丹酚酸B对高糖诱导心肌成纤维细胞增殖及转分化的作用研究
2020-03-20隋海娟符丽娟
王 娜,隋海娟,符丽娟
(1.锦州医科大学基础医学院药理学教研室,辽宁 锦州 121000; 2.朝阳市第二医院药剂科,辽宁 朝阳 122000)
糖尿病心肌病是糖尿病患者主要的心脏并发症,心肌间质纤维化为其主要病理改变,也是引起糖尿病心室重塑导致心力衰竭的关键原因;其表现为心肌间质细胞增殖及表型转分化,细胞外基质(extracellular matrix,ECM)过度沉积等。Wnt/β-catenin信号通路是参与细胞生长、增殖的重要途径,与肺、肝脏及肾脏等多脏器纤维增生性疾病的发生密切相关[1-3];Wnt/β-catenin通路的激活也参与了糖尿病心脏重构的病理过程[4]。目前,针对Wnt/β-catenin通路抑制剂的研究已成为防治纤维化相关疾病药物研究的重要靶点。丹酚酸 B(salvianolic acid B,Sal B)是用于心血管系统疾病的传统中药丹参的主要活性成分,基础研究发现[5,6]其对多种心血管损伤具有一定保护作用,如心肌缺血/再灌注损伤、2型糖尿病小鼠心肌病变等。但有关Sal B对糖尿病心肌纤维化的具体作用目前尚不明确,本研究通过体外培养心肌成纤维细胞(cardiac fibroblasts,CFs),高糖诱导模拟糖尿病高糖状态,基于Wnt/β-catenin信号通路,观察Sal B对高糖诱导CFs增殖及转分化的影响,探讨Sal B对糖尿病心肌间质纤维化的作用及可能机制,为传统中药防治糖尿病心肌病变提供理论依据。
1 材料与方法
1.1 材料
1.1.1动物、药品与试剂 1~3 d龄新生SD大鼠乳鼠(锦州医科大学实验动物中心提供),动物使用许可证号:SCXK(辽)2016-0004。Sal B(美仑MB6598);Wnt/β-catenin通路抑制剂XAV939(APEX BIO A1877)。DMEM培养基(Mediatech公司);增强型CCK-8试剂盒(上海尚宝生物科技有限公司);α-SMA抗体(博士德公司WL02509);β-catenin抗体(abcam公司ab32572);p-GSK 3β 抗体(abcam公司ARG51777)。
1.1.2仪器 CO2孵箱(三洋公司);相差显微镜(Olympus公司); Mini-REPOTEANⅡ型电泳槽、电泳仪、Semi-dry Transfer cell(Bio-Red公司);TGL-16G高速冷冻离心机(日立公司);Biocell 2010酶标仪(Anthos Labtec Instruments公司);数显气浴恒温震荡器(江苏CHA-SA);化学发光凝胶系统分析仪(美国UVP公司);Leica 4000B正置荧光显微镜(德国Leica公司)。
1.2 方法
1.2.1CFs分离与培养 新生SD大鼠乳鼠,♀ ♂不拘,体积分数75%的乙醇消毒,取出心脏,冷PBS冲洗2次,放于冷培养基中,剪约1 mm3组织碎块,加入0.25% 胰酶37 ℃水浴中进行消化。将消化的上清加至含10%胎牛血清的培养基中终止消化。将收集的细胞1 000×g离心10 min,弃去上清。向沉淀中加入含10%胎牛血清的培养基,吹打均匀,制成细胞悬液。剩余组织继续按上述操作消化至组织消化完全,之后将细胞悬液均匀接种于培养瓶中,放入37 ℃、5% CO2孵箱。差速贴壁90 min后,弃去培养液,PBS冲洗细胞3次,更换新的培养基。细胞生长近融合状态时0.25% 胰酶消化传代,本实验使用2~3代细胞。
1.2.2CCK-8检测细胞增殖 将对数生长期的CFs胰酶消化后,用培养液制成单个细胞悬液计数,以每孔300个细胞接种至96孔板,每孔100 μL,贴壁培养24 h,吸去上清,加无血清培养基,血清饥饿24 h。分别进行:① 不同浓度葡萄糖对CFs增殖的作用:分为正常对照组(Control,含5.5 mmol·L-1葡萄糖)和葡萄糖组(含10、20、25、30、50 mmol·L-1葡萄糖);② 高糖诱导 CFs增殖的时间效应:分为 Control组,高糖组(HG组,含25 mmol·L-1葡萄糖),作用时间分为 12 h、 24 h、48 h;③Sal B对高糖诱导 CFs增殖的作用:分为 Control组、HG组(25 mmol·L-1)、HG+ Sal B(12.5、25、50 μmol·L-1)、高渗透压对照组 (MG组,含 5.5 mmol·L-1葡萄糖+19.5 mmol·L-1甘露醇)。按上述不同的处理因素作用48 h,每组设 6复孔。每孔加入CCK-8试剂10 μL,轻轻摇晃96 孔板后放入37 ℃ 5% CO2培养箱中继续培养1 h,取出96孔板置于酶标仪上,490 nm测定各孔吸光度OD值。
