褐牙鲆白细胞介素21基因克隆及表达分析
2020-03-20薛婷婷郑津辉潘宝平孙金生
薛婷婷,郑津辉,周 密,潘宝平,孙金生,高 虹
( 天津师范大学 生命科学学院,天津市动植物抗性重点实验室,天津 300387 )
褐牙鲆(Paralichthysolivaceus)又称比目鱼,属鲽形目、牙鲆科、牙鲆属。近年来,随着养殖规模和养殖密度的大幅度提升,养殖环境日趋恶化,其在养殖的过程中极易受到病毒、细菌等多种病原体的侵染。迟钝爱德华氏菌(Edwardsiellatarda)属革兰氏阴性菌,不仅能使较多经济鱼类患病,给水产养殖业造成严重经济损失,而且也是一种人、鱼共患的病原菌[1-2],直接威胁人类健康,目前尚未创建针对迟钝爱德华氏菌的商业化防治措施。因此,全面深入的探讨研究水产鱼类关于该病原菌免疫应答的相关分子机制,将为建立一种有效、能够预防和控制迟钝爱德华氏菌感染的免疫防治新措施提供理论基础[3]。
白细胞介素21(IL-21)是近几年被发现并引起广泛关注的一个细胞因子,白细胞介素21大部分来自活化的CD4+ T、NKT细胞,和白细胞介素2、白细胞介素4以及白细胞介素15具有高度同源性,广泛分布在T细胞、B细胞、巨噬细胞和角质化细胞等一系列细胞外部。白细胞介素21受体广泛分布表达于各种器官(心、肝、脾、头肾、肌、肠)的细胞表面,细胞表面的受体结合后可以分泌细胞因子、炎症因子和趋化因子等,可以参与多重机体免疫调节[4-8]。目前,对白细胞介素21的免疫生物学功能、受体及相关的信号通路研究主要集中在哺乳动物[9-19],非哺乳动物中白细胞介素21的研究很少,因此,研究鱼类白细胞介素21对自身免疫的作用尤为重要。近年来,项黎新等[20-21]先后利用比较基因组学等方法,在红鳍东方鲀(Takifugurubripes)中鉴定了白细胞介素2和白细胞介素21的同源基因,表明在生物进化中,白细胞介素2细胞因子家族在鱼类中就已开始出现,王惠菊[22]研究发现,白细胞介素21基因在黑青斑河鲀(Tetraodonnigroviridis)、红鳍东方鲀以及人类中具有保守的共线性。
本研究以褐牙鲆及迟钝爱德华氏菌作为研究材料,探讨鱼类免疫因子白细胞介素21基因应答迟钝爱德华氏菌入侵的免疫机制,以加深对鱼类免疫系统调节机制的认识。
1 材料与方法
1.1 材料
1.1.1 试验鱼
健康褐牙鲆购于天津市滨海新区某水产养殖场,挑选生长发育状态优良的褐牙鲆6尾(试验组、对照组每组分别各3尾,设3个平行对照),体长(10±2) cm,选择可供氧、控温和水过滤的水循环鱼缸,盐度17~18,水温约20 ℃,充氧处理7 d,使褐牙鲆充分适应环境,并对鱼缸进行遮光处理,日投喂1次[1-2,23]。
1.1.2 病原体
试验所用迟钝爱德华氏菌,由天津市水产养殖病害防治中心提供,本实验室保存。将保存的菌种接种于LB液体培养基上,恒温振荡培养至菌液600 nm吸光值(OD600)为1.0。磷酸缓冲盐溶液离心洗涤菌体3次,将菌液密度调至1×107个/mL。
1.2 方法
1.2.1 RNA的提取和纯化
取急冻好的组织置于提前用液氮冷却完成的玻璃匀浆器中,滴加1 mL Trizol,进行充分研磨后,按照Trizol(Invitrogen)说明书中提供的方法提取总RNA。Na-nodrop检测总RNA纯度,琼脂糖凝胶电泳检验完整度,将提取的RNA存放于冰箱-80 ℃备用。
1.2.2 cDNA第一链的合成
