不同土地利用类型下岩溶泉域土壤CO2时空变化特征及来源分析
2020-02-21赵瑞一李建鸿董莉莉刘畅张灵艺
赵瑞一 ,李建鸿,董莉莉,刘畅,张灵艺
1. 自然资源部广西岩溶动力学重点实验室/中国地质科学院岩溶地质研究所,广西 桂林 541004;2. 重庆交通大学建筑与城市规划学院,重庆 400074
世界范围内,土壤储存的有机碳达 1.5×1012t(Amundson,2001),这是大气中碳含量的3倍,是陆地生物中碳含量的 2.5倍(Lal,1999)。同时,土壤通过土壤呼吸向大气中释放CO2,成为陆地碳循环中的重要组成部分。尤其是在全球气候变暖的背景下,土壤 CO2释放量增加可能会对大气 CO2浓度和全球变化产生正反馈作用(Liang et al.,2004)。而在岩溶区,土壤 CO2对生态环境的影响还在于土壤CO2是土下岩溶作用的重要驱动力(曹建华等,2011)。虽然土下岩溶作用并没有直接消耗大气CO2,但这导致土壤中一部分CO2以HCO3-的形式进入水体,并且HCO3-在运移过程中能够被水生植物吸收,转化成稳定的有机碳(Liu et al.,2010;王世杰等,2017),从而对大气 CO2起到了减源作用(Martin et al.,2013;White,2013)。因此,了解土壤CO2浓度的时空变化特征及其影响因素将有助于进一步研究岩溶碳循环影响机制。
由于碳酸盐岩溶蚀消耗土壤CO2,岩溶区土壤CO2浓度在垂向上往往表现出双向梯度,即土壤CO2浓度最大值出现在土壤中部而不是土壤底部(Xu et al.,1996;曹建华等,2011),这也被认为是岩溶碳汇效应的重要依据。然而,除土壤CO2形成的 H2CO3溶蚀碳酸盐岩外,外界输入的 H2SO4和HNO3等强酸可能也参与到岩溶作用中。H2SO4和HNO3溶蚀碳酸盐岩首先产生CO2,部分或全部CO2可能再次参与岩溶作用,并以HCO3-的形式进入水体(Perrin et al.,2008)。从地质时间尺度上讲,无论H2SO4和HNO3溶蚀碳酸盐岩最终产生CO2还是HCO3-,HNO3/H2SO4参与岩溶作用都被认为是大气的碳源(刘丛强等,2008;Torres et al.,2017)。