围垦植稻对崇明东滩湿地产甲烷微生物的影响
2020-03-14张鑫磊宋怡轩汪方圆张耀鸿贾仲君
张鑫磊,宋怡轩,张 洁,汪方圆,张耀鸿,贾仲君
由温室气体甲烷(CH4)增加导致的全球气候变暖是当前威胁人类生存和发展的环境问题之一。根据IPCC的评估报告,大气CH4浓度已达到近百年来的最高水平[1],对全球气候变暖的增温贡献已达15%[2-3]。滨海湿地是介于海洋与陆地之间的一种特殊生态系统[4]。研究表明,滨海湿地CH4排放量占到全球海洋CH4排放总量的20%~39%[5]。因此,研究滨海湿地生态系统CH4排放特征及其影响要素不仅可以深入了解湿地碳循环的生物地球化学行为,还可为合理预测全球气候变化提供重要科学依据[6]。
崇明岛东滩是中国长江口规模最大、发育最完善的河口型滩涂湿地,强烈受到人类活动的影响[7]。随着区域经济的发展,人类对土地的需求越来越迫切,滩涂围垦改农田成为解决土地问题的有效途径[8]。围垦后的湿地不再受潮汐影响,与近海间的物质能量交换基本消失。随后的农作物种植、翻耕、施肥等农业管理措施,使围垦区生境发生显著变化[6],形成了独特的湿地类型——围垦区稻田湿地,与海堤以外的自然滩涂湿地形成了明显的差别[9]。这种土地利用变化改变了土壤生物地球化学过程[10-11],对土壤CH4产生过程会造成深刻的影响。已有研究表明,围垦稻田的甲烷排放通量与滨海滩涂湿地[12]、内陆淡水稻田[13]存在着明显的差异。对其与环境要素之间的相互关系已进行了一些初步研究[12],但是从功能微生物角度,尤其是结合分子生物学方法探索该区域中甲烷产生过程的微生物机理的研究报道较少。
尽管自然界中产甲烷菌分布十分广泛,但是仅能以乙酸、甲酸、H2/CO2、甲醇、甲硫醇类和甲胺类等C1甲基化合物作为产甲烷的底物[14]。根据其利用底物特点可将产甲烷菌分为H2/CO2营养型、乙酸营养型和甲基类营养型产甲烷菌。受到植被、土壤性质、底物类型和硫酸盐含量等环境因素影响,不同类型滨海湿地之间产甲烷菌种类和产甲烷途径存在着很大的变异性[15-18]。那么,围垦改稻田后土壤中各类营养型产甲烷菌的种类如何变化,其对甲烷产生过程的相对贡献如何?这些问题的探索对于深入了解围垦区稻田甲烷排放通量至关重要。
基于此,本试验选取长江口崇明岛东滩自然滩涂湿地和围垦区稻田为研究对象,以空间代替时间变化的方法,研究不同围垦年限情景下土壤产甲烷速率的空间变异特征,分析功能微生物的群落及数量特征,探索其与产甲烷速率之间的相互关系,从而合理评估围垦这一典型的人类干扰方式对区域甲烷排放通量的影响,以期为合理评价我国东部沿海地区围垦造田的生态效应提供理论指导和参考依据。
1 材料与方法
1.1 土壤样品采集