购于Sangon Biotech®公司的cDNA 试剂盒,按照其推荐的方法,用于cDNA第一链的合成。
1.2.3 褐牙鲆头肾组织转录组测序
转录组测序由北京诺禾致源生物信息科技有限公司提供技术服务。测序平台为Illumina HiSeqTM2500。
1.2.4 IL-21基因编码区序列的克隆
根据褐牙鲆转录组测序结果得到褐牙鲆白细胞介素21类似cDNA基因序列,对该序列进行阅读框分析后,设计引物,利用PCR方法对蛋白质编码区域的基因序列进行克隆。引物见表1中的IL21-R和IL21-F。PCR扩增反应程序:94 ℃ 3 min;94 ℃ 30 s、60 ℃ 30 s、72 ℃ 30 s,35个循环;72 ℃ 10 min[24-29]。扩增的目的片段用试剂盒回收后,连接到pMD18-T 载体上,转化大肠杆菌(Escherichiacoli)DH5α感受态细胞,于37 ℃培养过夜。进行蓝白斑筛选和菌液PCR鉴定[30],将阳性克隆扩增产物送北京诺禾致源生物信息科技有限公司进行测序。
1.2.5 生物信息学分析
利用ORF软件 (http:∥www.ncbi.nlm.nih. gov/gorf/gorf.html)查找目的基因的开放读码框;利用BLAST(http:∥blast.ncbi.nlm.nih.gov/Blast.cgi)、ClustalX 及DNAstar中的MegAlign等软件完成序列的比对及生物信息学分析;使用Compute pI-Mw (http:∥web.expasy.org/compute_pi/)计算蛋白质等电点和相对分子质量;使用SMART程序分析氨基酸的结构,SignalP 4.1 Server进行信号肽的预测。使用Mega 5进行系统发育树的构建。
1.2.6 迟钝爱德华氏菌脂多糖提取
将90%苯酚溶液于66 ℃水浴预热10 min,加入到10 mL密度为1×107个/mL的迟钝爱德华氏菌菌液中,室温下搅拌25 min并冷却,放入冰箱冻存一晚,翌日取出,5000 r/min离心15 min,吸取上清液保留。剩下的残渣和酚层加入10 mL水,66 ℃水浴加热30 min,离心,吸取上清液。将两次吸取的上清液加入到0.85%的氯化钠溶液中进行透析,得到粗脂多糖,将得到的粗脂多糖用聚乙二醇提纯[31]。
1.2.7 褐牙鲆免疫
将购买的同一批生长状态、规格一致的健康褐牙鲆[体长(10±2) cm],随机分到两组鱼缸,第一组每尾腹腔注射100 μL提取的迟钝爱德华氏菌脂多糖,第二组作为对照组,每尾腹腔注射磷酸缓冲盐溶液100 μL[32]。每组鱼在各时间点随机抽取3尾进行解剖并分离各组织细胞,将每3尾鱼的相同组织混拌均匀,置于同一冻存管中保存备用。
1.2.8 基因表达相对定量分析
使用ABI 7500荧光PCR仪,使用 Promega®公司的GoTaq®qPCR Master Mix试剂盒推荐的方法,以反转录的cDNA为模板,β-actin为内参基因,进行荧光PCR反应,获得各组织的免疫相关基因的相对定量表达数据[所用引物见表1中的IL21-F-q和IL21-R-q;MyD88-F-q(MyD88为髓样分化因子),MyD88-R-q和β-actin-F-q,β-actin-R-q]。反应体系(24 μL):cDNA模版4 μL,上游引物(10 μmol/L) 0.4 μL,下游引物(10 μmo l/L) 0.4 μL,GoTaq®qPCR Master Mix 10 μL,Nuclease-Free Water 9.2 μL。各组织样品设3个平行,用2ΔΔCt的方法进行相对定量分析,用Origin 8软件作柱状图分析。
