人工无瓣海桑林下灌层分布格局及多样性研究
2020-03-26周元满苏浩毅王平
周元满,苏浩毅,王平
(广东海洋大学农学院,广东 湛江 524088)
红树林在保护海滨湿地生态系统和生物多样性以及维护海湾、河口地区生态平衡等方面均有着不可替代的重要作用,是海岸带植被的重要组成树种群落。红树植物无瓣海桑(Sonneratiaapetala)的引种扩大了红树林的种质资源,促进了红树林的恢复与发展。无瓣海桑原产孟加拉国,是红树林中优良乔木树种之一,分布于印度、孟加拉国、泰国、马来西亚、斯里兰卡等国盐度较低的泥质滩涂,群落分布于低潮线的外缘,具有速生、耐浸淹、抗逆性强等特性[1,2],1993年引至广东省湛江市沿海种植,并取得成功,成为雷州半岛人工恢复红树林的先锋树种之一[3,4]。 近几年,无瓣海桑的研究主要集中在自然更新、果实的化学成分、木材性质及防护效应等方面[5-10]。但由于无瓣海桑有一定的扩散能力,是否会引起生物入侵等生态学问题值得关注。
林下植被是人工林生态系统的一个重要组成部分,也是林分生产力指标之一,植物种群分布格局是植物种群在群落中所处的空间结构可定量化描述的基本特征,是种群自身特性、种间关系及环境条件综合反映,也是影响种群发展的重要因素。目前,无瓣海桑是否可能成为沿海红树林生态系统的入侵种仍未有定论,为了更好地实施人工引进,加强红树林的防风削浪的作用,促进红树林生态系统的稳定,本文以雷州附城滩涂的人工无瓣海桑林为对象,研究无瓣海桑对当地乡土红树植物白骨壤(Avicenniamarina)、桐花树(Aegicerascorniculatum)、木榄(Bruguieragymnorhiza) 和秋茄(Kandeliacande)生长分布格局及多样性的影响,以期为深入了解其生态适应性(生态位) 、合理推广利用该树种、保护红树植物物种多样性以及红树林湿地生态系统的可持续发展提供理论依据。
1 研究地概况
研究地位于湛江市雷州附城镇,雷州半岛位于中国大陆最南端,属北热带海洋季风区,东临中国南海,南隔琼州海峡与海南岛隔海相望,西临北部湾, 西北与广西壮族自治区相连。地处109°31′—110°55′ E、20°12′—21°35′ N的热带、亚热带过渡地区。年最冷月平均气温15.5 ℃ ,年平均气温23.0 ℃ ,极端最高气温38 ℃ ,极端最低气温0 ℃,年均水温为25 ℃ ,2月约20 ℃ ,8月约28 ℃ ,且不随气温的急剧升降而起相应的变化,形成华南沿海红树林生长的良好水热条件。年降水量1 400~2 000 mm,年蒸发量1 800 mm,有明显的干湿季,7—8月为台风频繁的湿润季节,属于多台风多雷地区,潮汐为混合全日潮,平均潮差2.53 m,最大潮差6.25 m,高潮时水深可达2.5 m[4]。试验地林下植被主要有白骨壤、桐花树、秋茄、木榄和红海榄。
2 研究方法
2.1 样地设置与调查
基于无瓣海桑林内、林缘及光滩处各红树植物的更新状况不同,本研究按照目标树中心样圆法设置随机样地[11]。在土角村和芙蓉村的无瓣海桑人工林(分别于2002、1997年种植,文中分别记为无瓣海桑Ⅰ,无瓣海桑Ⅱ)中选择具有代表性的林分,设置400 m2的样地,在样地内按照克拉夫特分级法[12,13]分别在林缘及林内选择亚优势木和中等木5~8株作为目标树,实测并记录目标树的树高、胸径、冠幅等测树指标。以林内及林缘的目标树为圆心,分别以2 m、4 m和6 m为半径设置样圆,将样圆依次编号为C1、C2、……Cn,统计样圆内所有无瓣海桑、林下各种灌木的数量、盖度、分布情况等以及无瓣海桑幼树幼苗(高度≤1 m)的数量及株高。样地及样圆特征值见表1。

