不同补光位置下草莓光合特性、光能和电能利用效率研究
2020-03-16王昊马文礼陈永伟卜建华李宗泽靳韦韦峰
王昊, 马文礼, 陈永伟, 卜建华, 李宗泽, 靳韦, 韦峰,2*
(1.宁夏农垦农林牧技术推广服务中心,银川 750001;2.西北农林科技大学园艺学院,陕西 杨凌 712100)
光环境是设施栽培中最重要的环境因子,对植物的形态建成、光合特性、生理代谢、产量品质均有广泛的调控作用[1]。研究发现,对光质、光照强度和光照时间进行任何调控都会不同程度地影响植株内在复杂的光接收和信号转导系统的运转,且光通过对生物膜系统结构、透性变化或基因表达变化进行调节,也能够促成细胞分化并改变其结构功能,进而影响到植株的生长代谢。日光温室生产过程中寡日照通常为限制作物高产稳产的主要因素之一,弱光环境通常会导致植物叶片净光合速率下降,影响植株光合产物的合成与积累[2]。弱光环境下由于光合作用受限,植株干物质积累受到抑制,进而导致花芽分化延迟,开花节位提升,对作物(如番茄、草莓等)的开花授粉和坐果都造成严重影响,导致产量下降[3-4]。因此,利用补光设备调控植株形态建成和生长发育是温室高效栽培领域的一项重要技术。
随着植物补光灯在设施农业中得到大规模的应用和推广,补光灯类型和补光方式也呈现多种多样的趋势。目前,人工补光方式主要有顶端补光和株间补光。研究发现,温室中植株中下层叶片与上层叶片相比,可接受到的光照强度更低,在光合作用光响应曲线中处于光限制区域,因此,株间补光比顶层补光对光合速率的提升更高效[5]。也有学者在温室黄瓜、甜椒等作物中开展株间补光试验,研究其对果实产量的影响,发现株间补光能够显著增加作物早期产量,并且能增加果实的大小及果实数量;提高第一穗果的数量及干物质含量[6-7]。虽然株间补光较顶端补光有诸多优势,但其单位面积投入成本也高于水平补光,且由于大部分温室补光灯的部署只是依靠经验简单的距离和高度估计或者等间隔的悬挂,未考虑光照空间分布不均匀,导致有光照不足的地方,或者补光灯过密以及悬挂高度过低导致的光照强度过强给植物造成伤害等现象[8]。光能利用效率和电能利用效率与光源类型、作物种类、环境条件和栽培密度等密切相关。生产中改善光能利用效率的方法主要有:①增强光源本身的发光效率,如使用电光转化效率较高的LED灯代替散热量较高的高压钠灯和荧光灯;②提高植株冠层光截面和光捕获量;③提高植物光合速率增加产量的方法,如改善冠层结构、减小光呼吸、优化环境条件(CO2浓度、氮水平等)和应用基因技术。而针对温室环境可控的栽培系统中红蓝光作为人工光源的研究,多集中于不同光强或光质对植物生长和品质的影响,有关不同补光方式下作物光能利用效率和电能利用效率的研究还鲜有报道。本研究以激光植物生长灯为人工光源,基于光强分布的温室补光优化模型,对比分析LED光源顶端补光与水平补光对草莓光合特性、光能和电能利用率的影响,以期为日光温室草莓生产补光优化提供理论基础。
1 材料与方法
1.1 试验材料
1.1.1供试品种及栽培模式 供试品种为‘甜查理’,种苗购自宁夏农垦平吉堡庄园有限公司,采用椰糠槽无土栽培模式,栽培槽由槽体、基质和渗液层3部分组成,槽长800 cm,宽20 cm,槽与槽之间行距80 cm,草莓定植株距20 cm。
1.1.2试验地概况 试验温室位于宁夏农垦平吉堡现代农业园区(N38°,E106°),温室坐北朝南,东西延长。温室跨度8 m,长度80 m,脊高4 m,后墙高度2.4 m,骨架为钢架结构。温室前屋面采用“明静华”EVA-I型长寿无滴膜(宁夏禾佳丰有限公司),夜间前屋面覆保温被保温,采用电动卷帘机。温室内草莓于2017年9月定植。
1.1.3营养液 采用专用全水溶肥配置(苗期N∶P∶K=25∶25∶10,花期N∶P∶K=20∶20∶20,果期N∶P∶K=16∶8∶34),全生育期供应。
1.1.4LED光源 采用西安同凯电子科技有限公司的激光植物生长灯作为草莓专用灯,光质组成红∶蓝=7∶1,红光光谱线638.12 nm,蓝光光谱线467.75 nm,功率为8 W,发光面为20 cm×35 cm的矩形状。
1.2 试验设计与方法
试验设置3个处理,LED顶端补光(C1)、LED水平补光(C2)和不补光(CK)。LED光源部署参考矩阵多光源补光灯光强空间分布模型,处理间采用反光幕进行隔断,各处理均设置3个重复,每个重复小区面积为50 m2。每天早上4:00至揭苫(8:00左右)、晚上放苫(16:00—16:30)至20:00进行补光,每天补光8 h,阴天(温室中光照强度低于草莓光补偿点所需光照强度)全天开启补光灯。在植株整个生育期,保证水平及顶光补光处理下植株顶端接受的光照强度为500 μmol·m-2·s-1。