1.2.9 褐牙鲆头肾淋巴细胞分离
市场购买的健康褐牙鲆,体质量约1 kg,体长约45 cm,使用手术剪断颈处死,于超净台上解剖并分离头肾组织。将一个200目的无菌不锈钢筛置于含L-15培养基的培养皿上,培养基内含10%小牛血清、肝素10 U/mL、青霉素100 μg /mL及链霉素100 U /mL。用玻璃注射器针芯将头肾组织在不锈钢筛上轻轻研磨直至成细胞悬液,再用针头吹打悬液,使细胞分散为单个细胞。将所得头肾单细胞悬液小心铺于密度为1.0770 g/cm3的Ficoll分离液上,1500 r/min水平离心30 min,得到4层溶液,用针头小心将第二层的淋巴细胞吸出,细胞用培养液离心洗涤3次,台盼蓝染色后计数活细胞,调节细胞密度为105个/mL,分装于96孔板上,每孔100 μL,置CO2培养箱内25 ℃恒温培养[30]。

表1 试验所用引物序列Tab.1 Sequences of the primers used in qRT-PCR experiments
1.2.10 RNA干扰试验
用于沉默MyD88基因的siRNA (小干扰RNA,CCAGAGCGACTTTGAATTT)由广东锐博生物有限公司设计并合成。将100 μL褐牙鲆头肾淋巴细胞加入96孔培养板中,将0.25 μL siRNA的稀释液和1.6 μL转染试剂稀释液混合均匀后加入每孔中(对照组为0.25 μL control siRNA的稀释液),25 ℃培养3 h后,每孔加入2 μL质量浓度为10 mg/mL的脂多糖免疫刺激细胞,分别于不同时间点取样,每个时间点3个平行。由于基因沉默预试验显示,在3 h后MyD88的表达量开始降低,5 h达到最低,因此,试验设计在加入siRNA之后的3 h时,再加入脂多糖进行免疫刺激。
2 结果与分析
2.1 褐牙鲆白细胞介素21基因编码序列的分析
转录组测序结果得到褐牙鲆白细胞介素21 cDNA相似序列长度为1606 bp,已经提交到 GenBank,登录号KY783929。开放阅读框分析显示,该序列含有全部蛋白编码序列(cds)。根据转录组测序信息,设计引物通过PCR克隆了编码序列区域的全部序列,进行测序验证。对克隆的编码序列分析表明,读码框从起始密码子(ATG)开启,末尾以终止密码子(TGA)结束,表明该段cDNA序列包含完整的蛋白序列。褐牙鲆白细胞介素21基因开放阅读框区全长429 bp,一共编码142个氨基酸,蛋白质的理论等电点为9.13,分子量16.018 ku。利用SignalP 软件分析,发现白细胞介素21在N端存在19个氨基酸残基的信号肽序列(图1)。

图1 褐牙鲆白细胞介素21基因的编码序列Fig.1 The of coding sequence IL-21 gene in blastard flounder P. olivaceus 起始密码子(ATG)和终止密码子(TGA)用*表示,灰色阴影部分为信号肽. * denotes initiator codon(ATG)and termination codon(TGA), and the signal peptide is shaded in gray.
