黄翅绢野螟生物学特性及田间种群动态
2020-03-03张绍华杨陈军孟倩倩孙世伟高圣风刘爱勤
王 政, 张绍华, , 杨陈军, , 孟倩倩,*, 孙世伟, 高圣风, 刘爱勤,*, 杨 婧
(1. 中国热带农业科学院香料饮料研究所, 海南省热带香辛饮料作物遗传改良与品质调控重点实验室, 海南万宁 571533;2. 云南农业大学热带作物学院, 云南普洱 665000)
黄翅绢野螟Diaphaniacaesalis属鳞翅目(Lepidoptera)螟蛾科(Pyralidae)绢野螟属Diaphania,是菠萝蜜、榴莲蜜、面包果等热带特色果树的重要害虫(王平远, 1963; 罗永明和金启安, 1997; Kallekkattiletal., 2019a)。其雌成虫产卵于寄主嫩梢、果柄或花芽表面,幼虫孵出后就近在组织表面取食,约2龄后蛀入嫩枝、花苞及果实取食,老熟幼虫可在蛀道内化蛹,或钻出后吐丝于叶片包裹化蛹(Mridha, 2006; 王政等, 2017; Kallekkattiletal., 2019a)。蛀道内、外均有粪便堆积,易引起果蝇幼虫进入取食,使果实受害部位变褐腐烂,导致果实发育不良、脱落,严重时造成50%以上的减产(刘爱勤等, 2012; 孟倩倩等, 2017)。菠萝蜜树各部位均有利用价值,它的叶和果皮可作饲料,果肉可作水果和蔬菜,种子富含淀粉可作主粮,木材可供制作家具(Khan and Islam, 2004)。近10年来,随着南方果树种植结构调整,菠萝蜜作为木本粮食作物凭借营养丰富、经济价值高的优势,其种植面积每年增长10%以上,我国海南、广东、广西、福建、台湾、云南西双版纳和四川南部等地区均有种植,海南和广东种植面积最大,至2015年底我国约1.67万公顷(谭乐和等, 2017)。黄翅绢野螟在上述地区有不同程度的发生,然而该虫始发期不易发现,暴发成灾后难以有效防控,因此成为制约菠萝蜜产业发展的关键因素之一。
目前关于黄翅绢野螟的研究主要包括其发生为害特点、分布情况及天敌种类等(Soumyaetal., 2015; Rajkumaretal., 2018; Kallekkattiletal., 2019a)。国外已有该虫生物学方面的报道,但不同国家或地区黄翅绢野螟种群各虫态的发育历期、个体大小等生物学特性均存在较大差异,如印度黄翅绢野螟种群与孟加拉国相比,印度种群幼虫发育历期长5 d左右,成虫体长、翅展分别短约2 mm和6 mm,成虫寿命长7~10 d,单雌产卵量多100余粒(Mridha, 2006; Kallekkattiletal., 2019b)。国内研究中,王平远(1963)将黄翅绢野螟最终归类到绢野螟属Diaphania并沿用至今;罗永明和金启安(1997)记述了黄翅绢野螟在我国热带果树上的发生情况;刘爱勤等(2012)报道了该虫的识别及防治技术;王政等(2017)明确了黄翅绢野螟蛹和成虫的雌雄鉴别特征;孟倩倩等(2017)观察了该虫触角感器类型与分布。但该虫在我国的生物学特性和田间种群动态尚不清晰。本研究在实验室条件下,以菠萝蜜叶片作为寄主,系统研究了黄翅绢野螟各龄期形态特征、发育历期及繁殖能力,并在田间调查了该虫的寄主多样性和年种群发生动态,以期为该虫预测预报和科学防控提供理论依据。
