水稻直立短穗突变体esp的转录组研究
2020-04-13周坤能夏加发云鹏王元垒马廷臣张彩娟李泽福
周坤能,夏加发,云鹏,王元垒,马廷臣,张彩娟,李泽福
水稻直立短穗突变体的转录组研究
周坤能,夏加发,云鹏,王元垒,马廷臣,张彩娟,李泽福
(安徽省农业科学院水稻研究所/安徽省水稻遗传育种重点实验室,合肥 230001)
【目的】克隆水稻直立短穗基因(),分析其参与的基因调控途径,解析控制株型、穗长等农艺性状的分子机理。【方法】以直立短穗突变体及其野生型为材料,成熟期进行株高、穗长、粒长等表型测定;构建籼粳杂交F2定位群体,挑选与突变表型一致的F2单株,利用与突变性状连锁的分子标记对目的基因进行定位;对野生型和突变体进行基因组测序,结合定位结果,找到突变位点,克隆;利用生物信息学软件进行进化树和基因表达分析;提取野生型和突变体幼穗中的RNA并建库,GO(gene ontology)聚类分析表达差异基因,同时根据KEGG(kyoto encyclopedia of genes and genomes)数据库,分析野生型和突变体中植物激素信号转导和内质网蛋白加工相关基因的表达变化,并通过qRT-PCR验证。【结果】通过表型观察和农艺性状调查,与野生型相比,直立短穗突变体株高降低,穗长变短,穗型由弯曲变为直立,每穗粒数减少,粒长变短,粒宽和千粒重增加;有效穗数无显著差异。利用突变体与PA64构建籼粳F2定位群体,将目的基因定位于水稻第7染色体长臂标记C7-11和C7-14之间7.58 Mb区间内,基因组测序发现第6内含子与第7外显子连接位点由碱基G变异为A,导致第6内含子不能被剪切,蛋白翻译提前终止;该基因与已报道的/为等位基因。进化分析显示该基因广泛存在于单子叶和双子叶植物中;表达分析表明在茎秆、花序、雌蕊、内外稃和子房中高度表达,其表达水平随着子房变大而逐渐降低。利用转录组分析突变体和野生型幼穗中的基因表达,结果表明,与野生型相比,突变体中表达差异显著(差异>1.5倍)的基因630个,其中235个表达上调,395个表达下调。GO分析显示植物激素信号转导和内质网蛋白加工相关基因受到不同程度地调控,利用qRT-PCR进行验证,结果与转录组数据一致。【结论】直立短穗基因与已报道的直立穗基因/为等位基因,其突变导致株高降低、穗长变短等多个表型;可能通过调节植物激素信号转导、内质网蛋白加工过程中的基因表达,进而影响植株的发育。
水稻;直立短穗突变体;基因克隆;进化分析;转录组分析
0 引言
【研究意义】水稻是最重要的粮食作物之一,供世界上近一半的人口食用。利用株型筛选提高水稻产量是育种的一个重要方向。株型直立不仅能够提高光合效率,而且能够改善温度、湿度、CO2浓度、通风等田间条件,有助于提高植株生长速率和产量。直立穗品种穗型直立、株高较矮,能够提高植株抗倒性和耐肥性。【前人研究进展】目前,水稻中大量控制株型结构的基因被克隆,这些基因通过控制分蘖角度、分蘖数、穗型等调控株型,进而影响水稻产量[1-5]。然而,与直立穗性状有关的基因报道较少。通过QTL定位发现一个控制直立穗性状的基因,该基因的等位变异能够提高分生组织的活力,减少花序节间长度,增加二次枝梗和二次枝梗粒数,进而增加每穗粒数,提高水稻产量[6-8];与氮高效基因处于相同位点,具有耐肥特性,高氮条件下抗倒能力增强[9];研究表明在增加产量的同时对品质没有显著影响[10];此外,等位基因与和OsMADS1聚合可以同时提高水稻产量和米质[11]。