2.2 褐牙鲆白细胞介素21基因编码蛋白质的同源性及进化分析
褐牙鲆白细胞介素21蛋白序列BLAST比对结果显示,该蛋白质的氨基酸序列与其他物种白细胞介素21氨基酸序列具有很高的相似度(图2)。在美国国立生物技术信息中心上搜索已报道的白细胞介素21家族全长氨基酸序列,分别采用邻接法(图3)、最大似然法(图4)和非加权组平均法(图5)进行分子系统进化树的构建,3种方法得到一致的结果,褐牙鲆蛋白序列同其他物种的白细胞介素21蛋白序列处在同一进化分支上,在亲缘关系上同尖吻鲈(Latescalcarifer)以及深裂眶锯雀鲷(Stegastespartitus)最近,具有共同进化的祖先和相似的生物学功能。
2.3 褐牙鲆白细胞介素21基因的组织表达模式分析
利用实时荧光PCR方法,以β-actin为内参基因,相对定量地检测了白细胞介素21在健康褐牙鲆7个重要组织器官(心脏、肝脏、脾脏、头肾、肌肉、肠道、鳃)中的表达分布(图6)。研究结果表明,白细胞介素21基因广泛表达于所检测的褐牙鲆各组织中,但表达量存在差异,白细胞介素21在肝脏中的表达量最高,小肠次之,因此,白细胞介素21基因在褐牙鲆组织中的表达具有广泛性和特异性。
2.4 迟钝爱德华氏菌脂多糖刺激下褐牙鲆白细胞介素21基因的表达谱
试验用提取的迟钝爱德华氏菌脂多糖对褐牙鲆进行免疫刺激,通过实时荧光PCR方法检测褐牙鲆头肾、脾脏组织中白细胞介素21基因的表达变化(图7)。白细胞介素21基因在脾脏组织中6 h时表达量开始升高,在12 h的时候达到最大值,为对照组25倍(P<0.01);在头肾组织中的表达量自1 h时表达量开始上升,但变化不显著,在6 h时达到最大值,为对照组5倍(P<0.01)。由此可见,脾脏中的应答反应强于头肾。

图2 褐牙鲆白细胞介素21与其他物种白细胞介素21 氨基酸序列比对Fig.2 Comparison of amino acid sequence alignment of IL-21 in blastard flounder P. olivaceus with IL-21 in other species 尖吻鲈,登录号:XM_018703016.1;深裂眶锯雀鲷,登录号:XM_019258432.2;褐牙鲆,登录号:KY783929;尼罗罗非鱼,登录号:XM_025910006.1;伯氏朴丽鱼,登录号:XM_014334141.1;慈鲷鱼,登录号:XM_013909155.1;大黄鱼,登录号:XM_019258432.2. Lates calcarifer, accession