, while cross-incompatibility is an important restriction for breeding rapid development. Based on the previous researches, the analysis on different protein of stigma in pollen-stigma interaction and transcriptome was further explored. The mechanism of cross-incompatibility between tree peony and herbaceous peony was revealed, so as to provide the theoretical support for hybridized breeding.【Method】The stigmas of combinations‘Fenyunu’ בFenyunu’ and‘Fenyunu’ בFengdanbai’ were harvested at 24 h after pollination, which were used as materials for isobaric tags and analysis for relative and absolute quantitation (iTRAQ) and transcriptome, respectively. Bioinformatics was analyzed on the data of protein and transcriptome. Quantitative Real-time PCR (qRT-PCR) technique was used to validate the expression data of selected differentially expressed genes (DEGs). 【Result】iTRAQ was used to analyze DEPs of stigma of distant hybrid between tree peony and herbaceous peony, and the result showed that 685 DEPs were belonged to 188 pathways, in which 18 pathways were significantly enriched. There were four pathways with obvious difference in protein, including RNA degradation, mitogen-activated protein kinase (MAPK) signaling pathway, calcium signaling pathway, and phosphatidylinositol signaling system. In RNA degradation pathway, enolase, DnaK (HSP70), and GroEL (HSP60) were all down-regulated. In calcium signaling pathway, calmodulin (CALM) was down-regulated, while adenine nucleotide translocase (ANT) was up-regulated. In MAPK signaling pathway, Glyoxalase (GloI) was down-regulated. In phosphatidylinositol signaling system, CALM was also down-regulated. 6 genes were selected randomly to confirmed their expression by qRT- PCR, and the result showed that the expression profiles of the selected genes was in agreement with the results from protein analysis, and they were all down-regulated. A total of 52 998 annotated Unigenes were obtained by transcriptome sequencing, accounting for 40.37% of all Unigenes. Based on the RPKM (Reads Per Kilobase per Million) of six samples, 16 224 DEGs were obtained, among which13 61 were up-regulated, and 2 863 were down-regulated. Based on pathway enrichment analysis of DEGs, it indicated that the level of enrichment of DEGs in “Oxidative phosphorylation”, “ABC transporters” and “Biosynthesis of secondary metabolites” pathways were more significant and reliable than that of selfing. The genes related with incompatibility of distant hybridization were(Callose enzyme), and, which were up regulating expression, butthat was down regulating expression. 【Conclusion】In the data of transcriptome and protein, 6 proteins and 4 genes were closely related to incompatibility of distant hybridization. These proteins and genes might play an important role in incompatibility of distant hybridization.
;; iTRAQ; transcriptome; distant hybridization
2019-08-12;
2019-10-10
国家自然科学基金(31600568,31870698)、河南农业大学科技创新基金(KJCX2015A03,KJCX2018A05)、河南省科技攻关项目(192102110062,202102110234)、河南省高等学校重点科研项目(19A220003)
贺丹,E-mail:dandan990111@163.com。通信作者何松林,E-mail:hsl213@yeah.net
(责任编辑 赵伶俐)