本研究采样地点为上海市崇明岛东滩自然湿地和围垦区稻田(121°09'~121°54'E,31°27'~31°51'N),该区域属于典型的亚热带季风气候,终年温热,降水充沛,年均温15.3℃,年降水量1 003.7 mm。在东滩湿地保护区中,选取低滩位的裸地光滩湿地(BF)和高滩位生长芦苇植被的芦苇湿地(PA),作为围垦0年的样点。在围垦区内选取种稻27、51、86年的稻田(分别记为W-27、W-51、W-86)作为不同围垦年限的稻田样点。围垦植稻不同年限的试验样点根据Cui等[19]的参考文献选取。为了使采样点能更准确反映不同的土壤发育年限,更具代表性,在每个样区内以S形设置6个采样点,各采样点间距约为10~15 m,用土钻取表层0~20 cm鲜土,并将该6个采样点的土壤均匀混合成1个混合样本,作为一个重复,重复3次。放入冰盒中迅速运回实验室冷冻保存备用。
1.2 土壤理化性质测定
土壤理化性质的分析测定主要参考《土壤农业化学分析方法》[20]。土壤总有机碳采用浓硫酸-重铬酸钾消煮-硫酸亚铁滴定法测定。土壤全氮含量采用半微量凯氏定氮法测定。土壤铵态氮和硝态氮用2 mol·L-1KCl溶液浸提后,采用AA3流动分析仪测定。土壤pH采用水土比为2.5∶1提取后,用数字酸度计测定;土壤硫酸根含量采用离子色谱法测定。
1.3 CH4产生潜力测定
将5 g新鲜土壤样品放入40 mL的厌氧培养瓶中,加入10 mL过0.45μm滤膜的超纯水,用胶塞密封后抽真空-充氩气重复3次,以去除瓶中的氧气,达到严格厌氧状态。室温25℃避光条件下预培养7 d,以尽可能去除瓶中残余O2分子。预培养结束后,再抽真空-充氩气3次,进行正式培养14 d。培养期间每2 d从培养瓶顶部抽取500μL气体测定CH4浓度。每次采气前用力摇晃培养瓶60 s,使瓶内气-液相甲烷浓度平衡;采气后补充等量的氩气以保证培养瓶中气压不变,且在计算CH4浓度变化率时需将采气引起的CH4损耗量考虑在内。采用安捷伦气相色谱仪(7890N型)测定CH4浓度。土壤产甲烷速率通过分析培养期间瓶内CH4浓度的变化情况进行计算,公式如下:

式中:P是产 CH4速率,ng CH4·g-1·d-1;d c/d t为培养期间培养瓶上部气相CH4浓度变化率,μL·L-1·d-1;V为培养瓶上部气体体积,L;W为培养的干土质量,g;MW为甲烷摩尔质量,g·mol-1;MV为标准状态下气体摩尔体积,22.4 L·mol-1;Tst为标准温度,K;T为培养温度,℃。
1.4 土壤DNA提取
取0.5 g土壤样品,用Fast DNA Spin kit for soil试剂盒(MP Biomedicals,USA)提取土壤样品中的总DNA。用分光光度计(NanoDrop-1000 UV-Vis)测定土壤DNA浓度和纯度。土壤DNA保存于-80℃冰箱待用。
1.5 定量PCR扩增
采用C1000TMReal-Time System扩增仪定量扩增产甲烷菌的mcr A基因拷贝数。PCR扩增所用引物为ME1/ME2。反应体系为20μL:包括DNA样品1μL、Taq DNA聚合酶10μL、前后引物各0.5μL、无菌水8 μL。
提取产甲烷菌mcr A基因的重组质粒,先测序验证,再用分光光度计测定质粒浓度,并用无菌水将质粒稀释6~8个梯度,用于制作定量PCR的标准曲线,根据该曲线计算目的基因的拷贝数。
1.6 高通量测序