number: XM_018703016.1; Stegastes partitus, accession number: XM_019258432.2; Paralichthys olivaceus, accession number: KY783929; Oreochromis niloticus, accession number:XM_025910006.1; Happochromis burtoni, accession number:XM_014334141.1; Pundamilia nyererei, accession number:XM_013909155.1; Larimichthys crocea, accession number:XM_019258432.2.
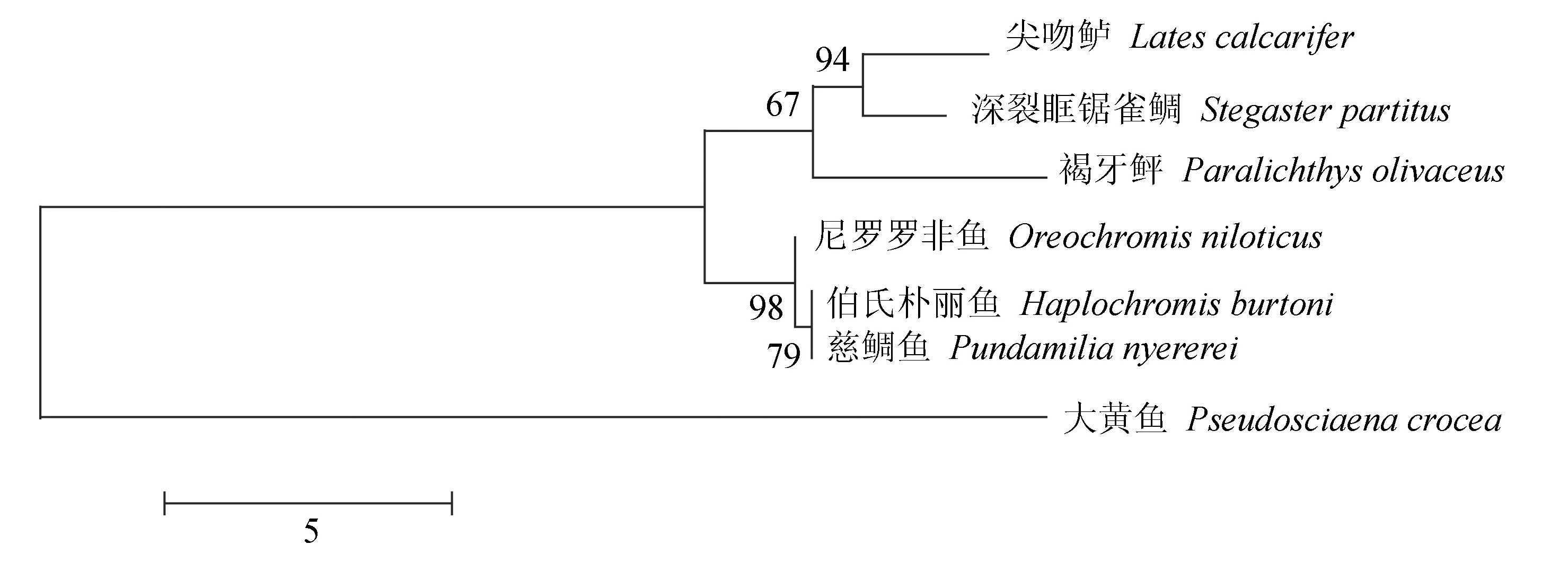
图3 邻接法构建白细胞介素21序列进化系统树Fig.3 The phylogenetic tree of IL-21 by neighbor-joining method 比例尺代表每个残基的替代率,位于节点上的数字表示置信度,重复次数为1000,下同. The scale bar represents replacement rate of each residue. Numbers on nodes represent confidence level of 1000 bootstrap replicates. et sequentia.
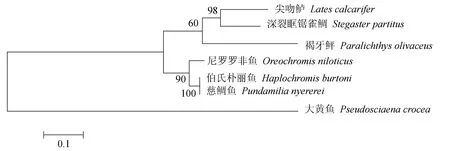
图4 最大似然法构建白细胞介素21序列进化系统树Fig.4 The phylogenetic tree of IL-21 gene sequence by MLE
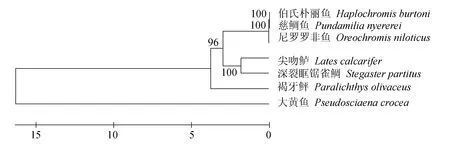
图5 非加权组平均法构建白细胞介素21序列进化系统树Fig.5 The phylogenetic tree of IL-21 gene sequence by UPGMA
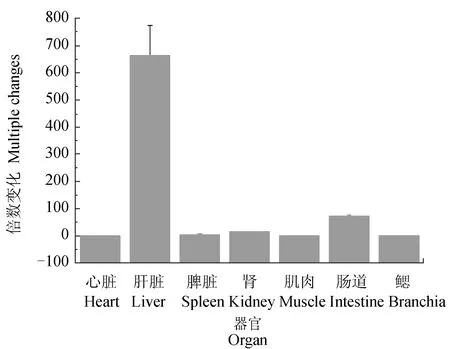
图6 褐牙鲆白细胞介素21基因在健康褐牙鲆中的组织表达Fig.6 The relative expression of IL-21 in different tissues of healthy bastard flounder P. olivaceus