表1 样地类型及特征值
2.2 物种多样性
物种多样性能够体现出生物群落和生态系统的结构复杂性,多样性指数越高,群落越趋于稳定。物种多样性测度指标选用物种丰富度指数(Margalef指数)、物种多样性指数(Simpson指数)和物种均匀度指数(Pielou指数)。计算公式[14]如下:
(1)Simpson多样性指数
(1)
(2)Margalef丰富度指数
(2)
(3)Pielou均匀度指数
(3)
各计算式中的N为样方中所有物种的个体总数,ni为第i个种的个体数,S为样方中的物种数。
采用SPSS软件进行数据统计与分析,采用DPS数据处理系统进行多样性指数分析[15]。
2.3 种群分布格局
植物种群分布格局是植物种群在群落中所处的空间结构可定量化描述的基本特征。本文引用泊松分布对样地种群分布格局进行分析,种群分布格局测定按方差(V)/ 均值m比值的t检验方法,方差(V)/均值(m)比率是一种较好的表征种群个体的水平空间分布特征的指标,其结论不存在难以判断的临界点,分析结果具有相对的客观性。计算公式[16,17]如下:
(4)
(5)
C=V/m
(6)

(7)
最后以N-1的自由度和95%的置信度检验。
3 结果与分析
3.1 无瓣海桑林下灌层多样性分析
物种多样性是群落生态组织水平独特的可测定的特征之一[18],是度量一个群落结构和功能复杂性的指标,对物种多样性的研究可以更好地认识群落的组成、变化和发展[19,20]。雷州附城无瓣海桑林下灌木种类在以样木为中心的不同范围内的组成及数量结构见表2和表3。通过不同类型无瓣海桑林下灌层多样性调查结果(表4)可见,各类圆形样方中物种的Pielou均匀度指数和Simpson指数均没有显著差异,而Margalef丰富度指数在各样圆中的差异显著(P<0.05),无瓣海桑样地Ⅰ中以样木为中心样圆的丰富度指数强度变化为C4>C3>C1>C2,无瓣海桑样地Ⅱ中丰富度指数强度变化为C10>C9>C7>C8,可见无瓣海桑林缘灌木层物种比林内更加丰富,无瓣海桑林分的林缘且与林缘样木中心距离最大的样方可以适应更多的物种生存,稳定性较高。从表4还可以看出,样地Ⅱ的物种均匀度比样地Ⅰ的物种均匀度高,丰富度指数较大,Simpson多样性指数无明显差异。而产生这种差异的原因可能是样地Ⅱ林龄较大,森林群落结构相对稳定。而样地Ⅰ的林龄小,丰富度较小是因为无瓣海桑引种时间长,根系和枝条旺盛,吸收大量的养分,使一些适应性较差的物种无法继续生存,林下灌木的分布多呈簇生状分布,均匀度指数比样地Ⅱ较小,该林分仍处于竞争阶段。

表2 样地Ⅰ物种分布盖度 %

表3 样地Ⅱ物种分布盖度 %
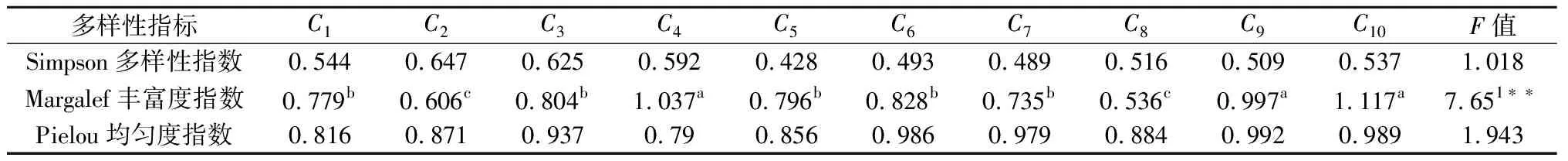
表4 不同圆形样地物种多样性及方差分析结果(P = 0.05)
注:*表示P< 0.05;**表示P<0.01;不同字母表示差异显著(P=0.05)。F0.05=2.39,F0.01=3.46
3.2 无瓣海桑林下灌层的分布格局
3.2.1 无瓣海桑林下灌层的分布规律及检验 从表5可见,白骨壤、红海榄、秋茄、桐花树和木榄的方差均值比例大于1,经T检验为T>T0.05,分布格局均为集群分布,集中强度为木榄>桐花树>秋茄>白骨壤>红海榄,主要原因是秋茄、桐花树及木榄等植物的胎生现象,种子在离开母株前萌发,胚轴细长,成熟后掉落在母株四周,从而造成它们的分布格局趋向集群分布这种状况。与同处于英罗湾的广西红树林上述5 种红树植物种群全部为随机分布的情况[21]有明显差异,同时表明研究区的无瓣海桑发育处于竞争阶段,因为随着年龄的增长,种群的分布格局将从集群分布趋向随机分布。梁士楚[22]研究表明,英罗湾的红海榄种群幼苗群落、小树群和老树群在其发展过程中分布格局的动态变化相应为集群分布→随机分布。