1 材料与方法
1.1 供试昆虫饲养及年生活史观察
1.1.1试虫饲养:黄翅绢野螟采自海南省琼中县菠萝蜜种植园(19°3′24″N, 109°50′58″E),室内饲养2代后作为供试昆虫。昆虫食料为菠萝蜜叶片,采自中国热带农业科学院香料饮料研究所试验基地(18°44′29″N, 110°12′11″E),未施农药。将采集的幼虫置于保鲜盒中以新鲜菠萝蜜叶片饲喂,待羽化为成虫后转入网笼,喂食10%的蜂蜜水。饲养条件:温度26±1℃,相对湿度70%±5%,光周期14L∶10D。
1.1.2年生活史观察:室内饲养的幼虫孵化后,批量接种至田间网室中事先种好的菠萝蜜苗上,当第二代幼虫孵化后将其接种至新的菠萝蜜苗上,进行继代饲养,在自然条件下饲养一年,借助放大镜等工具,逐旬观察记录各虫态出现的时间和时长。
1.2 黄翅绢野螟各龄期形态特征及发育历期观察
1.2.1形态特征:从室内种群中挑取卵、幼虫、蛹、成虫等不同虫态,在Olympus SZX16体视显微镜(Olympus, Japan)下观察其形态特征,并测量体长、体宽及蛹重,每个虫态20个重复。
1.2.2发育历期:(1)卵期:按王政等(2017)方法将新羽化的成虫区分雌雄,配对饲养于三角瓶中并用塑料保鲜膜封口(膜上扎孔透气),瓶底放入含10%蜂蜜水的棉球作为成虫的补充营养;每24 h统计一次产卵量,并更换新的三角瓶,每12 h记录一次孵化数量;每2对成虫为1次重复,共20次重复。(2)幼虫期:新孵化幼虫置于保鲜盒中饲养(方法同1.1节,下同),5头/盒,每12 h记录一次幼虫存活与蜕皮情况,并清理;每24 h利用网格纸记录一次取食面积并更换叶片,每5头虫为1个重复,共10个重复。(3)蛹期:按王政等(2017)方法区分雌雄蛹,分开放入保鲜盒中,10头/盒,每12 h记录一次羽化数量,每10对蛹为一个重复,共5个重复。(4)成虫期:将同一天新羽化的成虫区分雌雄,分开置于纱笼中饲养,10头/笼,每日更换棉球至成虫全部死亡,记录成虫寿命,每10对成虫为一个重复,共5个重复。
1.3 黄翅绢野螟幼虫寄主植物调查及取食选择性试验
1.3.1田间调查:2018年7月-2019年6月于中国热带农业科学院香料饮料研究所基地开展寄主多样性调查,包括菠萝蜜、榴莲蜜、面包果、莲雾Syzygiumsamarangense、荔枝Litchichinensis、龙眼Dimocarpuslongan、可可Theobromacacao、芒果Mangiferaindica、黄皮Clausenalansium和柠檬Citruslimon等10种作物。在作物营养生长期、开花期和结果期各调查3次,避开农药药效期,每种作物调查面积约2亩(0.13 hm2),管理水平较一致,每次随机调查10株,每株树分东、南、西、北、中5点取样,统计幼虫量。
1.3.2室内非选择性试验:将1.3.1节描述的10种作物新鲜嫩叶各2片分别放入保鲜盒中,取黄翅绢野螟10日龄幼虫饥饿处理4 h后接至保鲜盒,每盒10头,3次重复,利用网格纸统计6 h后各作物叶片的被取食面积。