图7 迟钝爱德华氏菌脂多糖刺激褐牙鲆后 白细胞介素21基因的表达Fig.7 The gene expression of IL-21 in blastard flounder P. olivaceus stimulated by E. tarda LPS
2.5 褐牙鲆白细胞介素21与Toll样受体信号通路的关联性
为了检测白细胞介素21与Toll样受体(TLR)信号通路是否存在相关性,试验以原代培养的褐牙鲆头肾淋巴细胞为材料,采用RNA干扰技术将Toll样受体信号接头分子MyD88基因进行沉默,检测白细胞介素21的表达变化[33-34]。Toll样受体是一种膜受体,可识别入侵微生物的保守结构分子,启动机体固有免疫。MyD88是一种接头蛋白,含有TIR结构域,可与膜受体的TIR域作用,向下游传递信号,因此是TLR信号通路中的关键接头分子,具有信息传递的承上启下作用[35-37]。siRNA是一种短片断双链RNA分子,能够与序列同源互补的靶mRNA特异性结合,诱导靶mRNA发生降解,从而使靶mRNA沉默。试验结果显示,褐牙鲆头肾淋巴细胞经MyD88的siRNA基因干扰作用后,MyD88的表达量在2 h开始降低,在5 h时干扰效果最明显,表达量降至对照组的1/3(图8)。同时,白细胞介素21的表达量检测到在第4 h降低,第5 h降至最低(图9),变化趋势与MyD88一致,说明白细胞介素21基因的表达也同样受到了抑制。试验中于第3 h加入提取的迟钝爱德华氏菌脂多糖对基因沉默后的细胞进行免疫刺激,则MyD88和白细胞介素21基因的表达量从第6 h开始逐渐回升,表明这两个基因对迟钝爱德华氏菌脂多糖都产生了应答反应。结果暗示,白细胞介素21与MyD88可能存在一定关联性。
3 讨 论
白细胞介素21作为近期发现的调节因子,受到广泛重视,已有大量关于白细胞介素21的研究。胡春蓉等[38]发现,白细胞介素21会加速慢乙肝患者HBeAb形成,会促进B淋巴细胞的扩增,在慢乙肝临床转归过程中的作用也具有重要临床价值;钮晓音等[39]发现,白细胞介素21通过上调白细胞介素17的表达参与了Graves的发病机制,且这一机制可能并不依赖于甲状腺功能,而与自身免疫过程相关,还发现白细胞介素21在类风湿性关节炎、系统性红斑狼疮、炎症性肠病、系统性硬化病等多种免疫过强型病疫的发病机制中发挥着极为关键的作用;项黎新等[21]利用脂多糖诱导黑青斑河鲀,发现白细胞介素21在头肾和脾脏中表达出现明显的上升,表明白细胞介素21很可能是一个炎症因子,参与感染免疫过程。然而目前,对白细胞介素21的相关研究主要集中在哺乳动物,非哺乳动物中白细胞介素21的研究很少,而关于褐牙鲆白细胞介素21基因的研究至今尚未见报道。