通过该基因的等位变异选育的直立穗型水稻品种在生产上大面积推广应用[12-13]。水稻直立穗基因/编码一个植物特异的蛋白,该基因在幼嫩组织特别是幼穗中高度表达,/突变抑制初级和次级枝梗生长,降低穗长,穗型直立,但不影响植株产量[14-15]。小圆粒基因与/为等位基因,系列突变体中株高降低,粒型变小,穗型表现为直立或半直立表型[16]。突变体由中华11组培后代中获得,该突变体表现为穗部直立,株高、千粒重和穗数均显著降低,将其精细定位于水稻第7染色体,突变体存在大片段的DNA插入,该位点可能与/等位[17]。突变导致穗型直立,穗长变短,每穗粒数增加,显著提高植株产量[18]。水稻突变后导致穗数和穗粒数减少,穗长变短,结实率降低,穗型直立,同时光合能力下降,参与调控气孔保卫细胞的发育[19-20]。通过对直立穗基因的克隆,利用分子标记结合常规育种手段,可以快速、有效地选育出高产水稻品种[21-22]。【本研究切入点】尽管直立穗水稻品种的培育较早,但关于直立穗基因的研究较少,调控该性状的分子机制仍不够清楚。【拟解决的关键问题】本研究通过MNU(N-甲基-N-亚硝基脲)化学诱变水稻品种宁粳36,获得一个直立短穗突变体;通过图位克隆和基因组测序,克隆控制该突变表型的基因;利用转录组研究分析该基因参与的生物学途径,进而解析穗部发育的分子机制。
1 材料与方法
1.1 试验材料
水稻直立短穗突变体来源于粳稻品种宁粳36的MNU化学诱变突变体库,经过多代自交选择,性状稳定遗传。
1.2 农艺性状调查
突变体和野生型于2017年5月在合肥同时播种移栽,成熟期调查株高、有效穗数、穗长、每穗粒数、剑叶长、剑叶宽、千粒重等主要农艺性状,10次重复。
1.3 基因定位与克隆
构建突变体和PA64籼粳杂交F2定位群体,利用54个具有突变性状的F2典型单株和覆盖水稻12条染色体的分子标记对目的基因进行初定位(电子附表1)。通过基因组测序,分析定位区间内突变体和野生型之间的序列差异,进一步测序验证,发现变异位点,初步确定候选基因。
1.4 生物信息学分析
通过RGAP数据库(http://rice.plantbiology.msu. edu/cgi-bin/gbrowse/rice/)预测候选基因功能;利用NCBI数据库(http://www.ncbi.nlm.nih.gov/)分析ESP的同源蛋白,利用MEGA v4.1软件对ESP进化树分析;通过RiceXPro数据库(http://ricexpro.dna.affrc.go. jp/)对进行表达模式分析。
1.5 转录组分析
利用天根RNA提取试剂盒(RNA Prep Pure Plant kit)提取突变体和野生型幼穗中的总RNA,琼脂糖电泳和Nanodrop分别检测RNA的完整性和纯度,保证提取的RNA达到建库要求;转录组分析由北京诺禾致源公司完成。
表达差异基因通过GO聚类分析,根据差异基因在分子功能、生物学过程和细胞组分上的分布,筛选不同的GO term进行分析;根据KEGG数据库,分析植物激素信号转导和内质网代谢途径中相关基因的表达。
1.6 RNA提取、反转录以及qRT-PCR分析
利用天根RNA提取试剂盒(RNA Prep Pure Plant kit)提取突变体和野生型幼穗中的总RNA。剪取约100 mg孕穗期新鲜幼穗组织,液氮研磨成粉状,根据试剂盒操作步骤提取组织中的总RNA,琼脂糖电泳和Nanodrop分别检测RNA的完整性和纯度。进一步利用TaKaRa反转录试剂盒(SuperScript II kit)将RNA反转成cDNA,-20℃保存备用。