使用古菌通用引物Arc519/Arc806对古菌的16S rRNA基因的V4区序列进行扩增。扩增体系为:稀释后的DNA 2μL,Premix TaqTM25μL,上游引物1μL,下游引物1μL,用灭菌后的去离子水补至50μL。519F/806R的扩增条件为:预变性95.0℃,15 min;变性95 ℃,20 s;退火55 ℃,30 s;延伸,72 ℃,30 s;40个循环。PCR扩增产物使用1.2%(m/V)的琼脂糖凝胶电泳检测是否扩增出特异条带,使用的Marker为DL2000TM,电泳电压设置为180 V。
用OMEGA D6492试剂盒对PCR产物进行纯化,然后用AxyPrep DNA试剂盒回收凝胶后测定DNA的浓度和纯度。
将切胶回收的产物送到中国科学院南京土壤研究所测试分析中心进行建库,并在Illununa MiSeq测序仪上进行16SrRNA基因的高通量测序,获得下机数据后利用QIIME 1.9.1软件进行处理。
1.7 数据统计分析
用SPSS18.0软件进行统计分析,通过单因素方差分析(One-way ANOVA)和Pearson分析土壤理化性质、微生物丰度的差异性以及相关性检验,显著性水平α=0.05。
2 结果与分析
2.1 不同湿地土壤的理化性质变化特征
崇明岛湿地不同围垦年限土壤理化性质如表1所示。围垦稻田的土壤总有机碳(TOC)在15.29~18.34 g·kg-1,显著高于未围垦的自然滩涂湿地,且随围垦年限增加有逐渐增加的趋势。土壤总氮(TN)含量介于0.84~1.20 g·kg-1,围垦稻田与自然湿地之间没有明显差异。围垦稻田土壤NH+4和NO-3含量分别为14.48~23.37 mg·kg-1和 11.79~18.67 mg·kg-1,比光滩湿地(BF)土壤分别高出1.2~2.7倍和0.8~1.9倍。未围垦滩涂湿地(BF和PA)土壤的pH值介于7.67~7.86,显著高于围垦稻田。围垦稻田的土壤SO2-4浓度范围是196~300 mg·kg-1,显著低于光滩湿地(BF)和芦苇湿地(PA)。围垦稻田的土壤EC值显著低于自然湿地,且随围垦年限增加而显著降低。
2.2 不同湿地土壤的甲烷产生潜力差异
如图1所示,光滩湿地的甲烷产生潜力最低,仅为 3.36 ng CH4·g-1·d-1,芦苇湿地的甲烷产生潜力为4.95 ng CH4·g-1·d-1,比光滩湿地高出32%。围垦稻田的甲烷产生潜力速率范围为8.3~23.2 ng CH4·g-1·d-1,且随着围垦年限增加而显著增加。围垦稻田甲烷产生潜力速率均值是自然滩涂湿地的3.3倍。

表1 不同围垦年限土壤的理化性质Table 1 Physicochemical properties of soils with different reclamation years
2.3 不同湿地土壤产甲烷菌的绝对丰度和相对丰度