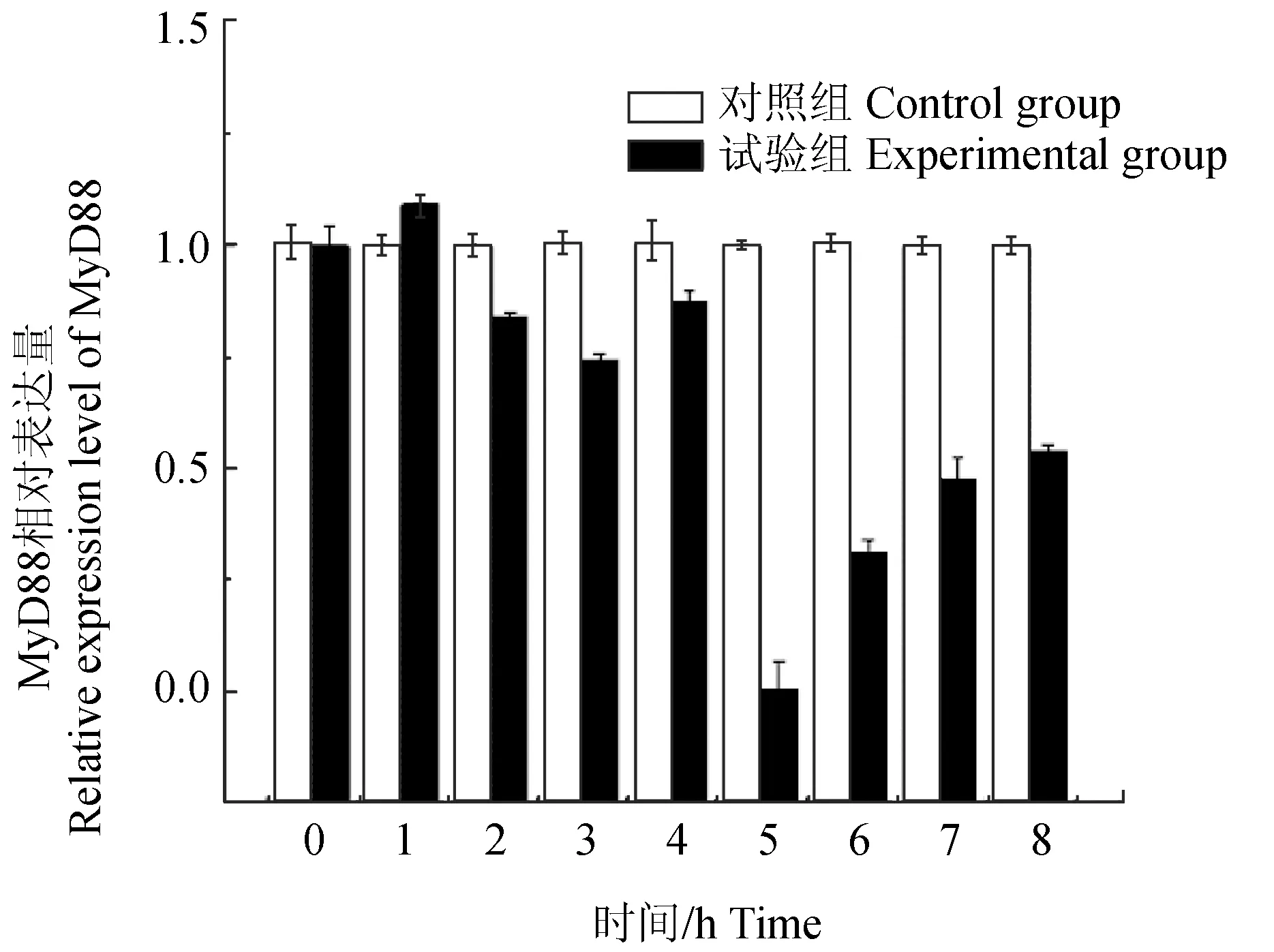
图8 MyD88基因沉默后脂多糖刺激下MyD88基因的表达Fig.8 The expression of MyD88 stimulated by LPS by silencing MyD88
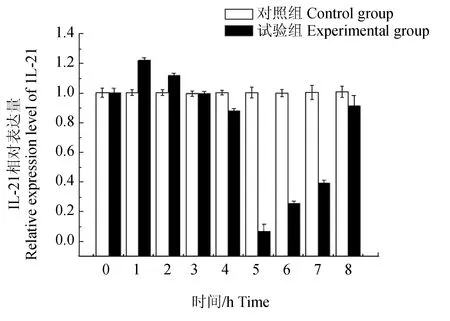
图9 MyD88基因沉默后脂多糖刺激下白细胞介素21基因的表达Fig.9 The expression of IL-21 stimulated by LPS silencing MyD88
3.1 褐牙鲆白细胞介素21基因的组织表达模式分析
有研究表明,在黑青斑河鲀中白细胞介素21仅在肠、鳃和性腺组织中发现有少量表达,作为鱼类主要免疫器官的头肾和脾脏反而未发现有表达;在红鳍东方鲀中,仅健康鱼体的头肾中有少量表达;而在哺乳动物如牛和小鼠,正常组织中不表达[22]。本试验中,健康褐牙鲆组织表达量结果显示,在褐牙鲆中白细胞介素21基因具有组织表达的特异性,在肝脏组织表达量最高,在脾脏、头肾中的表达量并不是很高,这可能是因为头肾和脾脏是鱼类极为重要的免疫器官,白细胞介素21作为一类重要的免疫细胞因子,在重要免疫器官的表达量较其他器官明显偏高,而白细胞介素21在肝脏中表达量最高,这可能是因为肝脏中储存大量的肝糖原,是重要的代谢器官。由此可见,白细胞介素21在不同生物不同组织中的表达有差异。
3.2 褐牙鲆白细胞介素21基因对病原体刺激下的免疫应答反应
使用脂多糖进行刺激,试验组白细胞介素21的表达量出现了明显的上升,说明脂多糖刺激诱导了白细胞介素21的表达,这与转录组测序结果相似,也和实验室前期工作不矛盾。已有研究表明,脂多糖诱导黑青斑河鲀白细胞介素21基因在头肾和脾脏的表达量较高,在鳃、肠道和皮肤中也有少量表达[21],这与本研究结果基本一致。白细胞介素21基因表达量上升,原因可能是白细胞介素21是一个炎症因子,和B细胞的增殖有关,B细胞作为获得性免疫过程中非常关键的一种免疫细胞,参与其中的免疫反应,在机体清除病原体的过程中,B细胞会大量增殖,所以白细胞介素21的表达量升高,由此可见,白细胞介素21参与了褐牙鲆的免疫调节过程,是一类重要的免疫因子,在迟钝爱德华氏菌致病性方面极为关键,这一结果为确定白细胞介素21基因免疫功能研究病原体的选择以及制备褐牙鲆细胞组织的选择提供了理论依据。