1.3.3室内选择性试验:将菠萝蜜、榴莲蜜、面包果、莲雾、可可和芒果等6种作物新鲜嫩叶各2片放入同一保鲜盒,取黄翅绢野螟10日龄幼虫饥饿处理4 h后接至保鲜盒,每盒10头,3次重复,利用网格纸统计6 h后各作物叶片的被取食面积。
1.4 黄翅绢野螟田间种群动态
2018年1-12月,在海南省琼中县菠萝蜜种植园系统调查黄翅绢野螟种群动态,该园为坡地,150余亩(10余公顷),树龄2-3年,植被覆盖率高,节肢动物种群丰富。调查地块约5亩(0.33 hm2),全年不施任何农药,每次随机调查20株树,采用五点取样法统计株虫量(幼虫),折算为百株虫量,约15 d调查一次。
1.5 数据分析
采用SPSS 20.0软件对数据进行单因素方差分析,以Duncan氏新复极差法进行差异显著性检验(P<0.05)。雌、雄蛹和成虫的蛹重、体长、体宽及发育历期等参数均采用独立样本t检验(P<0.05)进行差异显著性分析。
2 结果
2.1 黄翅绢野螟的形态特征
卵:白色至淡黄色,椭圆形,扁平,表面具网状纹(图1: A, B),长0.89±0.06 mm,宽0.71±0.05 mm(表1)。雌成虫单个散产或不规则成堆产卵,数量1~10粒不等,初产卵略透明,后逐渐变白,孵化前成淡黄色。
幼虫:共6个龄期,头部坚硬,初孵和刚蜕皮幼虫头部白色,后渐变为黄褐色或黑色。唇基三角形,额狭长,蜕裂线明显,呈倒“Y”字形,腹节背面有两排对称大黑斑,节间处有两排较小黑斑,胸腹两侧各有一排黑斑,黑斑具毛。1龄幼虫体淡黄色,头黑色,前胸盾黄褐色,臀板淡黄色(图1: E),体长1.92±0.44 mm,头宽0.23±0.02 mm;2-3龄幼虫体、头和臀板黄褐色,前胸盾黑色(图1: F, G),2龄幼虫体长3.56±1.00 mm,头宽0.43±0.03 mm,3龄幼虫体长6.50±0.98 mm,头宽0.64±0.03 mm;4-5龄幼虫体和头黄褐色,前胸盾和臀板黑色(图1: H, I),4龄幼虫体长9.33±0.93 mm,头宽0.95±0.04 mm,5龄幼虫体长12.50±1.02 mm,头宽1.38±0.03 mm;6龄幼虫体黄褐色,化蛹前变为白色,头和前胸盾黄褐色,臀板黑色(图1: J),体长18.30±3.06 mm,头宽1.84±0.03 mm,各龄期幼虫的体长和头宽均存在显著差异(P<0.05)(表1)。

图1 黄翅绢野螟各虫态
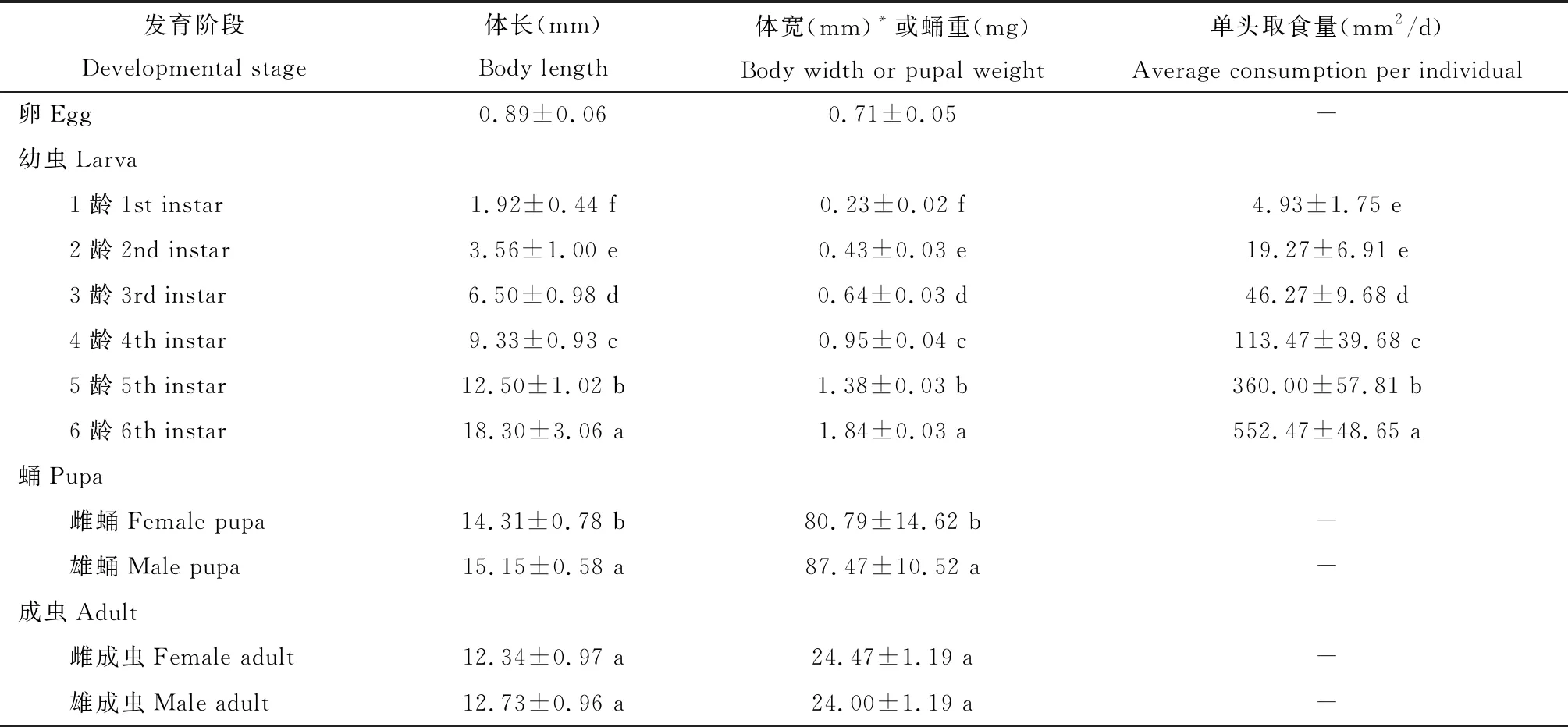
表1 黄翅绢野螟各虫态的基本生物学参数