光滩湿地中产甲烷菌的mcr A基因拷贝数为2.4×106copies·g-1,比芦苇湿地中低一个数量级(表2)。围垦稻田中mcr A基因拷贝数为1.101×108~1.443×108copies·g-1,随着围垦年限增长而增加。根据高通量测序结果计算出产甲烷菌占整个古菌的百分比表征其相对丰度值。可以看出,光滩湿地的产甲烷菌相对丰度显著低于芦苇湿地和围垦稻田,在围垦稻田中随围垦年限增长而相对丰度增加;其中,围垦51年和86年的稻田中相对丰度比滩涂自然湿地高出一个数量级。H2/CO2营养型产甲烷菌的丰度值随趋内陆方向和围垦年限增长而数量级水平增加,其中围垦86年稻田的相对丰度比光滩湿地增加了近100倍。乙酸营养型产甲烷菌的丰度值在各采样点中处于一个数量级,其中围垦稻田中没有显著差异。甲基营养型产甲烷菌的丰度值在光滩中最高,随着趋内陆方向和围垦年限增长而数量级水平减小,其中围垦86年稻田中的丰度值不足光滩湿地的1%。高通量测序分析土壤古菌16SrRNA多样性发现,H2/CO2营养型产甲烷菌主要包括甲烷杆菌属(Methanobacterium)、甲烷砾菌属(Methanoregula)和Methanocella属。其中,甲烷杆菌属(Methanobacterium)是优势属,比其他两个属的相对丰度高出两个数量级。各样点湿地土壤中均检测到乙酸型产甲烷菌,为甲烷鬃菌属(Methanosaeta),相对丰度变异较小。甲基型产甲烷菌只检测到两个属,是甲烷叶菌属(Methanolobus,Methanimicrococcus)和甲烷八叠球菌属(Methanosarcina),其中甲烷叶菌属Methanolobus是优势属。
2.4 产甲烷速率与土壤性质和产甲烷菌丰度之间的相关性分析

图1 不同采样点土壤甲烷产生速率Figure 1 CH4 production rate
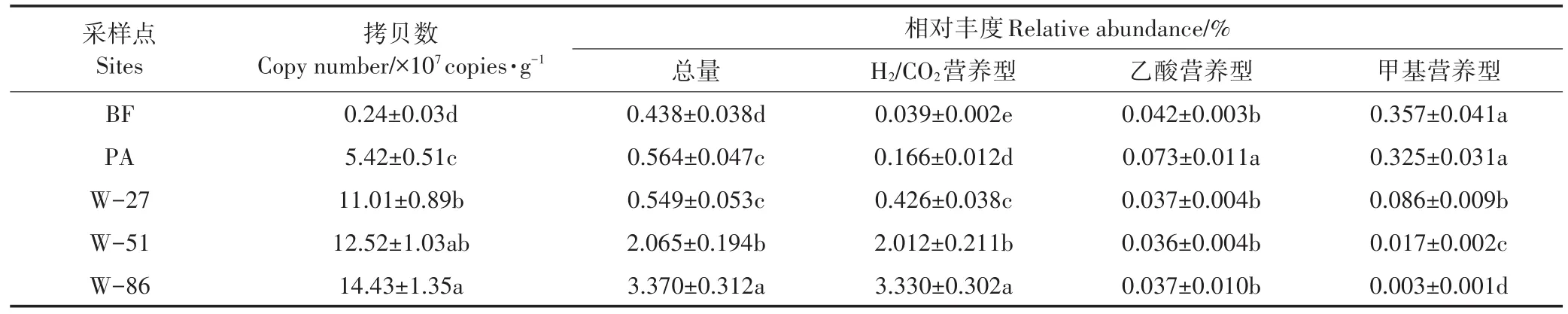
表2 产甲烷菌功能基因mcr A的拷贝数及不同营养型产甲烷菌16SrRNA基因的相对丰度Table 2 Absolute abundance of mcr A gene copies and relative abundance of 16SrRNA gene copies
产甲烷速率与土壤TOC的回归方程达到极显著水平,说明土壤TOC是影响产甲烷速率非常重要的环境要素(表3)。产甲烷速率与土壤TN的回归方程没有达到显著水平,而活性氮(NH+4+NO-3)含量与产甲烷速率的回归方程达到显著水平,说明土壤活性氮在一定程度上影响土壤的甲烷产生过程。相反,产甲烷速率与土壤SO24-含量之间的相关性表现为负对数回归方程,且达到极显著水平。说明随着围垦年限增加土壤SO24-含量降低,是产甲烷速率显著增加的主控因素之一。产甲烷速率与土壤中mcr A基因拷贝数表现为极显著的线性关系,说明产甲烷菌功能基因mcr A数量的增加是产甲烷速率增加的重要因素。
对各营养型产甲烷菌相对丰度与土壤TOC和SO24-含量进行了回归分析(图2)。结果发现,H2/CO2营养型产甲烷菌的丰度值与土壤TOC之间存在极显著正相关关系,而与土壤SO24-含量之间存在极显著负相关关系。乙酸营养型产甲烷菌丰度值与TOC和SO24-含量之间的回归方程均未达到显著水平。甲基营养型产甲烷菌丰度值与土壤TOC之间存在显著负相关关系,而与土壤SO24-含量之间存在显著正相关关系。