顶端补光位置及高度确定方法按照几何光学的强度定律简化公式I2/I1=l2计算,I1为两测量点的直线距离。补光灯悬挂高度随植株生长高度调整[6],每小区安置4盏顶端补光灯。水平补光位置以光线直射植株生长点为标准,高度位置随植株生长高度调整,每小区安置2盏水平补光灯。
1.3 测定指标及方法
1.3.1植株调查 从育苗开始,记录各个生育时期,记载田间管理,各处理小区植株总数60%出现某一特征显著变化为某一生育时期记录标准。株高、茎粗等指标的测定采用定期定株观测,即定苗后,在每一小区选取生长均匀一致的植株10株挂牌标记进行定期观测,每隔7 d定株测定株高、茎粗等。
1.3.2光合速率加密观测 采用CIRAS-1光合仪(浙江托普云农科技有限公司)进行叶片光合加密观测,在草莓盛果期(12月25日)典型阴天早上9:00—10:00测定成熟叶片,光照强度为500 μmol·m-2·s-1,叶温18~20 ℃,外界CO2浓度为480 μmol·mol-1。水平方向,每处理测定净光合速率(net photosynthetic rate,Pn)、蒸腾速率(transpiration rate,Tr)、气孔导度(stomatal conductance,Gs)和胞间CO2浓度(intercellular CO2concentration,Ci)等参数。由北向南测定5个植株的叶片,分别以Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ代表,重复3次。
1.3.3光能、电能利用效率和生产单位重量草莓果实所需的光量子数和消耗的电能计算 在果实成熟过程中,分别在花后7 d即果实小绿、花后14 d果实大绿、花后21 d果实白果、花后25 d果实1/2红和花后28 d果实全红时进行取样,并按照下列公式[2,9]进行计算。
式中,p为生产单位质量草莓果实所需要的光量子数,mol·g-1;Fn为第n次采样所收获草莓果实干重的平均值,g·株-1;Ffinal为收获时草莓果实干重的平均值,g·株-1;D为栽培密度,13 株·m-2;PAR为冠层处处理光强,μmol·m-2·s-1。
式中,K为生产单位重量草莓果实所消耗的电能,MJ·g-1;P为光源功率,W;S为栽培面积, m2。
式中,EUEj为第j次与第j-1次采样期间草莓果实的电能利用效率,%;j为1~6的整数;FWj、FWj-1分别代表第j、j-1次所取草莓果实干重的平均值,g·株-1;Wche为每克干重对应的化学能,MJ·g-1;t为相邻2次采样期间累计总光照时间,s。
式中,LEUj为第j次与第j-1次采样期间草莓果实的光能利用效率,%;Wr为单位面积植物冠层所接收到的光合有效辐射能,W·m-2。
1.3.4叶绿素荧光观测 在草莓盛果期(12月25日)典型阴天早上9:00—10:00、下午14:00—15:00进行叶绿素荧光参数测定,每处理各取植株10株,测定采用FS-3000便携脉冲调制式荧光仪(北京恒奥德仪器仪表有限公司)测定荧光参数:初始荧(Fo)、可变荧光(Fv)、稳态荧光(Ft)、最大荧光(Fm)、暗适应30 min下PSⅡ最大光化学效率(Fv/Fm)、非光化学淬灭(NPQ)、PSⅡ实际光化学效率(ΦPSⅡ)等。
1.4 数据处理
采用Excel 2013对数据进行整理,采用 SPSS 20.0分析数据,采用GraphPad Prism 7.00作图。
2 结果与分析
2.1 不同LED补光位置对草莓植株营养生长的影响
由表1可知,不同补光位置下草莓植株营养生长情况差异显著,茎粗和根系干重方面C1、C2之间差异不显著,但均与CK存在显著差异;叶面积各处理均存在显著性差异,以CK最大,C1最小;根系鲜重各处理间以C1最大,CK最小;根系长度C1较CK存在显著差异,C2与CK不存在显著差异。

表1 不同补光位置对草莓生长的影响Table 1 Effect of different light supplement positions on strawberry growth
2.2 阴天不同LED补光位置下草莓叶片光合特性