*幼虫为头宽,成虫为翅展Head width for larvae and wingspan for adults. 表中数据为平均值±标准差,同一虫态同一列数据后不同字母表示差异显著(幼虫期为Duncan氏新复极差法检验,P<0.05; 蛹和成虫期为独立样本t检验,P<0.05)。Data in the table are mean±SD. Different letters following the data of the same developmental stage in the same column indicate significant difference atP<0.05 level by Duncan’s new multiple range test for larval stages and independent samplest-test for pupal and adult stages, respectively.
蛹:纺锤形,幼虫化蛹前以丝结茧,形成不食不动的预蛹。化蛹初期为红褐色,后变为黑褐色,表面光滑,翅芽和足分别长至第5 腹节的前缘和后缘,腹部末端生有8 根毛钩(图1: C, D)。雌蛹第8腹节腹面中央有纵裂缝,其两侧无突起(图1: C),蛹长14.31±0.78 mm,蛹重80.79±14.62 mg;雄蛹第9腹节腹面中央有纵裂缝,两侧各有1个半圆形的瘤状突起,此裂缝与肛门间距较雌蛹短(图1: D),蛹长15.15±0.58 mm,显著长于雌蛹(P<0.05),蛹重87.47±10.52 mg,显著重于雌蛹(P<0.05)(表1)。
成虫:触角丝状,复眼外突呈红褐色,两复眼间有褐色斑点,体麦黄色,前胸背板中央有深棕色鳞片,腹部末端被有鬃毛。前翅三角形,前缘有2个瓜子状空心黄斑,斑周围有黑色曲纹,近顶角处有一塔状黄斑。雌蛾腹部较肥大,末端钝圆,外生殖器被有整齐的短黄色鬃毛(图1: K),体长12.34±0.97 mm,翅展24.47±1.19 mm;雄蛾腹部瘦小,末端狭长,外生殖器被有整齐的长黑色鬃毛(图1: L),体长12.73±0.96 mm,翅展24.00±1.19 mm,雌雄个体差异不显著(体长:P=0.073; 翅展:P=0.082)(表1)。
2.2 黄翅绢野螟的发育历期
黄翅绢野螟个体发育经历卵、幼虫(1-6龄)、预蛹(末龄幼虫吐丝卷叶后不食不动的时期)、蛹和成虫等阶段。在室内饲养条件下,各阶段发育历期分别为:卵期为4.58±0.50 d,幼虫期为21.00±1.36 d,预蛹期为2.50±0.51 d,蛹期为10.20±0.53 d,成虫期为12.31±3.16 d,平均世代历期为50.50±3.54 d(表2)。雌蛹发育历期(10.25±0.63 d)和雄蛹的(10.15±0.86 d)无显著差异(P=0.556),雌蛾历期(13.00±3.05 d)显著长于雄蛾(11.38±3.54 d)(P=0.040)(表2)。

表2 黄翅绢野螟各虫态的发育历期
同一虫态数值后不同字母表示雌雄间发育历期差异显著(独立样本t检验,P<0.05)。Different letters following the data of the same developmental stage indicate significantly different developmental duration between female and male (independent samplest-test,P<0.05).