1.2.3分组及药物处理 实验分为:① Control组(含5.5 mmol·L-1葡萄糖),②MG组(含5.5 mmol·L-1葡萄糖+19.5 mmol·L-1甘露醇),③ HG组 (含25 mmol·L-1葡萄糖),④ HG + Sal B组(25 μmol·L-1Sal B预处理细胞1 h,加25 mmol·L-1葡萄糖),⑤ HG+XAV939组(1 μmol·L-1XAV939预处理细胞 1 h,加25 mmol·L-1葡萄糖),⑥ XAV939组(1 μmol·L-1),⑦ Sal B组(25 μmol·L-1)。
1.2.4苦味酸天狼猩红染色 取出药物作用后的24 孔培养板,弃掉培养基, PBS 清洗3次,4%多聚甲醛室温固定15 min。PBS清洗3次,0.5% 的聚乙二醇辛基苯基醚(Triton-X-100)破膜20 min,PBS 清洗3 次,天狼猩红染液染色1 h,苏木素染液复染细胞核5 min,倒置显微镜下观察并拍照。
1.2.5免疫荧光法 各组CFs加不同条件培养基药物处理24 h后,吸去培养液,冷PBS 漂洗3次;新鲜配制4% 多聚甲醛室温固定30 min,PBS 漂洗3次;0.3% Triton X-100作用30 min,PBS漂洗3 次;室温山羊血清工作液封闭60 min;滴加α-SMA一抗溶液,4 ℃孵育过夜;PBS漂洗,异硫氰酸荧光素(FITC)标记的二抗室温孵育1 h,PBS漂洗,用含DAPI的抗淬灭封片剂封片,荧光显微镜观察及拍照。
1.2.6Western blot 各组CFs加不同条件培养基药物处理24 h后,冷PBS冲洗3次,立即放入预冷的裂解液中,每隔5 min震荡20 s,4 ℃ 12 000×g离心,取上清,BCA法进行蛋白质定量。每个泳道蛋白上样量为20 μg,行 SDS-PAGE凝胶电泳,观察Marker移动情况。电泳后将凝胶中的蛋白质转移至硝酸纤维素膜上,室温封闭1 h,分别加兔抗大鼠α-SMA一抗、兔抗大鼠 β-catenin一抗、兔抗大鼠p-GSK 3β 一抗、兔抗大鼠GSK 3β 一抗,4 ℃孵育过夜。次日早上取出膜,TBST洗膜3次,每次10 min,将膜放入山羊抗兔二抗 (1 ∶1 000)中,室温孵育1 h。TBST洗膜3次,每次10 min,行ECL化学发光显影。利用 Visionworks 6.3.3图像采集及分析软件对蛋白带进行分析。实验重复3次。
1.2.7统计学方法 实验数据用表示,SPSS 18.0软件进行统计学分析。采用One-way ANOVA和LSD-t检验进行两两比较。
2 结果
2.1 不同浓度葡萄糖对CFs增殖的作用CCK-8结果(Tab 1)显示,不同浓度(5.5、10、20、25、30、50 mmol·L-1)葡萄糖诱导CFs 24 h,25、30、50 mmol·L-1葡萄糖诱导,CFs增殖率明显增加,实验中25 mmol·L-1高糖诱导时细胞增殖率已经达到196 %(P<0.01),本实验确定高糖诱导浓度为25 mmol·L-1。
Tab 1 Effects of different concentrations of glucoseon
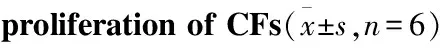

GroupConcentration/mmol·L-1OD490Proliferationrate/%Control5.50.276±0.016100.00HG100.278±0.020100.79200.291±0.038105.50250.541±0.277∗∗196.07300.572±0.226∗∗207.56500.571±0.321∗∗207.25
**P<0.01vscontrol
2.2 高糖诱导CFs增殖时间效应根据Tab 1结果,应用25 mmol·L-1葡萄糖作用12、24及48 h,结果(Tab 2)显示,作用24 h时CFs增殖率最高,达到204.6%(P<0.01),本实验确定高糖作用时间为24 h。