而在短时间尺度(<3 ka)上,若H2SO4和HNO3溶蚀碳酸盐岩产生的 CO2能够再次参与岩溶作用并以HCO3-形式进入地下水,此过程并不会向大气排放CO2,只是碳在岩石圈与水圈之间的转移;若此部分CO2没有再次参与岩溶作用,而是直接返回到土壤中,此时H2SO4和HNO3溶蚀碳酸盐岩将直接成为碳源。之前的研究发现,在H2SO4和HNO3输入较大的泉域,雨季土壤CO2垂向变化转变为单向梯度,并且单位水体通过岩溶作用消耗的CO2量降低。但这是否是H2SO4和HNO3溶蚀碳酸盐岩后向底部土壤中释放了CO2造成的,仍需进一步研究(Zhao et al.,2019)。
综上所述,土壤CO2浓度及其来源的差异将对岩溶碳循环产生重要影响。因此,本研究结合之前的水化学监测结果,通过对比不同土地利用类型下土壤 CO2浓度与 δ13C的时空变化特征,分析土壤CO2浓度的影响因素,并对其来源进行示踪,以期为准确估算岩溶碳通量提供理论依据。
1 研究区概况
本研究选择重庆市南川区的柏树湾(BSW)和后沟(HG)2个小型岩溶泉域为研究对象(图1),并对其上覆土壤CO2进行监测。柏树湾泉域出露地层为下三叠统嘉陵江组灰岩(T1j)、白云质灰岩、角砾状灰岩。由于该泉域近几十年受到政府保护,人类活动对其干预较小。在植被恢复过程中,逐渐演化为以马尾松(Pinus massoniana)林为主的次生林,且林下灌丛生长较茂盛。后沟泉域出露地层为下三叠统嘉陵江组白云质灰岩(T1j),土地利用类型主要为农用地。但在本研究期间,后沟埋设土壤CO2收集装置的地点并没有种植农作物。后沟农田中施加化肥并且土壤中含有大量煤铁残渣。柏树湾和后沟监测点的土壤剖面厚度分别为110 cm和150 cm。
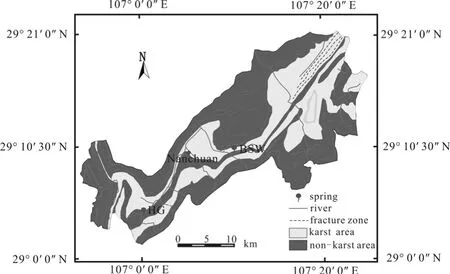
图1 研究区概况示意图Fig. 1 Location of study site
2 野外取样与监测
考虑到土-气碳通量主要来自于深度为 50 cm以内的土壤层(Jassal et al.,2005),柏树湾和后沟均在土下20 cm和50 cm处埋设土壤CO2收集装置。同时根据2个泉域土壤厚度,分别在土壤与基岩交界处(柏树湾110 cm处、后沟150 cm处),以及50 cm处与土-岩界面的中间部分(柏树湾80 cm处、后沟100 cm处)埋设CO2收集装置。将长30 cm、直径为16 mm的PVC管(每隔2 cm钻取1个直径约为3 mm小孔,每根PVC管共钻取3列小孔以避免土壤水储存在 PVC管中而影响实验结果)水平插入土壤中以收集土壤CO2样品。