蛭态轮虫中国新记录种及其研究展望
2020-04-06崔宗斌杨宇峰
李 莹 王 庆 魏 南 曾 悦 崔宗斌 杨宇峰
(1. 暨南大学生态学系, 广东省高校水体富营养化与赤潮防治重点实验室, 热带亚热带水生态工程教育部工程研究中心, 广州510632; 2. 中国科学院水生生物研究所淡水生态与生物技术国家重点实验室, 武汉 430072)
轮虫是一类体形较小的后生假体腔动物(50—2000 μm), 蛭态类轮虫广泛分布于各水生、半水生(苔藓、土壤、地衣等)等生境。根据现被广泛采用的分类系统, 轮虫动物门包含三个纲: 单巢纲、双巢纲和尾盘纲[1,2]。蛭态亚纲轮虫具有两个卵巢,终身行孤雌生殖, 在环境处于缺水或缺氧等恶劣情况时, 脱水可以进入隐生状态(Anhydrobiosis), 待环境好转, 重新恢复到正常的生活状态[3]。蛭态类轮虫是现今具有基因和形态特征的最古老且通过无性生殖进化的类群[4,5]。根据以上蛭态轮虫的特征,研究蛭态轮虫, 对揭开生物进化、物种分化以及隐生等机制等具有重要的推动作用。
敖突轮属(Otostephanos)作为宿轮科三属之一[6],具有宿轮科轮虫体内特有的食物泡(Food pellet), 轮盘伸展时带有的圆环状结构为该属轮虫最重要的分类学依据[3,7], 我国仅有该属记录一种。环颈敖突轮虫(Otostephanos torquatus)在我国蛭态轮虫研究中未被报道过, 作为敖突轮属的常见种, 该轮虫初始发现于苔藓样品中, 营自由生活, 摄食时, 其形态发生改变, 轮盘从头部伸展出来, 转动轮盘上的纤毛通过滤食方式经咀嚼器的作用将食物储存在内腔(Lumen)的食物泡中, 然后以“粪丸”的形式排出体外。
本研究通过对环颈敖突轮虫形态学特征描述,咀嚼器扫描电镜图片和COⅠ序列分析, 对该种的鉴定进行进一步确定。为我国今后蛭态轮虫分类和物种鉴定提供参考。
1 材料与方法
1.1 样品采集与活体轮虫获取
于2018年4月11日在广东省珠海市万山群岛(N21°56′15.81", E113°43′5.77")采集苔藓样品, 苔藓着生于石壁上, 常年处于阴暗潮湿的环境, 该苔藓植物含水量丰富, 适合蛭态轮虫生长繁殖。慢慢将苔藓撕下, 并放置于牛皮种子袋中, 待室温自然风干后密封保存。
对于附藓蛭态轮虫的提取, 采取震荡复苏法,取少量风干后的苔藓样品置于50 mL离心管中, 加入约40 mL纯水浸泡样品, 水需没过苔藓, 放置0.5h及以上, 剧烈震荡离心管, 将苔藓与附着的蛭态轮虫分离, 等待蛭态轮虫从休眠状态复苏, 后经20 μm筛网过滤, 加纯水重复过滤5—10次, 以便获取更多的轮虫, 此方法仅适用于蛭态轮虫的定性分析[8,9]。