2.3 黄翅绢野螟的取食与繁殖能力
黄翅绢野螟低龄幼虫取食能力较弱,从3龄幼虫开始取食量迅速增加,每个龄期的取食量与上一龄期相比均存在显著性差异(P<0.05),至6龄幼虫取食能力最强,其取食量是1龄幼虫的100余倍(表1)。黄翅绢野螟单头雌蛾平均产卵量为147.25±84.24粒,范围介于44~298粒(表3)。少数雌蛾羽化当日即可交配产卵,死亡当日尚能少量产卵,大部分雌蛾羽化后2-3 d开始产卵,死亡前2-3 d停止产卵,产卵前期为2.25±0.67 d,产卵期为8.33±1.42 d,其中产卵盛期为6.75±1.24 d(表3)。

表3 黄翅绢野螟雌蛾的繁殖能力
2.4 黄翅绢野螟年生活史
室内和网室饲养结果表明,黄翅绢野螟一年发生8代,世代重叠,结合田间调查发现,该虫全年均可发生,无明显的越冬现象,最早出现成虫是1月中旬,为上年第8代羽化的成虫。第1代为1月中旬至3月下旬,第2代为2月下旬至5月上旬,第3代为4月上旬至6月中旬,第4代为5月中旬至7月下旬,第5代为7月上旬至9月中旬,第6代为8月下旬至11月上旬,第7代为10月中旬至12月下旬,第8代为12月上旬至次年2月中旬(表4)。

表4 黄翅绢野螟在海南兴隆地区的年生活史
F: 上旬The first ten-day period of a month; M: 中旬The middle ten-day period of a month; L: 下旬The last ten-day period of a month. ●: 卵Egg; -: 幼虫Larva; △: 蛹Pupa; +: 成虫Adult.
2.5 黄翅绢野螟的幼虫寄主植物种类及取食偏好性
2018年1-12月田间调查黄翅绢野螟幼虫对菠萝蜜等10种作物的为害情况发现,该虫仅在菠萝蜜、榴莲蜜和面包果等3种菠萝蜜属作物上取食,在莲雾等其他7种作物未见发生(P>0.05)(表5)。其中,在作物营养生长期和开花期,该虫显著偏好取食菠萝蜜和榴莲蜜;在结果期,该虫在3种菠萝蜜属作物上的发生量无显著差异(P>0.05)(表5)。室内非选择性试验结果表明,黄翅绢野螟幼虫在菠萝蜜叶片上的取食面积最大,其次为面包果和榴莲蜜上的,均显著大于莲雾等其他7种作物上的取食面积(P<0.05)(图2: A)。选择性试验结果与非选择试验相似,黄翅绢野螟偏好选择取食菠萝蜜和榴莲蜜,其次为面包果,在这些作物上的取食面积均显著大于莲雾等其他3种作物上的取食面积(P<0.05)(图2: B)。

表5 黄翅绢野螟幼虫在10种作物不同生育期的为害调查
表中数据为平均值±标准差,同一列数据后不同字母表示经Duncan氏新复极差法检验在P<0.05水平差异显著。Data in the table are mean±SD. Different letters following the data in the same column indicate significant difference atP<0.05 level by Duncan’s new multiple range test.