利用QuantStudio®3(ABI)定量PCR仪,参考SYBR®Premix ExTMkit(TaKaRa)试剂盒说明书进行qRT-PCR分析,反应体系为cDNA模板2 µL(质量为50—100 ng)、10 μmol·L-1前后引物各0.8 μL、SYBR Premix ExII 10 μL、50×ROX Reference Dye II 0.4 μL,补ddH2O至20 μL,3次样品重复和2次生物学重复。试验中所用到的定量引物见电子附表1,(,Ubq)为内参对照,利用2-ΔΔCT法分析相对基因表达结果。
2 结果
2.1 直立短穗突变体esp的表型分析
与野生型相比,突变体从抽穗期至成熟期株高降低,穗型由弯曲变为直立(图1-A和图1-B),突变体粒长变短约9.12%,粒宽增加约25.09%(图1-C、图1-D和图1-E)。农艺性状调查显示突变体株高、穗长和每穗粒数明显低于野生型,剑叶长和千粒重增加,然而有效穗数和剑叶宽与野生型无显著差异(表1)。
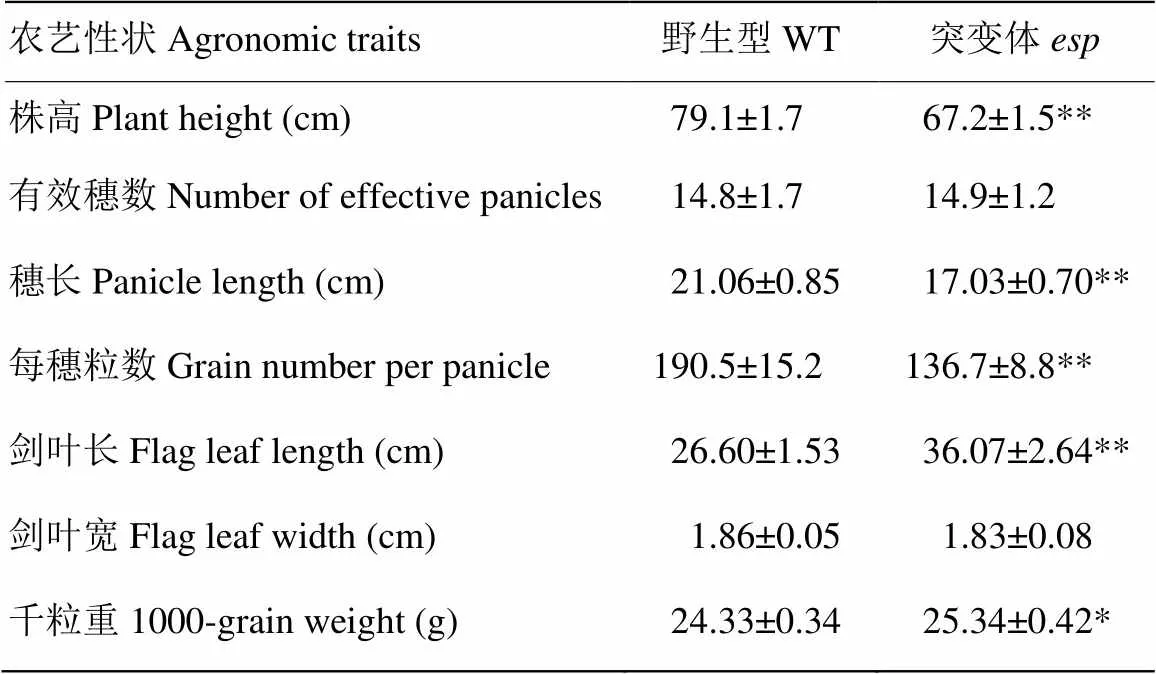
表1 突变体和野生型的农艺性状分析
数据为10次重复数据平均值,来源于2017年合肥正季。*和**分别代表差异显著(<0.05)和极显著(<0.01)
Values are the mean±SD from ten replications in Hefei, in 2017. *and ** separately indicate significantly different at<0.05 and<0.01
2.2 ESP的克隆
利用突变体与籼稻品种PA64杂交,F1植株表现为野生型性状,F2群体中出现弯穗和直立穗表型分离。利用54个具有突变表型的单株以及与突变表型连锁的分子标记将目的基因定位于水稻第7染色体长臂上Indel(insertion-deletion)标记C7-11和C7-14之间7.58 Mb区间内(图2-A)。通过对突变体和野生型进行基因组测序,结合定位结果以及测序验证,发现在其第6内含子与第7外显子连接位点由碱基G变异为A,导致第6内含子不能被剪切,蛋白翻译提前终止(图2-B);该基因与已报道的水稻基因/为等位基因,/突变体与表型类似,表明即为控制突变表型的目的基因。
2.3 ESP及其同源蛋白的序列分析
由10个外显子和9个内含子组成,编码一个含1 365个氨基酸残基的蛋白。序列分析显示ESP不含任何结构域,功能未知。突变基因编码一个含191个氨基酸残基的蛋白(包括182个原有氨基酸和9个新氨基酸)。进化树分析显示ESP蛋白广泛存在于单子叶和双子叶植物中(图3)。
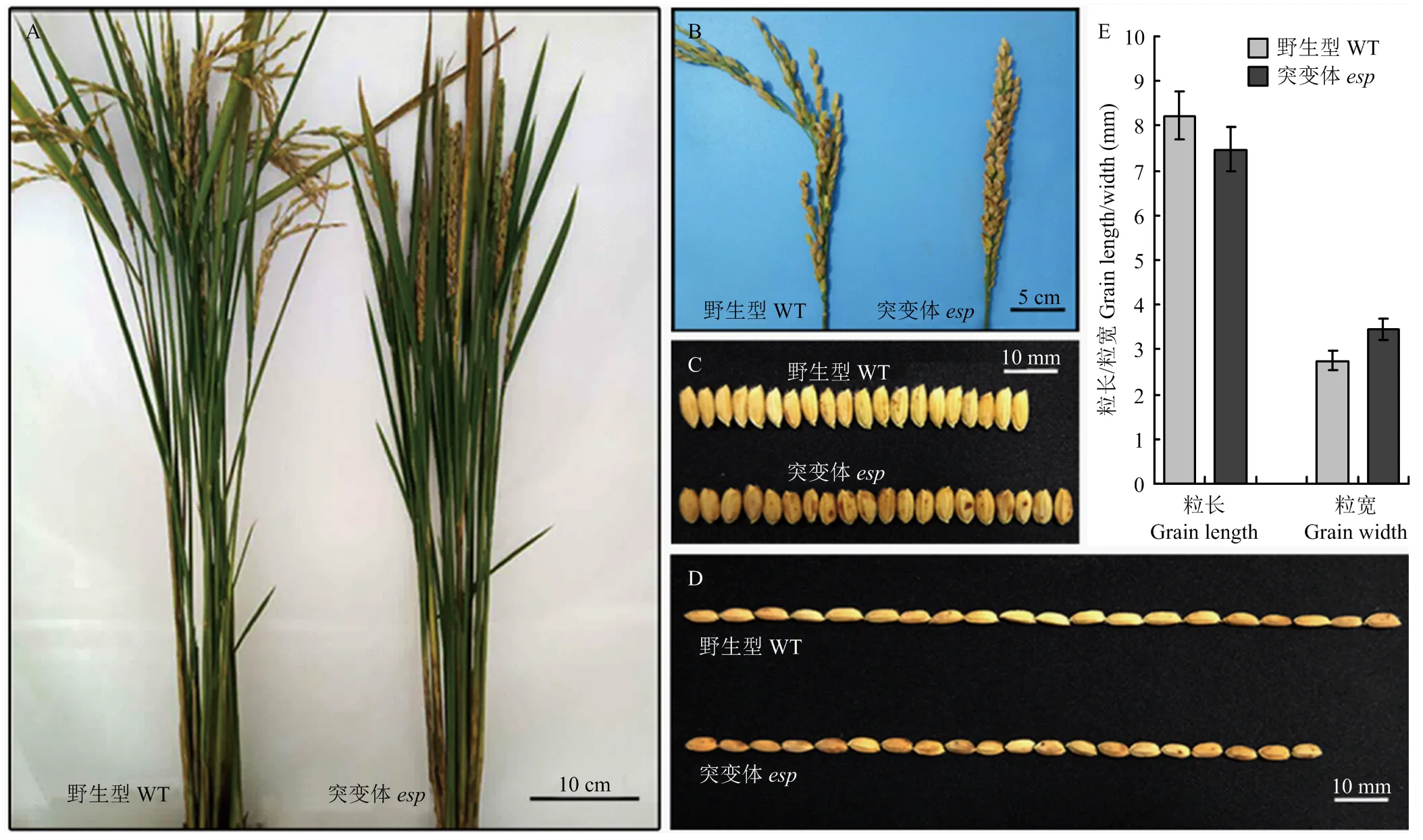
A:成熟期植株表型;B:成熟期穗部表型;C、D和E:种子粒宽和粒长测定
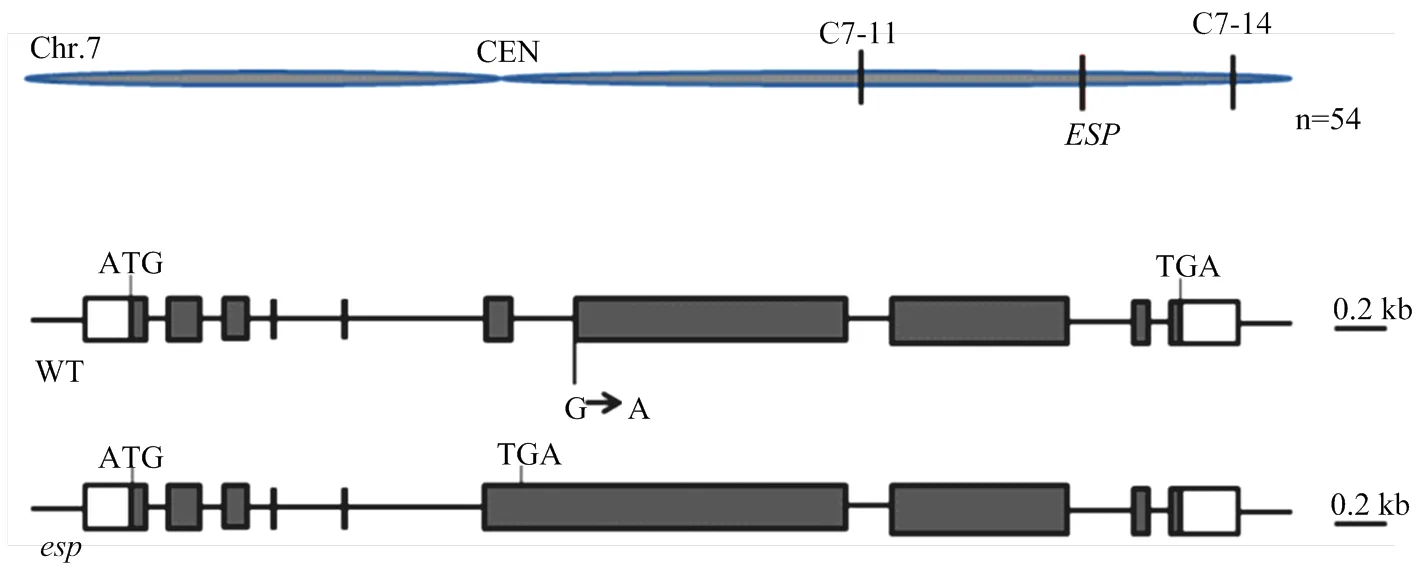
A:ESP初定位;B:ESP结构和esp突变位点;ATG和TGA分别代表起始和终止密码子
2.4 ESP组织表达分析