1.2 形态学鉴定
使用显微镜(OLYMPUS BX51)进行外部形态学观察, 通常状态下蛭态轮虫处于未摄食状态, 待蛭态轮虫开始摄食后拍摄其头冠、咀嚼器等重要的分类学依据, 分类依据参考《Ordnung Bdelloidea》[10]。拍照后用T Capture软件测量该轮虫各部位长度, 测量模型参考Iakovenko等[11]。轮虫形态模式图使用Adobe illustrator CS6 2017软件绘制。
1.3 咀嚼器扫描电镜制片
在载玻片上滴加微量甘油, 将方形盖玻片(边长0.9 cm)固定于载玻片上, 保证盖玻片和载玻片表面均干净无杂质。用毛细玻璃管吸取一只蛭态轮虫放在盖玻片上(吸取的水滴尽可能小), 加入3%次氯酸钠溶液将轮虫溶解, 显微镜下观察虫体的溶解过程, 然后用毛细管将次氯酸钠溶液吸出, 加入超纯水冲洗5—10次, 直至咀嚼器周围无杂质和结晶,用马克笔标记咀嚼器的位置, 24h自然风干后, 喷金后在扫描电子显微镜(ZEISS EVO/MA15)下拍摄观察该轮虫咀嚼器形态, 具体咀嚼器制备过程参考De Smet[12]。
1.4 蛭态轮虫COⅠ基因克隆及序列分析
挑取环颈敖突轮虫若干只置于载玻片上, 用纯水清洗至表面无杂质, 将单个虫体放入0.2 mL PCR管中, 使用Hotshot方法提取单只蛭态轮虫样品的DNA[13], 向PCR管中加入碱性裂解液10 μL, 95℃裂解0.5h后取出立即放入-20℃冰箱内冷冻5min,冷却后加入中和液10 μL, 即完成蛭态轮虫的DNA提取, 随后进行PCR扩增, 1%琼脂糖凝胶电泳验证PCR产物后测序。引物反应程序: 95℃预变性1min→95℃变性40s→48℃退火40s→62℃延伸45s,循环5次 →94℃变性40s→ 循环退火40s→ 循环延伸45s, 循环35次, 最后72℃延伸5min→保存[14]。该过程使用引物为(ZplankF1_t1: TGTAAAACGACG GCCAGTTCTASWAATCATAARGATATTGG;ZplankR1_t1: CAGGAAACAGCTATGACTTCAG GRTGRCCRAARAATCA)[15]。测序结果与NCBI上已有序列进行Blast比对, 寻找亲缘关系相近的物种, 将测序结果经Bioedit软件处理后, 使用MEGA7.0软件分析该轮虫与同科及同属轮虫COⅠ序列之间的遗传距离, 采用邻接法(Neighbor-Joining method,NJ)构建系统发育树。

图 1 环颈敖突轮虫形态特征Fig. 1 Morphological characteristic of O. torquatus
2 结果