植株叶片的光合特性可直接体现植株合成营养物质能力的大小,由图1可知,阴天下各处理由北向南(I~V)植株叶片的光合特性差异显著。除胞间CO2浓度外,各处理的净光合速率、气孔导度和蒸腾速率由北向南(I~V)随着光强的增加整体呈上升趋势。同时也可发现,通过人工补光减少了由于日光温室结构导致的南北光照强度分布不均的影响,各处理植株的净光合速率(Pn)南北差值CK(6.78)>C1(3.93)>C2(1.85),蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)的南北差值均为CK最大,C2最小。由此可见,C2可使温室光照更为均匀。
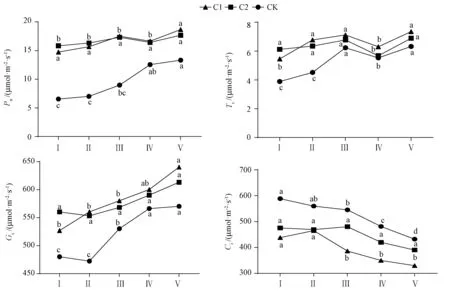
注:同一处理不同小写字母表示差异在P<0.05水平有统计学意义。Note: Different small letters under the same treatment indicate significant difference at P<0.05 level.图1 阴天不同补光位置下叶片光合特性Fig.1 Photosynthetic characteristics of leaves under different light supplement positions in cloudy
2.3 阴天不同LED补光位置下草莓叶片荧光特性
Fv/Fm反映暗适应下PSⅡ反应中心完全开放时的最大光能转化效率,由图2可以得出, 10:00 以C2最高(0.82)、CK最低(0.68),14:00测定值以C2最大(0.55)、CK最低(0.33)。因14:00作物存在光合午休的情况,故Fv/Fm有一个急剧的下降趋势,C1下降最小(0.23),C2下降最大(0.27);Fv/Fo代表PSⅡ潜在光化学活性,C2测定值最大,但下降幅度也为最大,C1相对稳定。ΦPSⅡ为作物叶片实际光化学效率,变化趋势与Fv/Fo相同。10:00各处理NPQ,以CK最高(2.59),C2最小(0.88),说明C2的光能利用效率更高。

注:不同小写字母表示差异在P<0.05水平具有统计学意义。Note: Different small letters indicate significant difference at P<0.05 level.图2 阴天下不同补光位置草莓叶片荧光特性Fig.2 Fluorescence characteristics of leaves under different light supplement positions in cloudy
2.4 不同LED补光位置下草莓果实光能和电能利用效率
图3为不同补光位置下草莓果实光能利用效率和电能利用效率,通过分别对C1、C2和CK下草莓果实不同生育期光能利用效率和电能利用效率进行方差分析,各处理下草莓果实光能利用效率在果实成熟过程中整体为先下降后上升然后再下降的趋势且各处理下在不同生育期的光能利用效率和电能利用效率差异显著,C1和C2的光能利用效率均高于CK,C1和C2的光能利用效率在花后21 d达到最大,分别为0.001 94%和0.001 89%;各处理下生产单位质量草莓果实所需的光量子数(p)分别为CK(4.905)>C2(3.266)>C1(3.171);在电能利用效率方面,C1和C2处理均为先上升后现将的趋势,在花后 14 d 两处理电能利用效率达到最大,分别为C1(0.039%)和C2(0.044%);果实成熟过程中,C1和C2下生产单位质量草莓果实消耗的电能(K)分别为8.207和8.507 kJ·g-1。
2.5 不同LED补光位置下草莓果实产量及品质统计分析
由表2可知,LED灯不同补光位置下草莓果实产量品质相比CK差异显著,可溶性固含物表现为C2>C1>CK,且两两差异显著;可滴定酸含量则表现为C2
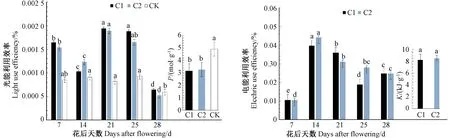
注:同处理下不同小写字母表示差异在P<0.05水平具有统计学意义。Note: Different small letters under the same treatment indicate significant difference at P<0.05 level.图3 不同补光位置下草莓果实光能利用效率和电能利用效率Fig.3 Utilization efficiency of light and electric of strawberry fruits under different light supplement positions