水稻表达模式数据库(RiceXPro)分析表明是一个组成型表达基因,其在水稻发育的不同时期和不同组织器官中都有表达,尤其在茎秆、花序、雌蕊、内外稃和子房中高度表达(图4-A),表达水平随着子房变大而逐渐降低(图4-B)。
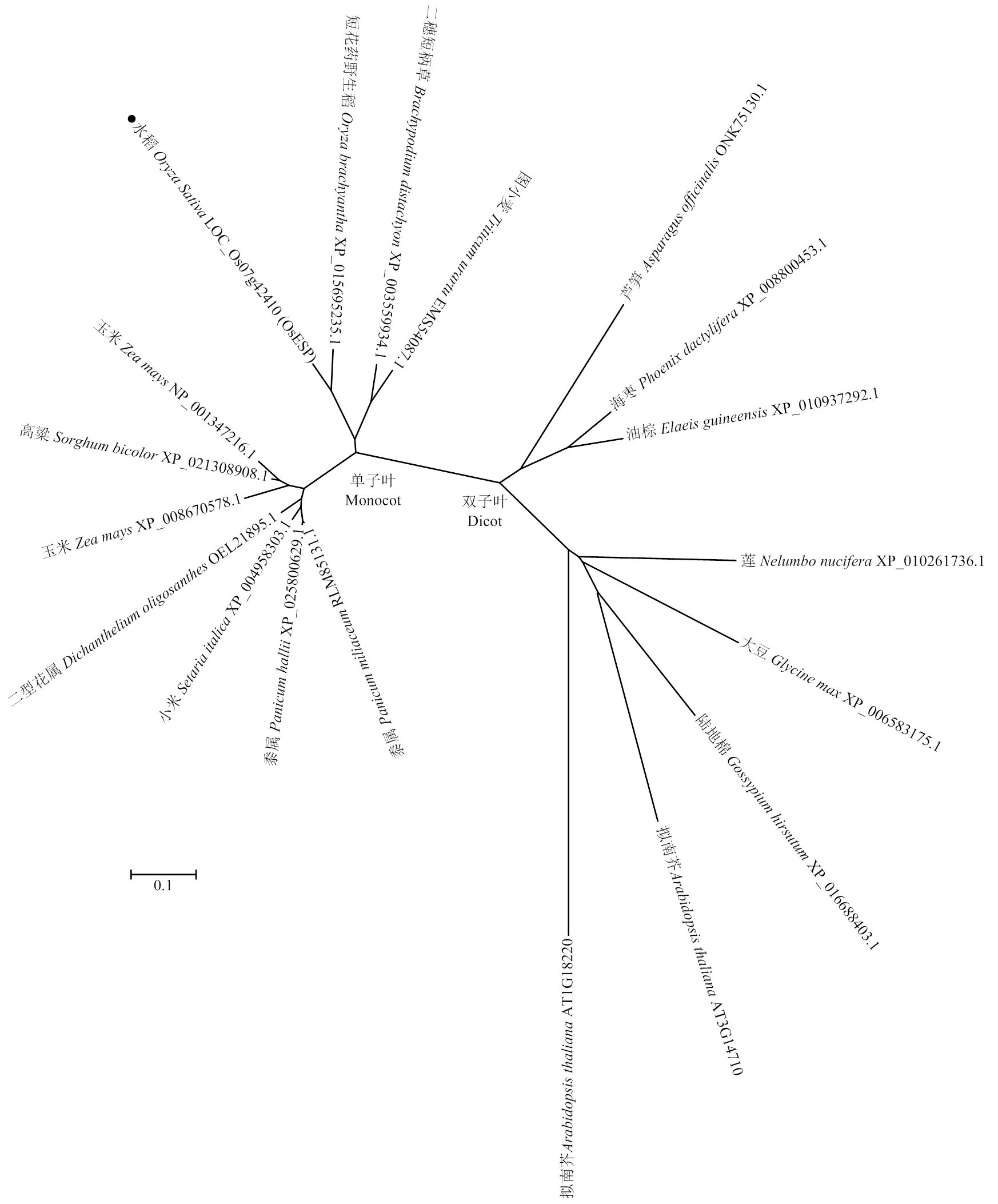
图3 ESP及其相关蛋白的进化树分析
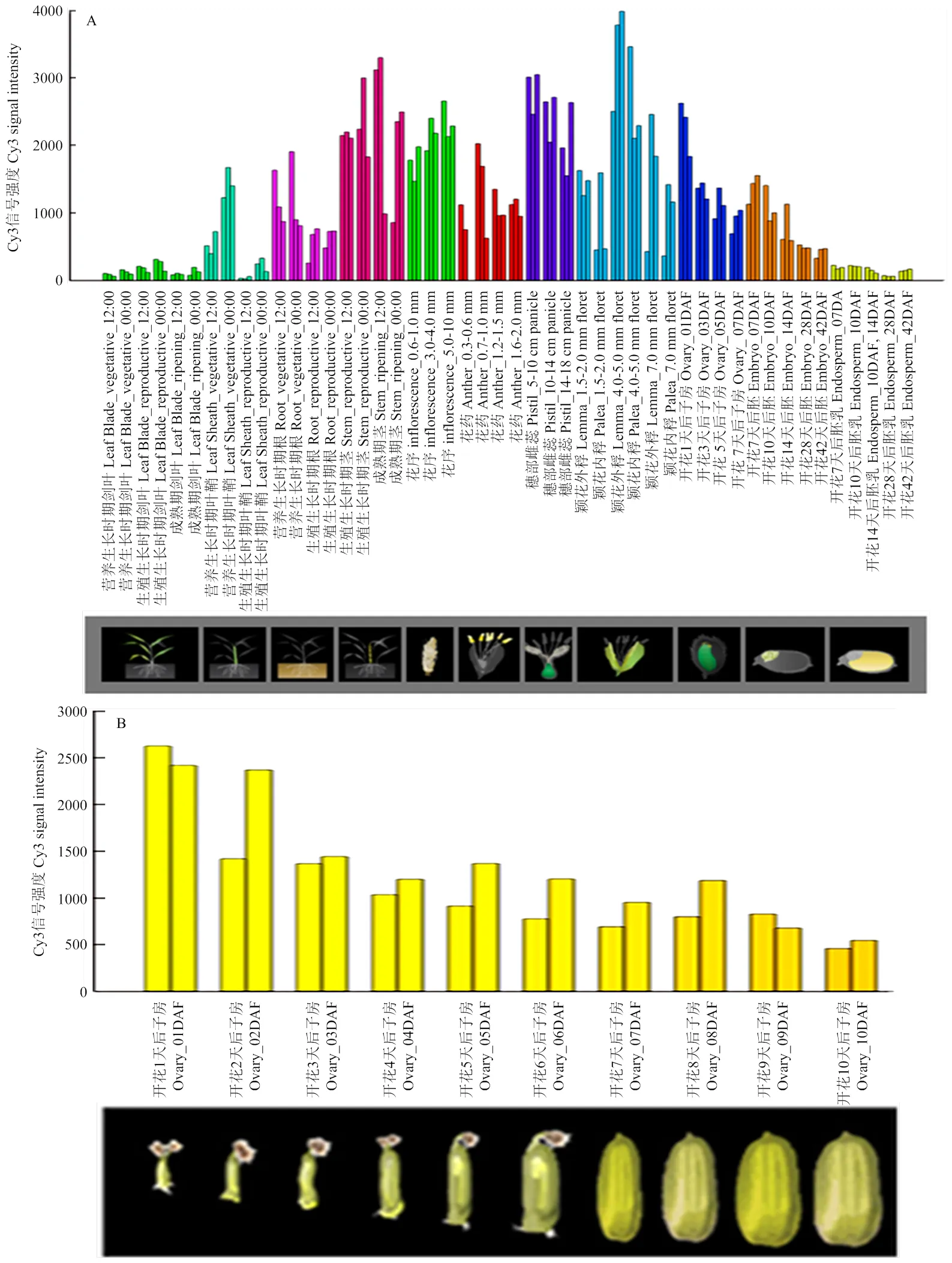
A:不同时期不同组织中ESP的表达模式;B:ESP在子房发育过程中的表达分析。数据来源于水稻表达模式数据库RiceXPro
2.5 突变体esp和野生型幼穗的转录组分析
为了解析的分子机理,利用转录组分析突变体和野生型幼穗中的基因表达,结果显示,与野生型相比,突变体幼穗中675个基因表达上调,1 074个基因表达下调(图5-A);表达差异>1.5倍的基因630个(<0.01),其中235个表达上调,395个表达下调(图5-B),这些基因涉及到15项生物学过程、3项细胞组分和12项分子功能(图5-C)。
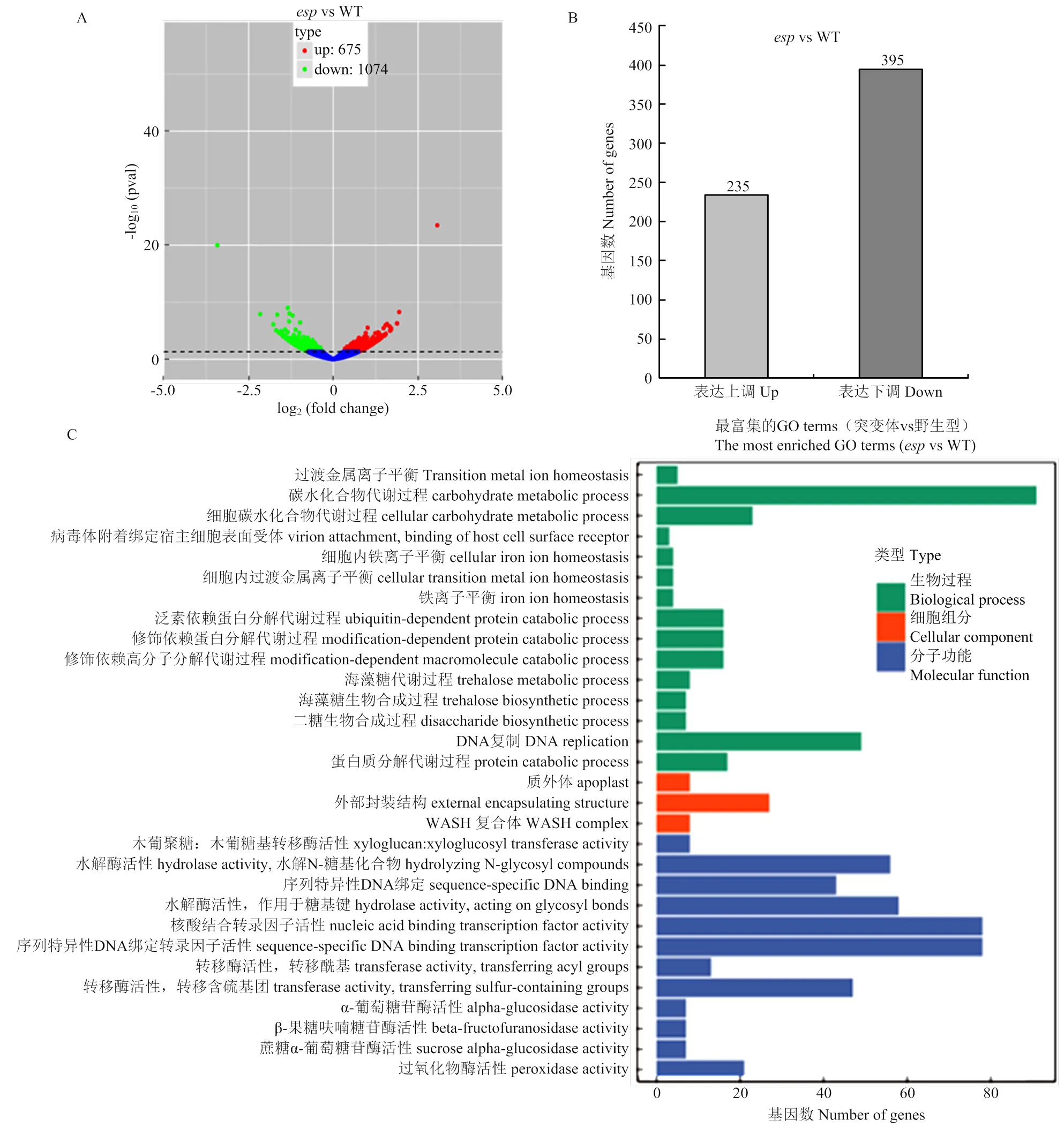
A:突变体esp和野生型幼穗转录组分析火山图;红点和绿点分别代表上调基因和下调基因,蓝点代表无显著差异表达基因;B:表达差异>1.5倍的上调和下调基因数目;C:表达差异基因的功能分类
2.6 ESP突变对植物激素信号转导和内质网代谢途径的调控
根据转录组测序结果,与野生型相比,突变体中检测到包括次生代谢产物生物合成、淀粉和蔗糖代谢、内质网蛋白加工等11个代谢通路受到明显调控(<0.1)(表2)。
突变后,植株多个性状发育受到抑制,推测可能参与植物激素信号转导。结果表明,与野生型相比,突变体幼穗中与生长素信号转导有关的基因()、()、()、()和()上调表达,()下调表达;与细胞分裂素有关的基因()上调表达;与脱落酸有关的基因/()、()和()下调表达;与乙烯有关的基因()、()和()下调表达,()上调表达;与茉莉酸有关的基因()和()上调表达,()下调表达(图6-A和图6-B)。

表2 代谢通路及其差异性表达基因数量
亚细胞定位显示EP2是一个内质网蛋白[15],可能参与调控内质网代谢,转录组结果显示,内质网蛋白加工有关的代谢过程在突变体中受到明显的调控(图7-A)。包括与内质网中蛋白绑定和运输有关的基因()上调表达、()和()下调表达;与内质网中多聚糖合成有关的基因()、()和()上调表达;参与内质网蛋白的折叠、分选和降解有关的基因()、()、()、()、()、()、()、()、()、()、()、()、()、()、()、()和()下调表达,()、()、()和()上调表达(图7-B)。
2.7 转录组验证
为了进一步验证转录组结果,对植物激素信号转导和内质网蛋白质加工途径相关基因进行定量分析。结果与转录组数据一致,同时发现与油菜素内酯信号转导相关基因()和()在突变体和野生型之间无显著差异(图8-A和图8-B),表明转录组结果准确。
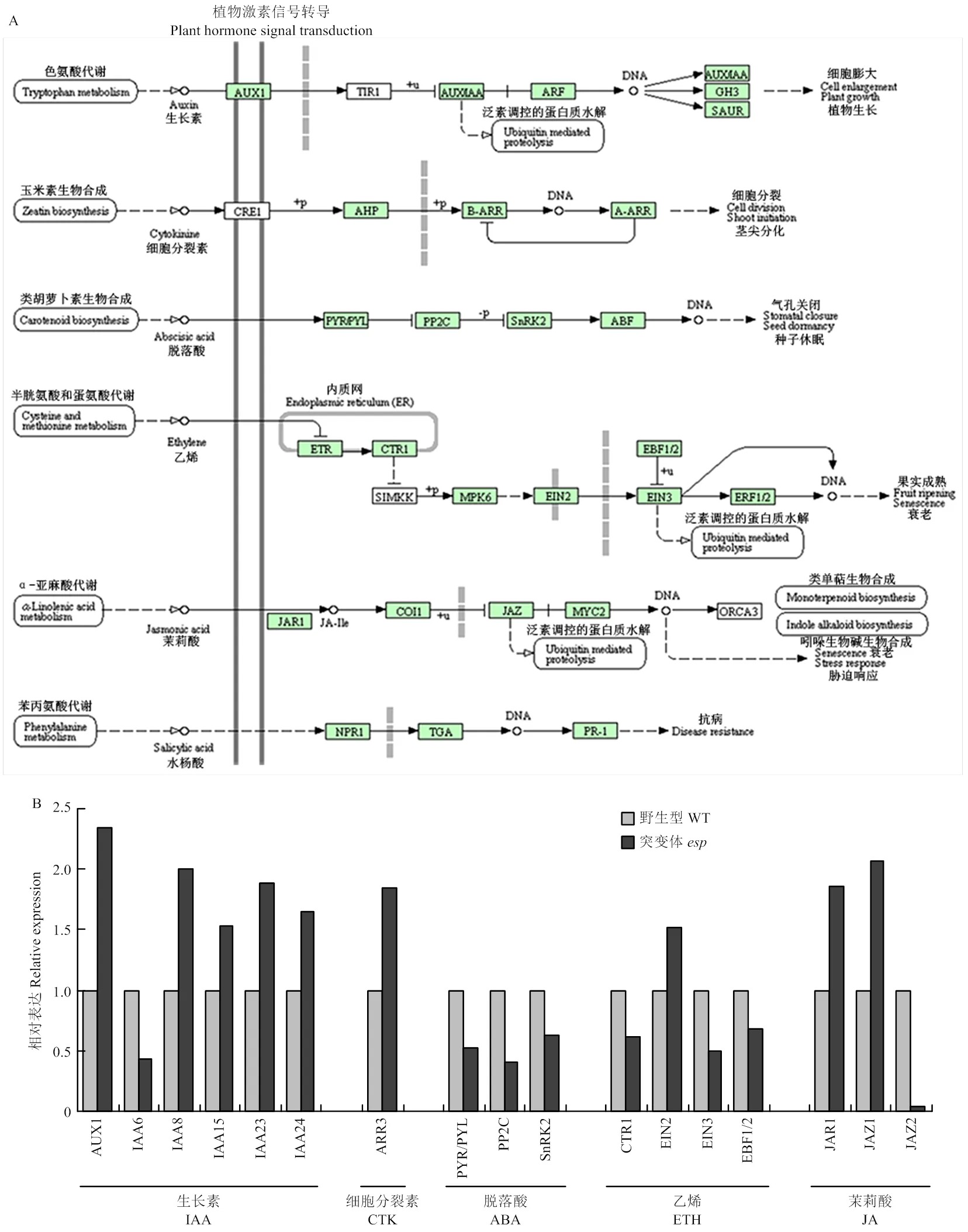
A:植物激素信号转导KEGG代谢途径;B:转录组分析植物激素信号转导途径相关基因的表达
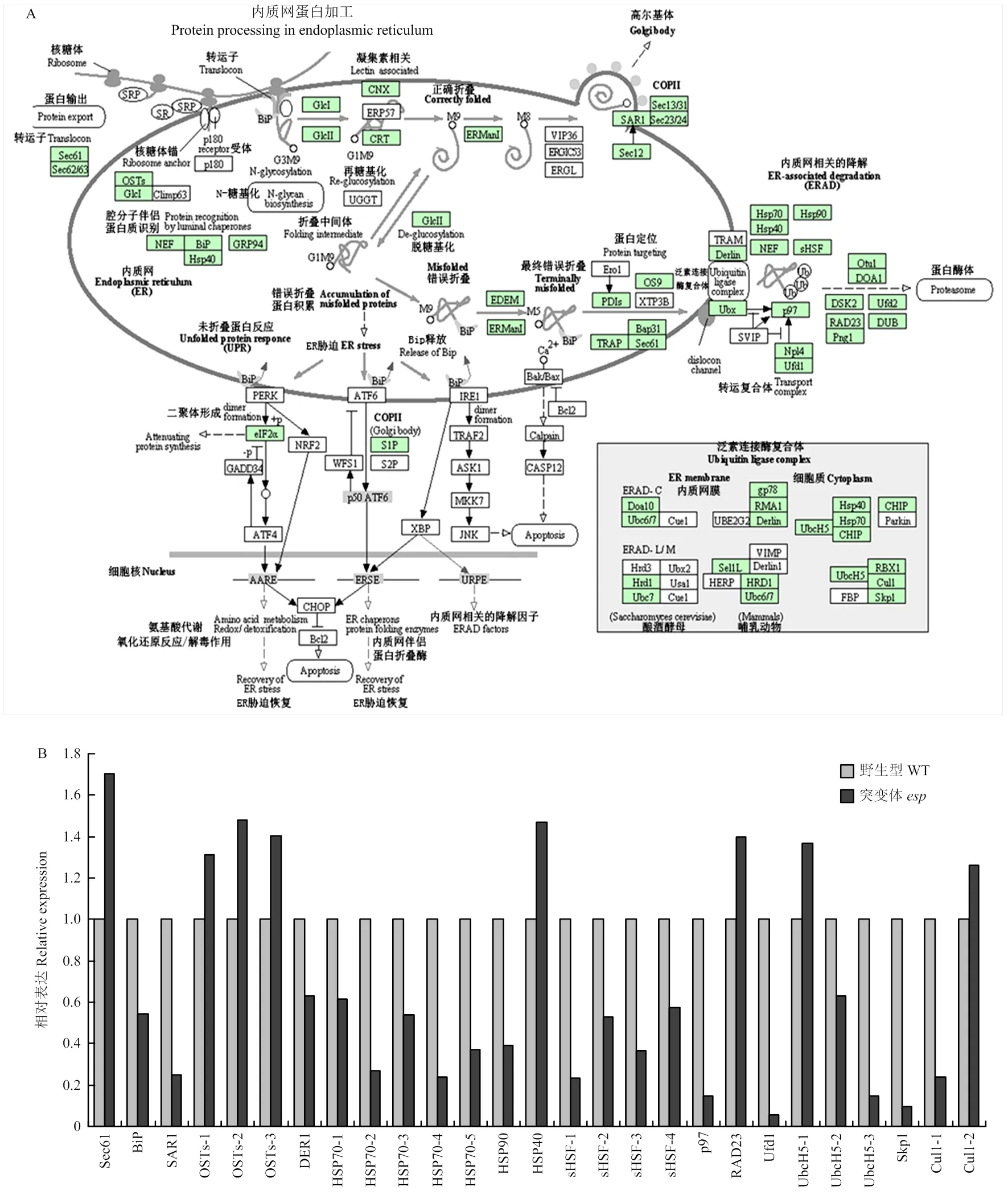
A:内质网蛋白质加工KEGG代谢途径;B:转录组分析内质网蛋白质加工途径相关基因的表达
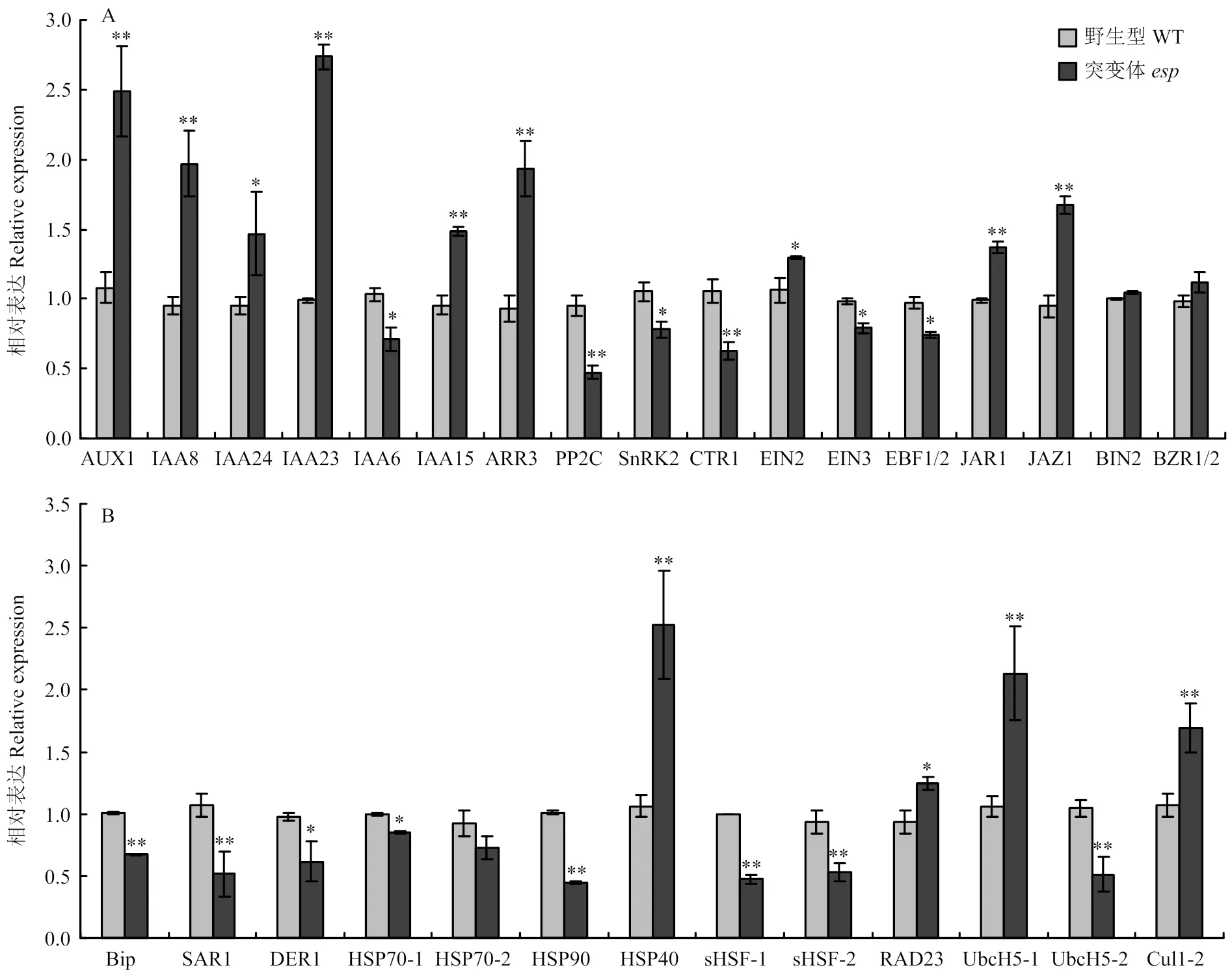