2.1 形态描述
环颈敖突轮虫身体纵长(图 1), 主要分为头部,躯干和足部三个部分, 无眼点, “卵生”。体内器官呈黄棕色, 表皮较光滑, 无附着大颗粒杂质, 有明显的纵长条纹。头部顶端的“吻突”内有二分裂半圆形喙片, 长有细短的纤毛。虫体最宽处通常位于躯干的后半部, 颈部较长, 背触手较短, 足部较短, 刺戟近似三角形, 趾3个; 体内含有大小不等的食物泡, 卵巢两个, 躯干两侧各一个(图 2)。该轮虫处于正常活动状态时, 轮盘位于头的内部, 不伸展, 趾吸附在附着物上, 依靠纵行肌的收缩前进。在摄食状态下, 轮盘从头部伸出, 左右各一个, 宽度明显大于头部, 通过转动其上方的纤毛进行滤食, 食物经咀嚼器储存在内腔的食物泡中。食物泡是宿轮科轮虫特有的结构, 食物在食物泡中被消化利用后, 整个食物泡通过泄殖腔以球形粪丸排出体外。头冠外侧带有特殊圆环状结构, 上唇裂片呈三角状且裂片高度明显低于轮盘的高度, 此时头部宽度大于其长度。咀嚼器为蛭态轮虫特有的枝型咀嚼器, 齿式为7/7, 还存在若干细长的肋条状小齿(图 3)。
2.2 不同生长状态的形态学变化
挑取样品中的环颈敖突轮虫进行各部位长度测量(n=10)。结果如下: 体长(331±41.8) μm, 头部长度(22.8±1.7) μm, 头部宽度(27.9±2.6) μm, 头冠宽度(38.9±2.6) μm, 颈长(54.5±5.9) μm, 足部长(22.7±3.0) μm, 刺戟长(8.4±0.9) μm。通过对比发现, 该轮虫在生长过程中, 虫体大小的改变主要取决于躯干部的长度, 头部和趾的长度在虫体整个生长过程中无明显变化。

图 2 环颈敖突轮虫背面观Fig. 2 Dorsal view of Otostephanos torquatus
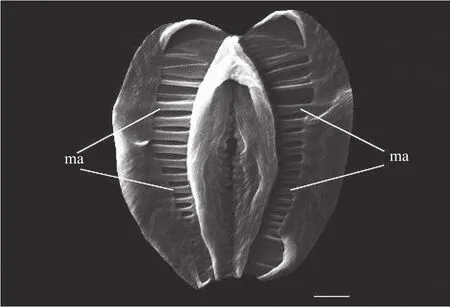
图 3 环颈敖突轮虫咀嚼器扫描电镜照片(ma. 咀嚼器齿式, 比例尺为2 μm)Fig. 3 Trophi of Otostephanos torquatus (ma. major teeth, scale bar=2 μm)
2.3 遗传距离和进化树分析
从GenBank中下载与环颈敖突轮虫COⅠ基因序列相似度高的物种的COⅠ基因序列, 通过MEGA7.0分析这些物种的遗传距离。由于Gen-Bank数据库中无环颈敖突轮虫的COⅠ基因序列,我国对同属的多纳敖突轮虫(Otostephanos donneri)的描述也仅限于形态特征, 选择基因库中同属的多纳敖突轮虫的COⅠ基因序列聚类分析, 以旋轮科的Pleuretra lineata及盘网轮科的游荡盘网轮虫(Adineta vaga)作为环颈敖突轮虫的外类群, 宿轮科宿轮属的Habrotrocha constricta作为同科物种,使用邻接法构建系统发育树, 聚类分析结果表明该轮虫的遗传位置属于蛭态亚纲、宿轮科、敖突轮属(图 4)。