将各营养型产甲烷菌丰度值取自然对数后,分析其与产甲烷速率之间的相互关系(图3)。结果发现,产甲烷速率与H2/CO2营养型产甲烷菌丰度值存在显著的正线性关系,与乙酸营养型产甲烷菌丰度值之间的线性方程没有达到显著水平;而与甲基营养型产甲烷菌丰度值之间存在极显著的负相关。

表3 产甲烷速率与土壤理化性质之间的回归方程Table 3 Regression relationship between CH4 production potential(y)and soil characteristics(x)
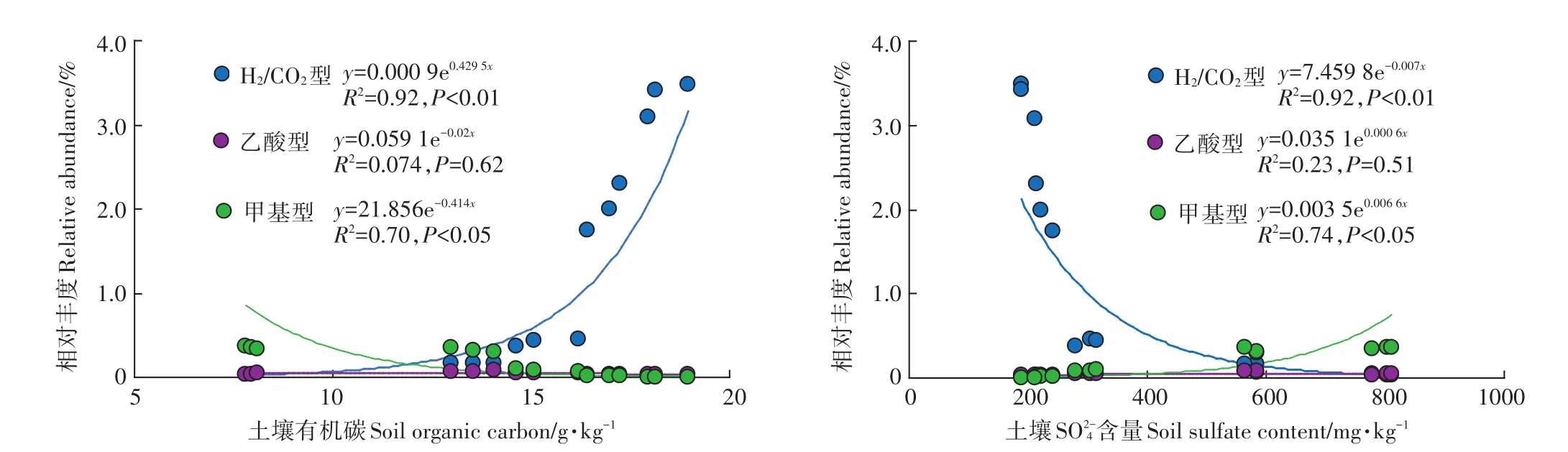
图2 不同营养型产甲烷菌丰度与土壤理化性质的相关性分析Figure 2 Relationship between the relative abundance of three methanogen groups and soil characteristics
3 讨论
人工围垦造田的土地利用方式促使滨海土壤的理化性质发生了显著改变[10-11],这种利用方式会深刻影响该地区湿地土壤的产甲烷过程。本试验结果表明,围垦稻田中产甲烷速率均值达到13.8 ng CH4·g-1·d-1,是自然滩涂湿地的3.3倍,说明围垦造田工程诱发了土壤甲烷的大量排放,这可能与土壤理化性质的变化密切相关。其中,围垦促使土壤的电导率和SO2-4离子浓度显著下降以及围垦后农耕种稻导致土壤TOC含量增加,是导致滨海湿地甲烷产生速率增加的主要原因。一般认为,滨海湿地土壤环境中存在的高浓度SO2-4离子会强烈抑制产甲烷过程[21],其原因是土壤中的硫酸盐还原菌在与产甲烷菌竞争电子供体时具有明显优势[22];且滨海湿地表层土壤受到潮汐冲洗外力作用的影响,土壤有机物及电子供体普遍缺乏,这更加剧了硫酸盐还原菌对产甲烷菌的底物竞争性抑制。另一方面,围垦稻田位于海堤之内,不再受海水潮汐作用的影响,土壤盐分总浓度和SO42-离子显著下降;并且受种稻、施肥、耕作等农业措施的强烈影响,土壤性质发生显著改变,表现为TOC、TN和活性氮显著增加,对产甲烷过程具有重要的促进作用。
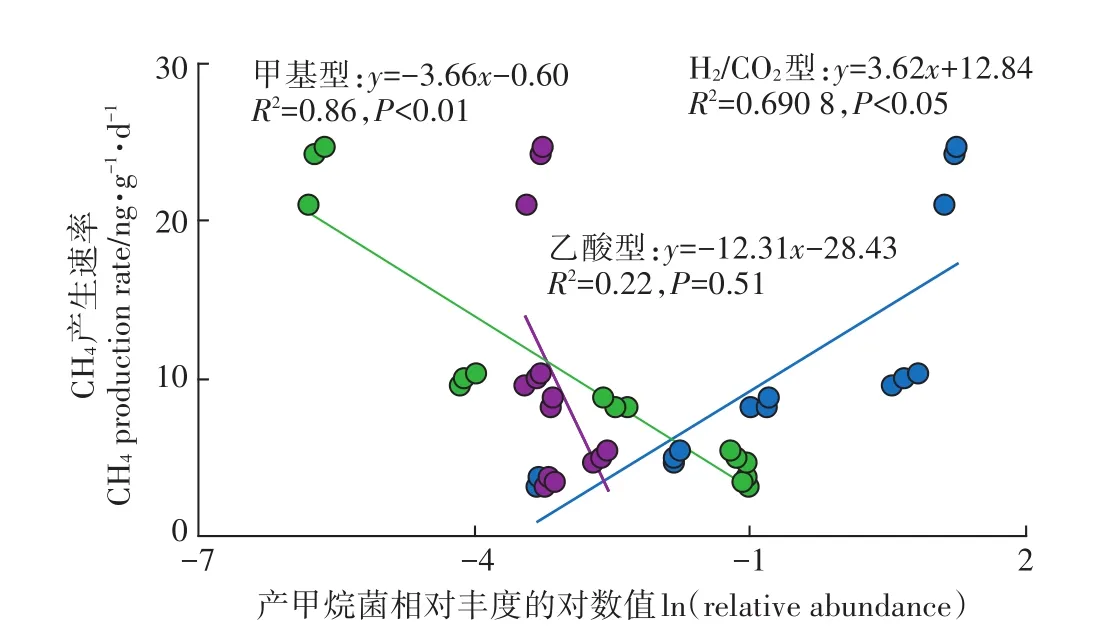
图3 产甲烷速率与各营养型产甲烷菌相对丰度自然对数值的相关性Figure 3 Relationships between the CH4 production rate andln(relative abundance of acetotrophic,hydrogenotrophic and facultative methanogens)