PVC管一端用胶带封住,防止插入过程中土壤堵塞管道,另一端用相同直径的 PVC管接出地面并在端口用试管塞封住。研究期间,每月收集1次土壤CO2样品。在收集土壤CO2样品时,用100 mL的注射器插入密封的试管塞抽取土壤CO2样品,并将其注入1 L的铝箔气体采样袋中以测试土壤CO2浓度和δ13C,每个样品重复测试3次。测试工作在自然资源部岩溶动力学重点实验室完成,仪器为 Picarro G2301分析仪,误差<1%。采集的土壤样品剔除石子及植物残体后风干,研磨后过0.15 mm样品筛。用HCl浸泡以去除土壤中无机碳,再用超纯水润洗3次,烘干后测定土壤有机碳含量及 δ13C。测试工作在中国农业科学院农业环境与可持续发展研究所环境稳定同位素实验室进行,仪器为 Isoprime100-EA。气象数据根据中国气象网(http://www.weather.com.cn/)每天发布的整点天气实况进行统计。日平均温度为每天02:00、08:00、14:00、20:00 4个时刻温度的平均值,降水量为每天整点时刻降水量的总和。利用Origin 8软件进行数据分析。将数据导入Origin 8软件中绘制折线图;通过Origin 8软件绘制散点图,求线性方程和回归系数后进行相关性分析。
3 结果与讨论
3.1 不同土地利用类型下土壤 CO2浓度差异及其影响因素
柏树湾20、50、80、110 cm处土壤CO2浓度平均值分别为 5745、9377.6、11413、9504 μmol·mol-1,后沟20、50、100及150 cm处土壤CO2浓度平均值分别为 3616.3、5120.4、5730.2、4814.4 μmol·mol-1。如上所述,考虑到表层和深部土壤CO2的影响因素不同,本研究分别对2个泉域20 cm处、50 cm处、柏树湾80 cm处与后沟100 cm处、柏树湾110 cm与后沟150 cm处的土壤CO2浓度进行了对比分析。除柏树湾20 cm处在5月小于后沟外,土壤CO2浓度均表现为柏树湾大于后沟(图2)。之前利用泉水离子浓度计算岩溶碳汇量时发现,柏树湾岩溶碳汇量也明显大于后沟(赵瑞一等,2015a;赵瑞一等,2015b)。这与其他研究结果一致(Xu et al.,1996;Zhao et al.,2010),说明柏树湾较强的土壤CO2浓度促进了土下岩溶作用可能是导致其具有较高岩溶碳汇效应的重要原因。