3.3 褐牙鲆白细胞介素21与Toll样受体信号通路的关联性
Toll样受体作为重要的病原相关分子模式识别受体家族成员之一,可通过Myd88依赖型信号通路激活机体的免疫应答反应,激活下游的调控因子NF-κB或AP-1,传导免疫信号,促进前炎症因子(如白细胞介素6、白细胞介素8、白细胞介素12和TNF的表达)的释放,进而诱导白介素、干扰素等刺激细胞生成效应分子最终杀死外来病原微生物活化转录因子,进而对疾病进行防卫、抵御和控制,是一类参与非特异性免疫非常重要的蛋白质分子[40-43],白细胞介素位于TLRs下游信号通路中,在传递信息,激活与调节免疫细胞,介导免疫细胞活化、增殖、分化及炎症反应中均起到举足轻重的作用,白细胞介素21信号途径在众多免疫细胞内均有表达,能够增强细胞的特异性反应。项文清[44]研究发现,在各免疫细胞内的作用过程中,肝细胞内通过增强MyD88的表达,从而部分增强其下游通路的激活通过激活MyD88-IRAK通路来增强炎症因子白细胞介素6的表达。本试验中通过siRNA干扰的手段研究褐牙鲆头肾细胞在MyD88基因沉默的条件下白细胞介素21的表达量变化情况,发现MyD88的表达量和对照组相比表达量明显降低,说明得到的 MyD88双链 RNA成功沉默了 MyD88的表达,与此同时,白细胞介素21的变化趋势与MyD88的变化趋势基本一致,MyD88基因被沉默后, 白细胞介素21基因的表达量也发生了明显的降低;在加入脂多糖免疫刺激后,又开始逐渐上升,变化趋势与MyD88一致,与其他学者研究结果相同,表明白细胞介素21同白细胞介素6相似,与MyD88在功能上具有一定关联性,暗示着白细胞介素21基因的表达受到MyD88的调控。这是因为白细胞介素21的表达受到TLRs信号通路的调节,MyD88作为Toll样受体信号通路中的一个接头分子,在传递上游信息和疾病发生发展中具有重要的作用[41-43],所以当MyD88表达被干扰后,Toll样受体的信号通路会发生阻断,下游的免疫基因白细胞介素21的表达量也会随之减弱,当再加入脂多糖进行免疫刺激时,白细胞介素21的表达量与MyD88又开始逐渐上升,与预期一致,验证了笔者的猜想。
4 结 论
本试验研究了白细胞介素21在各个健康组织、器官中的转录水平,深入探索鱼类免疫因子白细胞介素21在应答迟钝爱德华氏菌入侵的免疫应答过程中的分子机理,通过对比可以看出,白细胞介素21可能在抵抗炎症反应中有着极为重要的作用。所得结果将有助于加深人们对鱼类免疫系统调节机制的全面认识,为建立一种有效的、能够预防和控制迟钝爱德华氏菌感染的免疫防治新措施提供理论基础。