表5 无瓣海桑林下植被的泊松分布方差均值比例及T检验结果
3.2.2 不同类型无瓣海桑林下植被的分布格局 表6表明,样地Ⅰ中,白骨壤趋近于随机分布,秋茄和红海榄为集群分布。因为样地Ⅰ地处低潮位,而白骨壤是天然红树林在沿海滩涂前缘的先锋树种,对周期性潮汐淹浸深度具有很强的适应能力。在样地Ⅱ中,秋茄、桐花树和木榄均为集群分布,而且聚集强度较大,在两类样地中,秋茄的聚集强度明显不同,主要是因为样地Ⅱ的潮位比样地Ⅰ要高,风浪相对较小,再者由于秋茄和桐花树生态位宽度值很接近,而且桐花树生态位宽度值略大,生态位重叠值也较高,所以会出现两者相对较激烈的资源竞争[23],因此在样地Ⅱ中两者都适合大量繁殖,而桐花树会占据部分资源,以至样地Ⅱ的秋茄的聚集度较小。

表6 泊松分布方差均值比例及T检验结果
4 结论与讨论
4.1 物种多样性反映了生物群落在组成、结构及功能等方面的异质性[24],体现了群落结构类型、发展阶段及其稳定程度与生境差异。无瓣海桑林下各种灌木在以样木为中心的不同范围内分布盖度不同,不同样圆的无瓣海桑林下物种的均匀度和Simpson指数无明显差异,丰富度指数达显著差异(P<0.05),无瓣海桑林缘灌木层物种多样性指数比林内更高,可以适应更多的物种生存,稳定性较高。
4.2 无瓣海桑人工林下乡土红树植物白骨壤、红海榄、秋茄、桐花树和木榄呈现集群分布,且集中强度为木榄>桐花树>秋茄>白骨壤>红海榄;同一物种在不同年龄及潮位的无瓣海桑林下的分布格局不同,样地Ⅰ中,红海榄和秋茄呈现集群分布,白骨壤呈现随机分布,样地Ⅱ中,秋茄、桐花树和木榄均呈现集群分布;秋茄在两类样地中的聚集强度明显不同,可能与秋茄和桐花生态位宽度值很接近(桐花树略大),生态位重叠值也较高,导致两者在资源竞争中桐花树会占据相对优势,以致样地Ⅱ的秋茄的聚集度较小。
4.3 在研究地未发现无瓣海桑幼苗,而无瓣海桑林下的乡土红树植物如红海榄、木榄和秋茄等幼苗均分布在林内及林缘岸边均有不同程度的聚集,造成这种情况的原因可能是种子落地后,有的留在母株周围,而有的则经海浪冲到岸边。风浪大小是红树植物能否存活的重要生态影响因子,而无瓣海桑冠幅大,树体高,在红树林中的盖度也大,因此它在林中阻挡了风浪,使风浪到达海岸边时的冲击力减小,从而促进乡土红树植物幼苗的天然更新。
4.4 在研究区两种类型的无瓣海桑样地中均未发现无瓣海桑幼苗,可能是因为无瓣海桑为阳性树种,林下乡土红树植物郁闭成林,林内缺乏光照,其种子成熟后难以在这样荫蔽的林内大量萌发形成天然更新[25],因此无瓣海桑在林内难以形成幼苗的天然更新,也不具有向其他群落竞争扩张的优势,对乡土红树植物的幼苗天然更新和生长没有抑制作用,并且由于无瓣海桑具有速生、耐浸淹、抗逆性强等特性,在生长过程中能促进滩涂泥沙的淤积,使滩涂高程增大,加上无瓣海桑呼吸根密集,能消减波浪和滞留乡土红树植物繁殖体的作用[26],反而更有利于当地乡土红树植物的生长,并为乡土红树植物的天然更新和生长创造有利条件,在沿海红树林生态系统经营中可考虑以乡土树种为主,引进种与乡土种相结合的模式进行人工林恢复建设,但对无瓣海桑生态影响仍需进一步跟踪监测,以便为红树林的生态恢复提供参考。