表2 不同补光位置对草莓果实产量和品质的影响Table 2 Effect of different light supplement positions on strawberry fruit yield and quantity
3 讨论
光是高等植物唯一的能量来源,是影响植株叶绿素含量及光合作用的重要因子,植株接受的光照质量不同,将直接影响植株生长发育和形态建成。大量研究发现,植株在果实成熟期时长期处于源库不均衡状态,即光合产物的合成不能满足植株各器官潜在生长需求。有研究表明,减少相应果实数量能够增大剩余果实的大小[10-11]。因此,株间光环境强弱限制了植株群体光合作用与产量的形成,也是植株生殖生长时期处于源库不均衡状态的原因之一。人工补光作为调节植株源库关系的有效手段,能够促进作物的光合作用,提高产量。本研究中,不同补光位置对草莓形态建成影响显著,地下部发育以C1最优,茎粗以C2最优,而CK的叶面积最大,分析原因可能是C1处理植株上部冠层光照面积优于C2、CK,但C2的植株中下部光环境优于其他两个处理。弱光条件下,植株叶面积会出现增大,当光强降低时,比叶面积就会增加,干重、茎粗及根冠比也会受光强降低影响而降低[12-13]。
光合作用是决定作物产量的关键因素之一,植物生命活动的有机物质都来源于光合作用。试验中,通过阴天加密测定温室中各处理由北向南(I~V)植株的光合参数发现,各处理植株叶片光合速率由北向南逐渐增加,这是由日光温室的独特结构特性导致的。但由于C2处理平衡了植株冠层垂直方向上光量,解决了植株叶片光照辐射量随冠层深度增加而迅速降低的问题,增加了单株作物光合效率,缩短了南北植株光合作用差异。C1处理虽也表现出相同作用,但效果较C2差。这说明,株间补光并未影响到植物叶片的表观量子效率和暗呼吸速率,只对最大净光合速率有显著影响[14-15]。随着植株底部冠层光照强度的增加,叶面积指数与比叶面积均出现先减少后增加的趋势。这两者的降低,说明单片叶对光能利用率上升,而非通过增加叶片面积提升对光能的截获能力[16]。试验中也发现,各处理Pn由北向南逐渐增加,Ci的值呈降低趋势,说明试验中各处理南北Pn的差异是由气孔因素导致,而光强的增加,减少了气孔的阻力。
叶绿素荧光参数用来描述植物光合作用机理和光合生理状况,反映植物“内在性”特点,被视为研究植物光合作用与环境关系的内在探针[7,17-18]。光系统Ⅱ的最大荧光效率(Fv/Fm)及其潜在活性(Fv/Fo)是衡量植物光能利用能力大小的指标,Fv/Fm和Fv/Fo值越大,表明该植物的光能利用潜力越大[19]。试验中,通过阴天下10:00和14:00两个时间各处理叶片的叶绿素荧光特性发现,C2处理下植株的Fv/Fm、Fv/Fo和ΦPSⅡ整体相对C1、CK最大,但随着光照辐射的增加,C2处理的下降幅度也最大,说明草莓PSⅡ光反应中心的内禀光能转化效率和潜在活性均随着光强的增加而提高,但在14:00时受到气孔因素影响整体下降,大部分光能以热能的形式转化,故NPQ值随之升高,Fo上升可以缓解植物体内活性氧增加和D1蛋白降解,从而可以避免PSⅡ活性中心发生不可逆转的破坏,对植物造成更大的伤害[4-5]。
光能利用效率和电能利用效率与光源类型、作物种类、环境条件和栽培密度等密切相关。同时也是衡量植物对人工光源所消耗电能和接收光能利用程度的重要指标[20]。试验中,各处理草莓果实的光能利用效率为先下降后上升然后下降,在花后21 d达到最大;而电能利用效率的变化为先升高后下降,在花后14 d达到峰值。两者变化规律没有相关性,这与前人研究结果不同,生菜幼苗定植后,电能利用效率和光能利用效率随定植时间的变化规律基本一致,均出现先上升后下降再上升的趋势,且光能利用效率数值约为电能利用效率的4~5倍。在生产单位质量草莓果实所需光量子数(p)和所消耗电能(K)方面,C1和C2差异不显著。可见光照强度并不是影响这两个指标的主要因子,可能与光质相关,具体可能性还有待进一步通过代谢组学和荧光定量PCR等手段进行验证。同时观测到C1、C2的果实所需光量子数(p)和所消耗电能(K)差异不显著的情况下,两者草莓果实的产量和品质差异不显著,但显著高于CK。说明补光对温室草莓的产量和品质提高有积极促进作用,但是,对于补光长期的生产效益和补光后提高品质的机理还有待进一步研究。