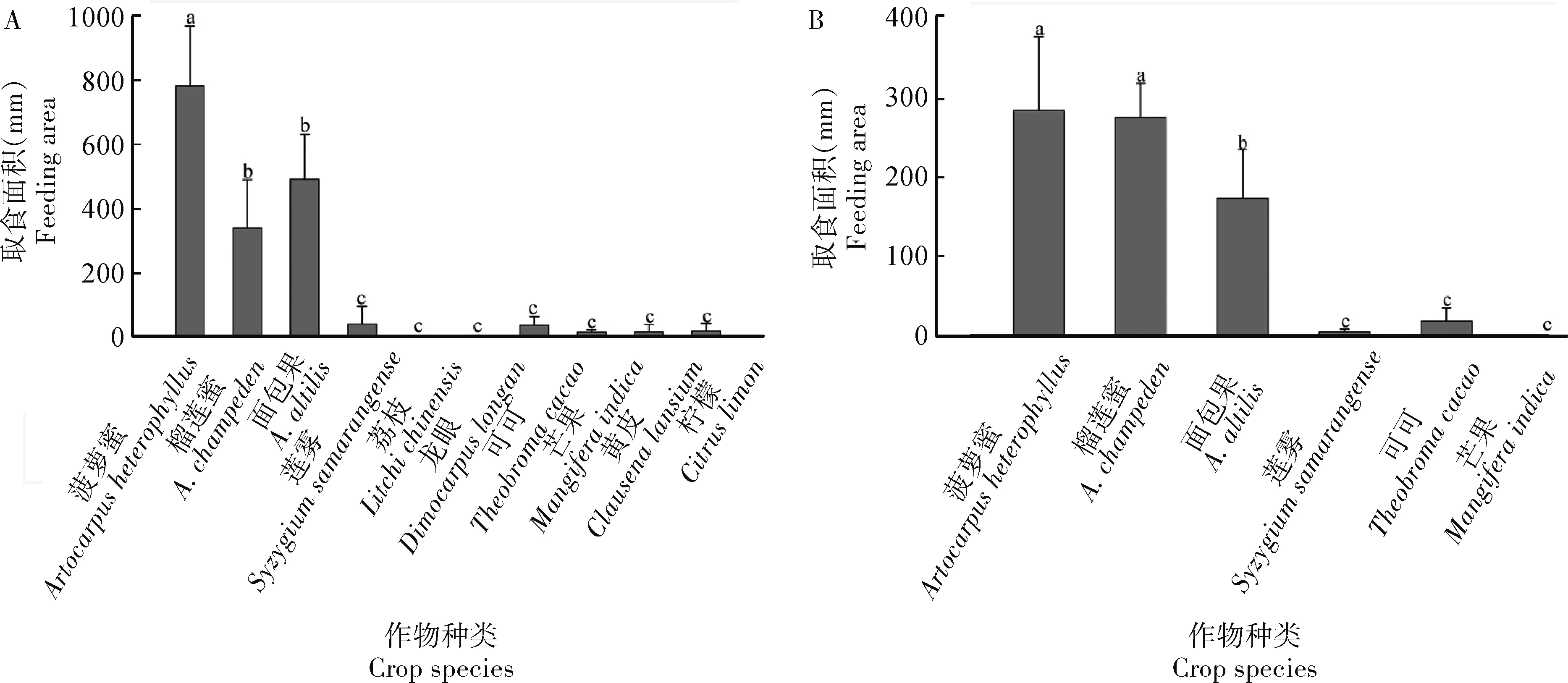
图2 黄翅绢野螟幼虫对不同作物的取食偏好性
2.6 黄翅绢野螟田间种群动态
2018年1-12月在海南琼中县系统调查了黄翅绢野螟田间种群发生动态,结果表明从3月上旬开始黄翅绢野螟种群数量明显增加,至7月上旬始终维持在一个相对稳定的水平;7月中下旬至9月下旬为黄翅绢野螟的种群高峰期,其中8月中下旬种群数量略有下降,但9月中上旬迅速回升至最高峰;10月份后种群开始回落,至12月中下旬种群数量减少到最低水平,并保持到次年2月下旬(图3)。
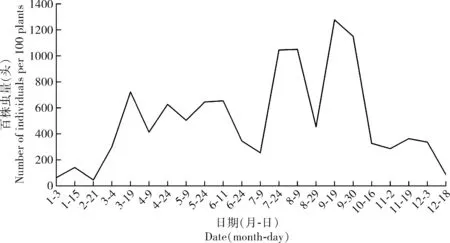
图3 2018年海南省琼中县黄翅绢野螟田间种群动态
3 讨论
昆虫生长发育过程受到诸多因素的影响,它们的个体形态和发育历期等生物学参数通常存在较大的地理差异(涂小云等, 2011; Wangetal., 2013; 智伏英等, 2018),这种差异是由昆虫在不同环境下为达到最大适应值所产生(邹超等, 2017)。许多研究表明,不同地理种群昆虫的生物学差异与纬度相关。例如,朱文超等(2015)发现灰飞虱Laodelphaxstriatellus高纬度种群的若虫历期和单雌产卵量显著长(多)于低纬度种群,成虫寿命则相反,而雌雄比、雄虫体长和雌虫长翅率等参数在不同地理种群间尽管差异显著,但未表现出规律性;邹超等(2017)认为大猿叶虫Colaphellusbowringi的幼虫历期随纬度升高逐渐延长,而生长速率和体重随纬度升高逐渐减小,显示了反贝格曼法则(Chown and Gaston, 2010)。本研究表明,我国海南(中纬度)黄翅绢野螟种群幼虫历期和世代历期分别为21 d和50 d,明显长于孟加拉国(高纬度,24°5′N)种群(分别为12 d和23 d)和印度(低纬度,12°8′N)种群(分别为16 d和31 d)(Mridha, 2006; Kallekkattiletal., 2019b),但不同地理种群间未表现出与纬度相关的规律性变化。另外,上述3个地理种群的成虫个体(体长和翅展)由大到小分别为:孟加拉国>海南>印度,无论雌雄虫均表现出纬度越高个体越大的梯度变化,整体符合贝格曼法则(Chown and Gaston, 2010)。可见,昆虫适应环境变化产生的地理变异是个复杂的过程,不同地理种群各生物学参数差异较大,且未呈现显著的规律性变化,要明确特定地理种群的生物学特性,除参考已报道的其他地理种群外,有必要对该地理种群进行系统性地观察与研究。