A:植物激素信号转导途径相关基因的表达分析;B:内质网蛋白质加工途径相关基因的表达分析。*和**分别代表t测验下差异显著和极显著水平
3 讨论
水稻株型一直是水稻遗传和育种中研究的重要性状之一,理想的株型能够通过提高植株光合效率、调节群体结构等增加产量;穗部结构是修饰水稻株型的重要因素之一。水稻直立穗株型可以增加植株的透光率,提高光能的利用效率,进而增加产量[6]。目前,大量的直立穗粳稻品种在生产上被广泛种植和推广[7]。然而,关于控制直立穗性状的基因研究较少,调控该性状的分子机制仍不够清楚。
本研究克隆了一个控制直立短穗表型的基因,该基因突变后株高降低,穗长变短,每穗粒数减少,粒长变短,粒宽和千粒重增加(图1-E和表1)。研究发现与已克隆水稻基因/为等位基因。Zhu等[15]鉴定2个分别由籼稻品种中籼3037和9311突变而来的等位突变体和;与野生型相比,和株高降低,穗长和粒长变短,粒宽增加,千粒重降低,然而不同的是每穗粒数减少,每穗粒数增加。此外,另2个等位突变体和分别由粳稻品种中花11和日本晴变异获得,和与野生型相比,株高降低,穗长和粒长变短,粒宽增加,千粒重降低,每穗粒数无显著差异[14]。由此可见,等位变异的突变体相对于野生型,株高、穗长、粒长均降低和变短,粒宽增加;然而,不同突变体中每穗粒数和千粒重变化较大,和中每穗粒数减少,中每穗粒数增加,和中无明显差别;中千粒重增加,、和中千粒重降低(表1和表3)。这些表型的差异可能与品种背景以及突变位点有关。
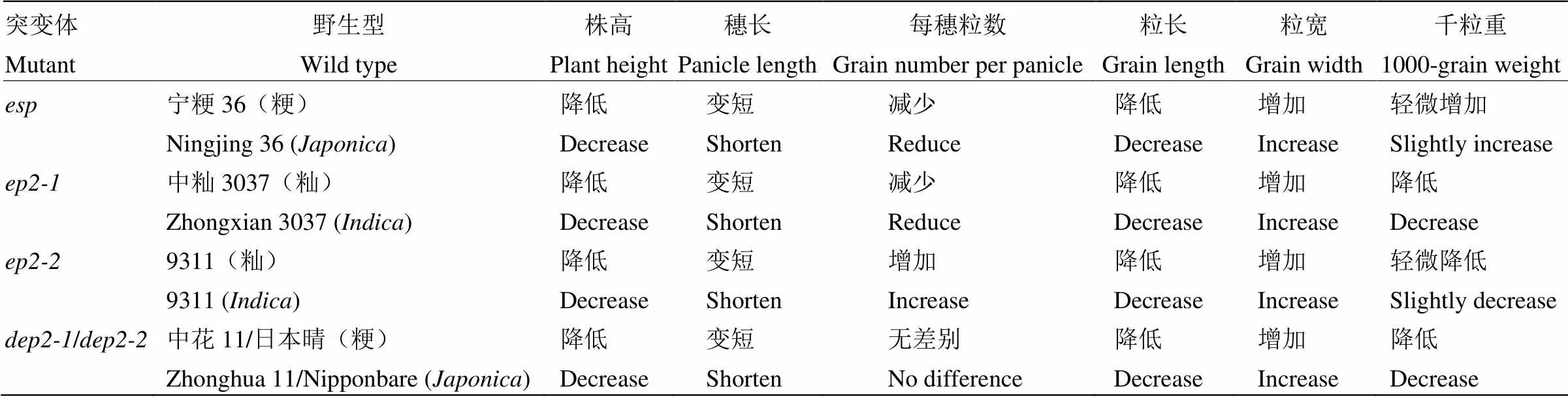
表3 直立穗基因ESP等位突变体表型特点
通过研究发现突变导致不同组织、不同性状发生变异,可能影响多个代谢途径。利用转录组分析显示630个与次生代谢产物合成、内质网蛋白加工等多个生物学过程有关的基因在突变体中受到显著调控(图5-B和图5-C)。植物激素通过影响细胞增殖和生长,进而控制粒型的发育;突变体不同组织细胞长度与野生型相比无显著差异,降低的穗长可能由于细胞增殖受到抑制[14],小圆粒突变体中细胞长度和细胞数量都明显减少[16],表明可能与生长素、细胞分裂素、油菜素内酯等激素的信号转导有关;转录组研究和定量分析表明生长素和细胞分裂素信号转导有关的基因表达在突变体幼穗中受到调控,然而,油菜素内酯信号转导有关的基因在突变体和野生型之间无显著差异(图6-A、图6-B和图8-A);这表明突变可能不参与调控突变体幼穗中油菜素内酯相关基因的表达。水稻基因编码IAA-葡萄糖水解酶,使IAA-葡萄糖水解成IAA和葡萄糖,通过调节生长素的水平调控粒型的发育[23];过量表达生长素响应因子能够导致水稻粒型变小、株高降低[24-25];通过调节植株细胞分裂素的水平,调控粒长和穗粒数[26]。水稻等位基因突变后导致植株矮化、每穗粒数增加,表现对油菜素内酯、赤霉素等敏感,但与油菜素內酯突变体间不存在遗传上位性,暗示可能与植物激素无关[27]。说明可能通过调节植物激素有关基因的表达,调控激素水平,进而调节植株穗型和粒型的发育。
直立穗株型能够改良株型结构和群体质量,提高植株生长速率和产量,改善抗倒性和耐肥性,育种利用潜力大[8, 12, 28-31]。解析的不同等位变异与水稻产量之间的关系,不仅能够为水稻育种提供丰富的种质资源,为水稻高产育种提供潜在利用价值,同时有助于理解穗部发育的分子机制。
4 结论