Tab 2 Time effect of high glucose on proliferation
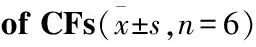

GroupTime/hOD490Proliferationrate/%Control120.267±0.026100.00HG120.294±0.022110.11Control240.304±0.019100.00HG240.623±0.019∗∗204.60Control480.397±0.027100.00HG480.503±0.044∗∗126.80
**P<0.01vscontrol
2.3 不同浓度Sal B对CFs增殖的影响CCK-8结果(Tab 3)显示,与Control组相比,MG组CFs无明显变化,提示高渗透压对CFs增殖无明显影响,而25 mmol·L-1高糖作用可促进CFs增殖(P<0.01);与HG组相比,经不同剂量(12.5、25、50 μmol·L-1)Sal B 干预,对CFs增殖均出现抑制作用(P<0.05),25 μmol·L-1差异具有极显著性(P<0.01),后续实验选取Sal B的给药浓度为25 μmol·L-1。
2.4 Sal B对高糖诱导CFs胶原纤维表达的影响天狼猩红染色(Fig 1)显示:倒置显微镜下,胶原纤维为粉红色,细胞核为紫红色。与Control组相比,HG组细胞数量增多,红色胶原面积明显增加。与HG组相比,经Sal B以及XAV939预处理后,红色胶原面积减少,细胞间隙变大;单独应用Sal B及XAV939对CFs胶原纤维表达无明显影响。表明Sal B可通过抑制Wnt/β-catenin信号途径减少高糖诱导CFs胶原沉积。
Tab 3 Effects of different concentrations of Sal B


GroupConcentration/mmol·L-1OD490Proliferationrate/%Control0.307±0.021100.00MG0.306±0.02999.78HG0.595±0.010∗∗193.71HG+Sal B12.50.560±0.033#182.15250.374±0.023##121.60500.363±0.009##118.18
**P<0.01vscontrol;#P<0.05,##P<0.01vsHG
2.5 Sal B对高糖诱导CFsα-SMA表达的影响免疫荧光(Fig 2)显示:红色荧光代表α-SMA,蓝色荧光为细胞核,与Control组相比,HG组细胞红色荧光明显增强;与HG组相比,经Sal B(25 μmol·L-1)以及XAV939预处理后,红色荧光强度明显减弱;单独Sal B组及单独XAV939组与Control组相比荧光强度变化不明显。
Western blot(Fig 3)显示:与Control组相比,HG组α-SMA蛋白表达明显增加(P<0.01);与HG组相比,HG+XAV939组与HG+Sal B组α-SMA蛋白水平降低(P<0.01),HG+XAV939组降低更为明显;单独Sal B组及单独XAV939组与Control组相比无明显变化。α-SMA为肌成纤维细胞的标志性蛋白,提示高糖可诱导CFs转分化为肌成纤维细胞,

Fig 1 Effects of Sal B on expression of collagen fiber in CFs induced by high glucose(400×)
A:Control group;B:HG group; C:HG + Sal B group; D:HG + XAV939 group; E:XAV939 group; F:Sal B group

Fig 2 Effect of Sal B on expression of α-SMA in CFs induced by high glucose(400 ×)
Sal B以及抑制Wnt/β-catenin信号可阻止高糖诱导CFs转分化程度。