昆虫的食性具有一定的稳定性,通常可将食性分为多食性、寡食性和单食性3种类型(彩万志等, 2001)。例如,棉蚜Aphisgossypii能取食不同科的多种植物,是典型的多食性害虫,其寄主超过70科600种植物(Lombaertetal., 2009);小菜蛾Plutellaxylostella取食十字花科(仅一个科)的175种植物,是寡食性害虫(Sarfrazetal., 2005);家蚕Bombyxmori仅取食桑叶Morusalba一种植物,是单食性害虫(Zhangetal., 2019)。本研究表明,无论田间调查还是室内试验,黄翅绢野螟幼虫偏好取食的3种植物均属桑科菠萝蜜属,这与已报道的该虫在印度仅为害7种菠萝蜜属植物的结果相一致(Kallekkattiletal., 2019b),因此该虫应归属于寡食性害虫。然而昆虫的食性又有一定的可塑性,如棉蚜在广泛寄主中,由于长期对单一寄主的适应,形成了不同的寄主专化型或生态专化型(刘向东等, 2004; Sataretal., 2013);又如许多昆虫(包括单食性昆虫)在缺乏正常食物时,可以被迫改变食性(彩万志等, 2001)。本研究仅调查了黄翅绢野螟在10种植物上的为害情况,但该虫能否取食其他科属的植物或存在转换寄主,还需长期调查确认。
寄主植物的质量影响植食性昆虫的繁殖力,包括营养物质和次生化合物(Awmack and Leather, 2002; 程琪等, 2017)。本研究与Kallekkattil等(2019b)均在黄翅绢野螟成虫期以10%蜂蜜水作为营养补充,二者单雌产卵量结果也基本一致(均为140余粒),然而Mridha(2006)未在成虫期进行营养补充的单雌产卵量仅为30余粒,表明该虫羽化后需补充营养才能达到性成熟(伍和平和李保平, 2007)。另外,部分昆虫幼虫期营养是影响成虫繁殖力的主要因素,如幼虫期营养对苹褐带卷蛾雌虫产卵量的促进作用大于在成虫期补充营养(李广伟等, 2019),因此黄翅绢野螟幼虫期营养对繁殖力有无影响还需进一步验证。
本研究发现黄翅绢野螟幼虫的取食量随龄期增加而显著性增大,从3龄开始进入暴食期,6龄幼虫取食量最大,单雌平均产卵量达147粒,具备了鳞翅目害虫食量大、繁殖力强、世代重叠等生态优势,同时该虫钻蛀为害初期不易发现,为其暴发成灾奠定了有利基础。尽管该虫在海南无越冬,全年均可发生,但其田间种群动态呈现一定的季节性规律,春季至夏秋季节(3-9月)种群数量较多,其中夏秋季节(7-9月)种群密度最大,冬季(10月-翌年2月)种群密度最小,这与螟蛾科昆虫桑卷叶蛾Diaphaniapulverulentalis和瓜绢野螟Diaphaniaindica的季节性发生规律相似(Peter and David, 1991; Samuthiraveluetal., 2010),可为该虫精准施药防控提供参考。然而本研究仅对黄翅绢野螟种群动态观察1年,其季节性规律有待进一步验证。另外,有学者报道了黄翅绢野螟寄生性天敌10余种,其中绢野螟绒茧蜂Apantelesstantoni在田间的自然寄生率超过30%(Kallekkattiletal., 2019a),这为该害虫因钻蛀为害而防控困难的问题提供了解决思路。