鉴定一个直立短穗突变体,并克隆控制该突变性状的基因,其与已报道的水稻基因/为等位基因。广泛存在于单子叶和双子叶植物中,且在茎秆、花序、雌蕊、内外稃和子房中高度表达,随着子房变大表达水平逐渐降低。可能通过调节植物激素信号转导、内质网蛋白加工过程中相关基因的表达进而影响穗部发育。
[1] Ashikari M, Sakakibara H, Lin S, Yamamoto T, Takashi T, Nishimura A, Angeles E R, Qian Q, Kitano H, Matsuoka M. Cytokinin oxidase regulates rice grain production., 2005, 309: 741-745.
[2] Jin J, Huang W, Gao J P, Yan J, Shi M, Zhu M Z, Luo D, Lin H X. Genetic control of rice plant architecture under domestication., 2008, 40: 1365-1369.
[3] Li P, Wang Y, Qian Q, Fu Z, Wang M, Zeng D, Li B, Wang X, Li J.controls rice shoot gravitropism through regulating polar auxin transport., 2007, 17: 402-410.
[4] Li S, Qian Q, Fu Z, Zeng D, Meng X, Kyozuka J, Maekawa M, Zhu X, Zahng J, Li J, Wang Y.encodes a putative PTR family transporter and determines rice panicle size., 2009, 58: 592-605.
[5] Yu B, Lin Z, Li H, Li X, Li J, Wang Y, Zhang X, Zhu Z, Zhai W, Wang X, Xie D, Sun C., a major quantitative trait locus controlling tiller angle in rice., 2007, 52: 891-898.
[6] Huang X, Qian Q, Liu Z, Sun H, He S, Luo D, Xia G, Chu C, Li J, Fu X. Natural variation at thelocus enhances grain yield in rice., 2009, 41: 494-497.
[7] Yan C J, Zhou J H, Yan S, Chen F, Yeboah M, Tang S Z, Liang G H, Gu M H. Identification and characterization of a major QTL responsible for erect panicle trait inrice (L.)., 2007, 115: 1093-1100.