2.6 Sal B对高糖诱导CFs β-catenin、 p-GSK 3β表达影响Western blot(Fig3)显示:与Control 组相比,HG组β-catenin、p-GSK 3β蛋白表达明显增加(P<0.01);与HG组相比,HG+XAV939组与HG+Sal B组β-catenin、p-GSK 3β蛋白水平降低(P<0.01),HG+XAV939 组降低更为明显;单独Sal B组及单独XAV939组与Control组相比无明显变化。表明Sal B可通过下调 β-catenin、 p-GSK 3β表达抑制高糖诱导CFs增殖及转分化程度。

Fig 3 Effects of Sal B on expression of α-SMA,β-catenin
1:Control group; 2:HG group; 3:HG + Sal B group;4:HG + XAV939 group; 5:XAV939 group;6:Sal B group;**P<0.01vsControl group;##P<0.01vsHG group
3 讨论
CFs是心肌间质的主要细胞成分,糖尿病状态下CFs增殖可合成大量胶原,ECM合成增加,导致心肌间质纤维化。然而,单纯CFs增殖不足以完成纤维化进程,CFs转分化(即CFs转分化为肌成纤维细胞)则是心肌间质纤维化进展的关键所在,转分化作用可导致ECM合成级联放大,从而加重心肌间质纤维化及心脏重构的病理进程[7]。α-SMA是肌成纤维细胞特征性标志物,本研究中应用高糖(25 mmol·L-1葡萄糖)诱导,CFs增殖率显著升高,α-SMA表达增加,胶原纤维增多;证实高糖可诱导CFs增殖转分化为肌成纤维细胞,特征性α-SMA表达增加,与Tao等[8]报道一致。实验中经 Sal B(25 μmol·L-1)预处理后,明显抑制了CFs增殖及胶原纤维沉积,α-SMA表达明显减少,提示Sal B对高糖诱导CFs增殖及转分化有一定的抑制作用。
Wnt/β-catenin信号通路是调控组织器官发育的重要途径,参与细胞增殖、迁移及间质转化等过程。其中β-catenin在细胞内的数量与状态对该途径起决定作用,β-catenin在胞质中积累后进入细胞核,是Wnt信号传递的重要环节,β-catenin异常激活参与纤维增生疾病的发生发展[9]。刘理静等[1]研究显示,高糖可通过激活β-catenin引起肺成纤维细胞增殖及胶原合成。也有研究显示,持续血糖可通过激活Wnt/β-catenin/TCF7L2通路,使β-catenin活化参与糖尿病心肌病的病理生理过程[4,10]。本研究中,高糖(25 mmol·L-1葡萄糖)诱导,CFs内β-catenin表达明显增加,经Sal B及XAV939预处理后,β-catenin表达明显减少。提示Sal B可抑制高糖诱导CFs增殖转分化,其作用与下调β-catenin表达有关。
正常情况下,β-catenin受多种蛋白调控, GSK 3β为其负向调节因素。当无 Wnt 配体时,GSK 3β的调控作用使胞质内β-catenin处于较低水平;当有Wnt配体时,Wnt配体与膜上Frizzled及LRP5/6受体结合,形成复合物,激活胞内的 Dsh,从而抑制GSK 3β活性,阻止胞质内β-catenin的降解;当GSK 3β的N端 9位丝氨酸被磷酸化后,p-GSK 3β增加,降低了GSK 3β活性,从而导致胞质β-catenin过度表达[11]。有学者在小鼠肾间质纤维化的研究中发现,p-GSK 3β水平与 β-catenin 表达同步增加[12];Liu等[13]发现, Wnt/β-catenin 信号通路在糖尿病诱导的心肌损伤中被激活,推测其与糖尿病诱导的心肌病变的发生有关。本研究中,高糖诱导CFs内p-GSK 3β 与 β-catenin亦出现同步增加;经Sal B及Wnt/β-catenin通路抑制剂XAV939预处理后,p-GSK 3β、β-catenin表达出现同步下调;提示GSK 3β磷酸化,可上调β-catenin表达,使α-SMA表达增多,促进高糖诱导CFs增殖及转分化;Sal B可通过下调p-GSK 3β、β-catenin表达抑制高糖诱导CFs细胞增殖及转分化过程。
综上所述,高糖作用CFs可上调p-GSK 3β与β-catenin表达,促进CFs增殖、转分化;Sal B可有效抑制高糖诱导CFs增殖及转分化作用,其机制与抑制Wnt/β-catenin信号通路有关。