3 讨论
目前蛭态轮虫的物种鉴定方法主要依靠形态学分类。咀嚼器作为蛭态轮虫的重要特征, 为种类鉴定提供了较全面的分类学依据[16]。不同蛭态轮虫咀嚼器的大小和齿的形式等方面都会存在明显差异。由于咀嚼器尺寸较小, 采用扫描电镜可以使咀嚼器的齿式以及细节特征更加清晰。在传统分类学的基础上, 通过对蛭态轮虫COⅠ基因的提取和序列分析, 为蛭态轮虫的物种鉴定提供更加可靠的分子生物学依据。
3.1 分类背景
全世界蛭态亚纲轮虫目前共记录4科19属461种[17], 我国共记录蛭态轮虫3科9属42种, 敖突轮属全球共记录15种, 我国仅记录1种[18]。对于蛭态轮虫的分类学地位, 我国一般将其归类于轮虫动物门、双巢纲、蛭态目[19]。根据最新的分类学研究,目前国际上将蛭态轮虫归类于蛭态亚纲[17,20,21]。相对于国外, 我国对蛭态类轮虫的分类和物种鉴定尚处于起步阶段。
3.2 敖突轮属特征及其与国内外相关物种的比较
轮盘上带有的环状结构为该属特有的分类特征, 且该属轮虫咀嚼器的齿式, 有助于食物在体内形成“食物泡”。咀嚼器的齿式是蛭态亚纲轮虫进行物种鉴定的重要分类特征。生活在不同生境的环颈敖突轮虫个体间咀嚼器齿的形式存在差异, 在我国分布的该种咀嚼器齿式为7/7, 相较于欧洲, 该种咀嚼器齿式有6/6、7/7、8/8三种形态[11]。在蛭态轮虫的形态学物种鉴定的基础上, 可以结合分子生物学的方法对轮虫进行基因测序, 从基因层面上进一步支持形态学鉴定结果。
我国敖突轮属的记录仅有一种, 即多纳敖突轮虫(Otostephanos donneriBartos, 1959)[18], 该种发现在我国云南省, 作者仅把多纳敖突轮虫作为轮虫群落结构的种类组成记录, 没有对其进行具体的形态学描述。环颈敖突轮虫和多纳敖突轮虫除了体内含有食物泡, 轮盘带有环状结构等共有特征外, 虫体大小长度相近, 上唇形状也极其相似。但在生活环境、咀嚼器形态、刺戟形态等方面都有明显差异。通过国外对多纳敖突轮虫的形态描述, 两个种的生境和形态对比如表 1所示。
环颈敖突轮虫在韩国和意大利地区也有记载[4,22],对比该轮虫在这三个地区的生存环境, 此物种均栖息在苔藓植物水膜中, 迄今, 未在其他环境中发现该物种。对比三个区域该种的形态结构特征, 未发现明显差异。通过对比我国与意大利和韩国的地理位置, 三个国家的经纬度跨度较大, 属于不同地理板块, 因此推测该轮虫在全球分布范围较广, 可以适应不同的气候类型和温度变化。对于某一种蛭态轮虫来说, 具有类似细菌和原生动物的全球分布特征, 这种高扩散能力, 对孤雌生殖的蛭态轮虫能够在地球上长期存在具有重要意义[23,24]。