[8] Zhou Y, Zhu J, Li Z, Yi C, Liu J, Zhang H, Tang S, Gu M, Liang G. Deletion in a quantitative trait geneassociated with panicle erectness improves plant architecture during rice domestication., 2009, 183: 315-324.
[9] Sun H, Qian Q, Wu K, Luo J, Wang S, Zhang C, Ma Y, Liu Q, Huang X, Yuan Q, Han R, Zhao M, Dong G, Guo L, Zhu X, Gou Z, Wang W, Wu Y, Lin H, Fu Q. Heterotrimeric G proteins regulate nitrogen-use efficiency in rice., 2014, 46: 652-656.
[10] Yi X, Zhang Z, Zeng S, Tian C, Peng J, Li M, Lu Y, Meng Q, Gu M, Yan C. Introgression of qPE9-1 allele, conferring the panicle erectness, leads to the decrease of grain yield per plant in japonica rice (L.)., 2011, 38: 217-223.
[11] Liu Q, Han R, Wu K, Zhang J, Ye Y, Wang S, Chen J, Pan Y, Li Q, Xu X, Zhou J, Tao D, Wu Y, Fu X. G-protein βγ subunits determine grain size through interaction with MADS-domain transcription factors in rice., 2018, 9: 852.
[12] 陈温福, 徐正进, 张龙步, 张文忠, 马殿荣. 北方粳型稻超高产育种理论与实践. 中国农业科学, 2007, 40(5): 869-874.
Chen W F, Xu Z J, Zhang L B, Zhang W Z, Ma D R. Theories and practices of breeding japonica rice for super high yield., 2007, 40(5): 869-874. (in Chinese)
[13] Wang J, Nakazaki T, Chen S, CHEN W, SAITO H, TSUKIYAMA T, OKUMOTO Y, XU Z, TANISAKA T. Identification and characterization of the erect-pose panicle geneconferring high grain yield in rice (L.)., 2009, 119: 85-91.
[14] Li F, Liu W, Tang J, Chen J, Tong H, Hu B, Li C, Fang J, Chen M, Chu C. Rice DENSE AND ERECT PANICLE 2 is essential for determining panicle outgrowth and elongation., 2010, 20: 838-849.