本试验定量PCR结果发现,产甲烷菌功能基因mcr A拷贝数为2.4×106~1.443×108copies·g-1,随着趋陆方向和围垦年限增长而显著增加,说明功能基因mcr A数量增加是产甲烷速率增大的微生物学机理之一。Yuan等[23]报道,在苏北滨海湿地中产甲烷速率与产甲烷菌的16SrRNA基因拷贝数呈显著正相关,与本试验结果一致。高通量测序结果发现,产甲烷菌16SrRNA基因的相对丰度随着趋陆方向和围垦年限增长而数量级水平增加,其与功能基因mcr A的变化趋势完全相同,且与产甲烷速率显著正相关。可见,滩涂湿地围垦改造为稻田后,提高了产甲烷菌的绝对数量和相对丰度,这可能与土壤TOC增加、SO2-4离子下降密切相关。
根据电子底物利用和代谢途径的差异,产甲烷过程分为H2/CO2营养型、乙酸营养型和甲基营养型途径,它们对产甲烷速率的贡献随着环境因素的变化而各异[15-18]。已有研究发现,H2/CO2和乙酸营养型途径是淡水湿地产甲烷过程的两种主要途径,理论上对总甲烷产生量的贡献达到90%以上[24]。本试验发现,乙酸营养型产甲烷菌的相对丰度在不同采样点中处于同一数量级,差异不大;而H2/CO2营养型产甲烷菌相对丰度随着趋陆方向而数量级水平增加,是产甲烷菌总相对丰度值发生分异的主要贡献者。在滨海湿地中H2/CO2和乙酸等底物更多地被硫酸盐还原菌利用,而不是产甲烷菌。所以这两类营养型产甲烷菌被强烈抑制,不是光滩湿地和芦苇湿地的主要产甲烷过程。而在围垦稻田中,耕作层在人工施肥、水稻根系生长等因素影响下,土壤TOC及活性有机碳含量明显增加,为H2/CO2营养型和乙酸营养型产甲烷菌提供了丰富的电子底物,促进了产甲烷过程。在本试验区域内,主要表现为对H2/CO2营养型产甲烷菌的刺激作用。进一步分析发现,H2/CO2营养型产甲烷菌主要包括3个属,其中以甲烷细菌属(Methanobacterium)为主,是H2/CO2营养型产甲烷菌相对丰度发生分异的主要贡献者。各采样点中甲基营养型产甲烷菌的相对丰度以光滩湿地为最高,且占产甲烷菌总相对丰度的81%。说明近岸滨海湿地中甲基营养型产甲烷菌是产甲烷过程的主要贡献者。已有研究发现,甲基营养型产甲烷菌可以利用甲胺类、甲硫醇类和甲醇等C1甲基化合物产生甲烷,而硫酸盐还原菌不能利用这类甲基化合物,对甲基营养型产甲烷菌不会产生底物性抑制作用[14]。因而在SO2-4含量丰富的滨海湿地中,C1甲基化合物歧化作用是产甲烷过程的主要途径[25]。进一步分析发现,本试验中甲基营养型产甲烷菌主要包括2个属,其中以甲烷叶菌属(Methanolobus)为主,是甲基营养型产甲烷菌相对丰度发生分异的主要贡献者。
综上所述,围垦种稻的土地利用方式改变了土壤的理化性质,促进了产甲烷过程。围垦稻田中产甲烷菌的数量大幅度增加,是产甲烷速率增加的主要原因。其中H2/CO2营养型产甲烷菌是产甲烷菌总量发生变异的主要贡献者。
4 结论
(1)滨海滩涂湿地围垦改造为稻田的土地利用方式,显著降低了土壤盐分总浓度和SO2-4离子浓度,促进了耕层土壤的产甲烷速率。
(2)滩涂湿地围垦为稻田后,产甲烷菌的数量大幅度增加,且随着种稻年限的增长而显著增加;其中,H2/CO2营养型产甲烷菌相对丰度大幅增加是产甲烷菌总量发生变异的主要原因。
致谢:在本试验的野外采样过程中,上海崇明东滩湿地自然保护区给予了大力支持和协助,在此表示衷心感谢!