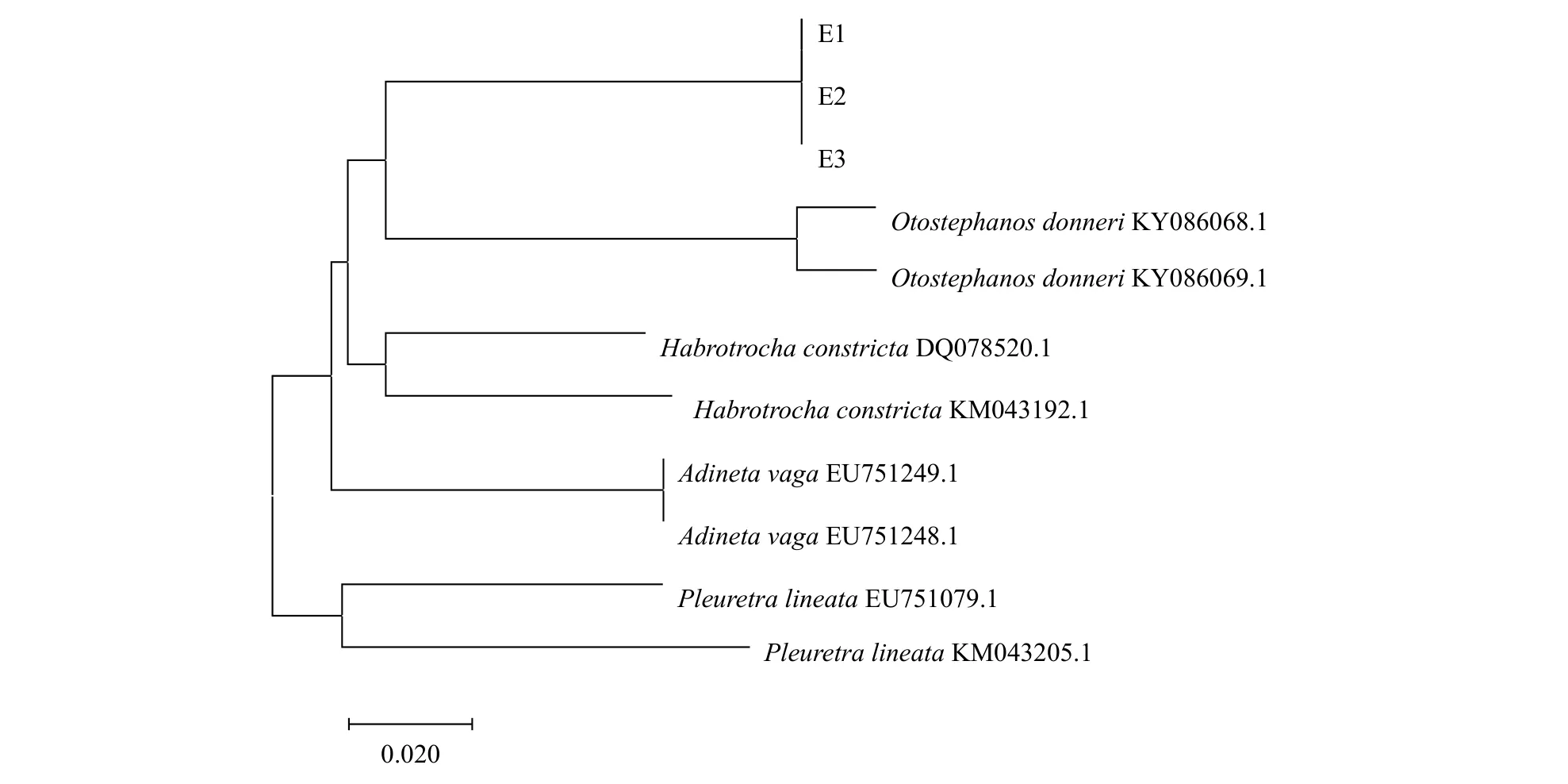
图 4 基于环颈敖突轮虫COⅠ基因序列构建的NJ系统发育树Fig. 4 The Neighbor-joining tree based on CO Ⅰgene sequences of Otostephanos torquatus

表 1 我国新记录种环颈敖突轮虫与已有记录多纳敖突轮虫的对比Tab. 1 Comparison of the newly recorded species Otostephanos torquatus and the existing record of O. donneri in China
3.3 展望
我国对蛭态轮虫的研究基本处于空白, 主要归结于三个方面的原因: (1)蛭态轮虫生境多样, 研究者对各种生境蛭态轮虫的提取方法不熟悉; (2)缺少蛭态轮虫物种鉴定的参考文献和书籍。目前对蛭态轮虫的种类鉴定主要参考德文版的《Ordnung Bdelloidea》[10], 语言限制性大; (3)蛭态轮虫无背甲, 易收缩, 活体状态下难以获得其形态学信息。随着科技水平的发展和研究手段的进步, 通过先进的摄影设备拍摄蛭态轮虫的形态特征, 使用电镜、DNA测序等多重手段可扩展蛭态轮虫的研究范围。
我国领土南北纬度跨越近50°, 高原、盆地、平原等地形复杂多样, 各地区蛭态轮虫多样性分布也会有所差异。蛭态轮虫在全球从南极到赤道各个纬度区域均有分布, 可以适应多样的生境变化[25]。我国具有独特的地理优势, 因此, 加强苔藓、水体、土壤和地衣等不同栖息地和不同生物地理格局的蛭态轮虫分类学研究非常重要。随着先进摄影设备、电镜技术和分子测序等多种技术方法的应用, 有望在我国发现更多的蛭态轮虫种类。
近年来, 随着分子生物学技术的迅速发展, 国际学者对蛭态轮虫的隐生行为和孤雌生殖领域的分子生物学研究有重大进展。从基因组层面分析低湿休眠中出现的耐干燥性等分子特征, 得出不同生境的蛭态轮虫在基因结构方面存在明显差异[26]。生态学方面, 不同种类的蛭态轮虫可以以整体形式作为小型水体营养状态的生物指标[27]。通过对大样本蛭态轮虫线粒体COⅠ基因的测序及构建系统发育树, 结果表明不同的蛭态轮虫具有不同的生态位和栖息地偏好[28]。因此在野外调查过程中, 调查不同生境, 获取更多具有栖息地偏好的物种, 对分子生物学和生物学等方面的进一步研究具有重要意义。