[15] Zhu K, Tang D, Yan C, Chi Z, Yu H, Chen J, Liang J, Gu M, Cheng Z.encodes a novel protein that regulates panicle erectness inrice., 2010, 184: 343-350.
[16] Abe Y, Mieda K, Ando T, Kono I, Yano M, Kitano H, Iwasaki Y. The(/) gene is involved in the regulation of seed size in rice., 2010, 85: 327-339.
[17] 朱海涛, 柯善文, 冯小龙, 邹龙海, 曾秀瑜, 张向前. 水稻直立穗突变体的鉴定及其候选基因分析. 华北农学报, 2014, 29(4): 44-48.
Zhu H T, Ke S W, Feng X L, Zou L H, Zeng X Y, Zhang X Q. Identification of an erect panicle mutant, 2014, 29(4): 44-48. (in Chinese)
[18] Qiao Y, Piao R, Shi J, Lee S I, Jiang W, Kim B K, Lee J, Han L, Ma W, Koh H J. Fine mapping and candidate gene analysis of,, which confers high grain yield in rice (L.)., 2011, 122: 1439-1449.
[19] Piao R, Jiang W, Ham T H, Choi M S, Qiao Y, Chu S H, Park J H, Woo M O, Jin Z, An G, Lee J, Koh H J. Map-based cloning of thegene in rice., 2009, 119: 1497-1506.
[20] Yu H, Murchie E H, Gonzalez-Carranza Z H, Pyke K A, Roberts J A. Decreased photosynthesis in the() mutant of rice is associated with reduced stomatal conductance and attenuated guard cell development., 2015, 66(5): 1543-1552.
[21] 朱金燕, 王军, 杨杰, 范方军, 杨金欢, 仲维功. 一个与水稻直立穗基因关联的InDel标记的设计与验证. 分子植物育种, 2012, 10(5): 583-588.
Zhu J Y, Wang J, Yang J, Fan F J, Yang J H, Zhong W G. Development and application for a functional marker for erect panicle geneof rice., 2012, 10(5): 583-588. (in Chinese)
[22] 胡运高, 郭连安, 杨国涛, 钦鹏, 范存留, 彭友林, 严维, 何航, 李仕贵. 直立密穗基因的遗传分析及在杂交稻中的育种利用. 遗传, 2016, 38(1): 72-81.
Hu Y G, Guo L A, Yang G T, Qin P, Fan C L, Peng Y L, Yan W, He H, Li S G. Genetic analysis of dense and erect panicle 2 alleleand its application in hybrid rice breeding., 2016, 38(1): 72-81. (in Chinese)
[23] Ishimaru K, Hirotsu N, Madoka Y, Murakami N, Hara N, Onodera H, Kashiwagi T, Ujiie K, Shimizu B, Onishi A, Miyagawa H, Katoh E. Loss of function of the IAA-glucose hydrolase geneenhances rice grain weight and increases yield., 2013, 45(6): 707-711.
[24] Zhang S, Wang S, Xu Y, Yu C, Shen C, Qian Q, Geisler M, Jiang de A, Qi Y. The auxin response factor, OsARF19, controls rice leaf angles through positively regulating OsGH3-5 and OsBRI1., 2015, 38(4): 638-654.
[25] Zhang S, Wu T, Liu S, Liu X, Jiang L, Wan J. Disruption of OsARF19 is critical for floral organ development and plant architecture in rice (L.)., 2016, 34(4): 748-760.
[26] Jin J, Hua L, Zhu Z, Tan L, Zhao X, Zhang W, Liu F, Fu Y, Cai H, Sun X, Gu P, Xie D, Sun C.encodes a secreted peptide that regulates grain number, grain length and awn development in rice domestication., 2016, 28(10): 2453-2463.
[27] Taguchi-Shiobara F, Kawagoe Y, Kato H, Onodera H, Tagiri A, Hara N, Miyao A, Hirochika H, Kitano H, Yano M, Toki S. A loss-of-function mutation of ricecauses semi-dwarfness and slightly increased number of spikelets., 2011, 61(1): 17-25.
[28] 赵明珠, 王青营, 孙健, 徐正进, 陈温福. 水稻直立穗型基因研究进展. 中国科学: 生命科学, 2017, 47(10): 1036-1042.
Zhao M Z, Wang Q Y, Sun J, Xu Z J, Chen W F. Research progress of rice erect panicle gene., 2017, 47(10): 1036-1042. (in Chinese)
[29] 高士杰, 陈温福, 张龙步. 直立穗型水稻的研究. 吉林农业科学, 1999, 24(6): 12-15.
Gao S J, Chen W F, Zhang B L. Studies of erect panicle in rice., 1999, 24(6): 12-15. (in Chinese)
[30] Wang Y, Li J. Molecular basis of plant architecture., 2008, 59: 253-279.
[31] Xu Z J, Chen W F, Zhang L B, Yang S R. Design principles and parameters of rice ideal panicle type., 2005, 50: 2253-2256.
Transcriptome Research of Erect and Short PanicleMutantin Rice
ZHOU KunNeng, XIA JiaFa, YUN Peng, WANG YuanLei, MA TingChen, ZHANG CaiJuan, LI ZeFu
(Rice Research Institute, Anhui Academy of Agricultural Sciences/Key Laboratory for Rice Genetics and Breeding of Anhui Province, Hefei 230001)
【Objective】In this study, we aimed to identify thegene, whose mutation caused a phenotype, namely, and to determine its regulatory role in the gene network that controls the related agronomic traits (e.g., plant types and panicle length).【Method】In this study, agronomic traits, such as plant height, panicle length and grain length at mature stages, were used as phenotypic marks to trace themutant. Individuals carrying mutant phenotypes were selected from the F2population to cross withandfor further gene mapping and genome sequencing that were used to map the potential mutation region/sites. The bioinformatics software was used to analyze phylogenetic tree and gene expression. The total RNAs isolated from wild type and mutants were used for transcriptome RNA-seq analysis. The differential expressed genes and expression levels of genes related to plant hormone signal transduction and protein processing in endoplasmic reticulum were analyzed by GO software and KEGG database. The transcriptome data were verified by qRT-PCR.【Result】Phenotypic analysis showed that themutant exhibited a erect panicle architecture. The plant height, panicle length, grain length and the number of spikelets per panicle were decreased in themutants, when compared to wild type control, whereas grain width and the 1 000-grain weight were increased, although no obvious difference in the number of effective panicles between mutants and control. Thegene was mapped to a 7.58 Mb interval between markers C7-11 and C7-14 on the long arm of rice 7th chromosome by using the F2population ofmutant and PA64. Genome sequencing demonstrated that a single nucleotide change (G to A) at the junction of 6th intron and 7th exon of, which led to a splicing defect, causing premature protein translation. Thewas allelic to/. Phylogenetic analysis revealed that the ESPs are widely present in monocot and dicot plants. Expression analysis predicted thatgene was highly expressed in stem, inflorescence, pistil, glumelle, lemma and ovary, and the expression level was gradually decreased with the ovary inflation. Transcriptome RNA-seq analysis of young panicle identified 630 differential expressed genes inmutants versus wild type, including 235 up-regulated and 395 down-regulated. GO and qRT-PCR analysis revealed that genes involved in plant hormone signal transduction and protein processing in endoplasmic reticulum were misregulated inmutants.【Conclusion】Thegene was allelic to/, the mutation of which leads to multiple phenotypes, such as decreased plant height and shorter panicle length. Transcriptome analysis suggested that thegene might affect plant development by regulating genes expression associated with plant hormone signal transduction and protein processing in endoplasmic reticulum.
rice (L.); erect and short panicle mutant; gene cloning; phylogenetic analysis; transcriptome analysis
2019-08-05;
2019-10-21
国家重点研发计划(2016YFD0100101-06和2017YFD0100406)、国家自然科学基金(31801331)、安徽省科技重大专项(18030701175)、安徽省农业科学院科技创新团队项目(18C0101)
周坤能,E-mail:zhoukunneng1986@163.com。通信作者李泽福,E-mail:lizefu@aliyun.com
(责任编辑 李莉)