土壤有机质和上覆植被是土壤CO2的两个重要来源,但在不同条件下二者的贡献有所差异(Jan et al.,2018)。虽然后沟土壤有机碳含量明显高于柏树湾(表1),但这主要是煤铁残渣混入造成的。相对于凋落物中新鲜有机碳在 1—2年即可转化成CO2,这些来自煤铁残渣的惰性有机碳矿化速率则较慢,往往需要上百年甚至更长的时间(Paustian,2014)。而已有研究发现,植被对土壤 CO2浓度的贡献有时比气候和土壤性质更加明显(Damien et al.,2014)。柏树湾主要为马尾松林,而后沟埋设土壤CO2收集装置的地块在本次监测期间并没有种植农作物,上覆植被主要为一年生草本植物。马尾松根系相对于草本植物更加发达,其呼吸作用也更强。加之柏树湾大量凋落物为土壤微生物提供了更多的新鲜有机碳源,这可能是导致柏树湾土壤CO2浓度明显高于后沟的主要原因。
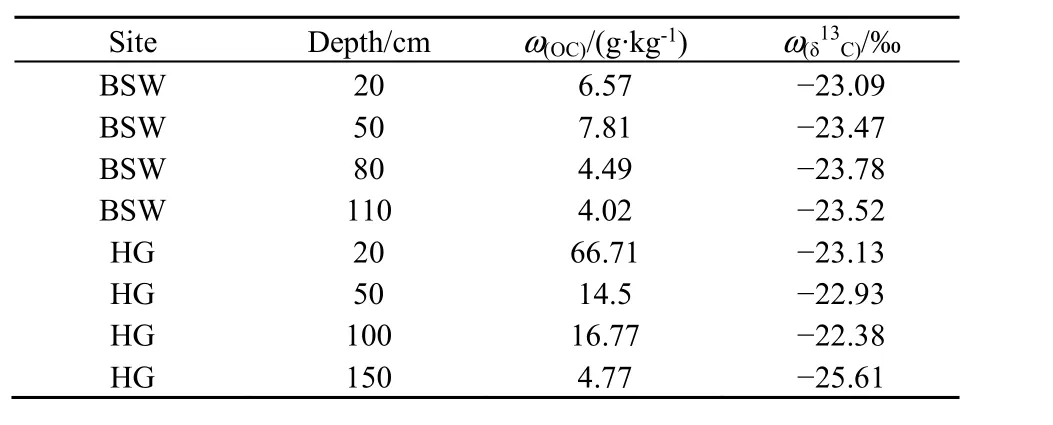
表1 2个泉域上覆土壤有机碳δ13CTable 1 The δ13C of soil organic carbon in spring catchments
另外,柏树湾上覆植被为C3植物,其δ13C在-27‰左右;后沟上覆植被为C4植物,δ13C在-14‰左右(Marion et al.,1991)。通过稳定碳同位素发现,虽然2个泉域土壤有机碳的δ13C比较接近(表1),但土壤CO2的δ13C差异明显。柏树湾土壤CO2的δ13C更偏向于C3植物而后沟则更偏向于C4植物(表2),说明2个泉域土壤CO2主要来源于上覆植被。这也证实了2个泉域土壤CO2浓度的差异可能是受上覆植被的影响。
3.2 土壤CO2浓度的时间变化特征
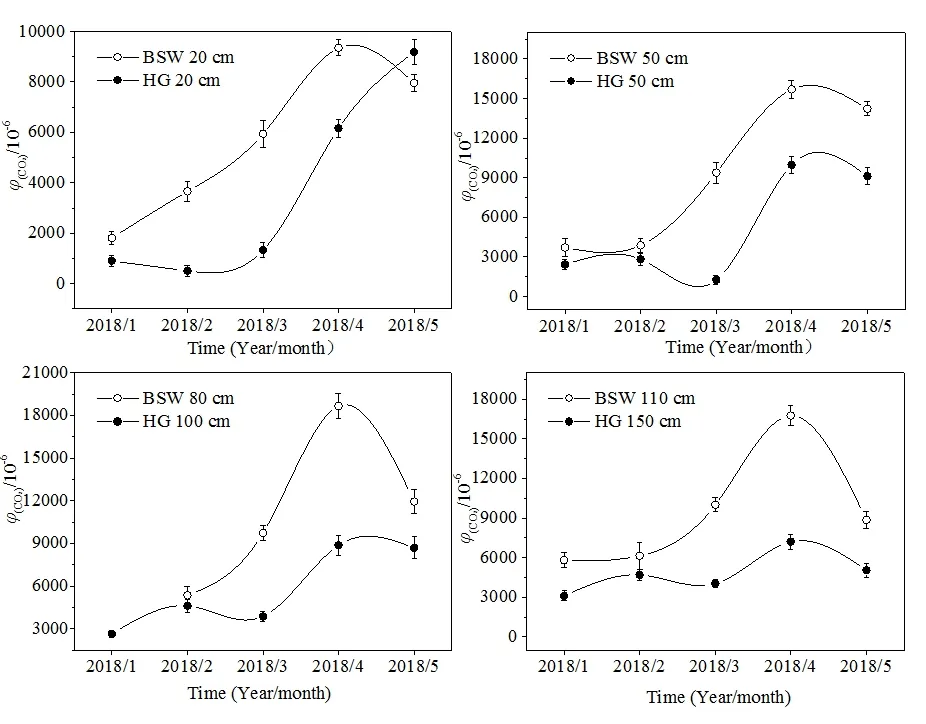
图2 柏树湾与后沟土壤CO2浓度对比Fig. 2 Comparison of soil CO2 concentration at BSW and HG
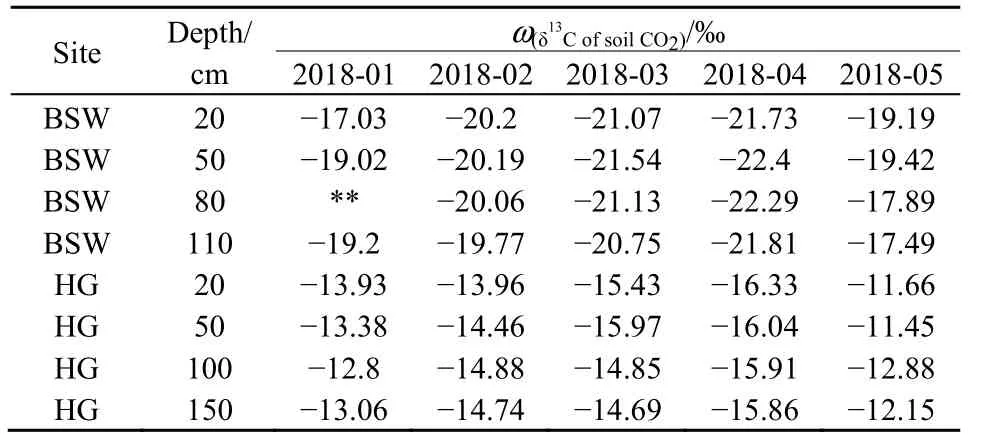
表2 不同时期柏树湾和后沟土壤CO2的δ13CTable 2 The δ13C of soil CO2 in different periods at BSW and HG
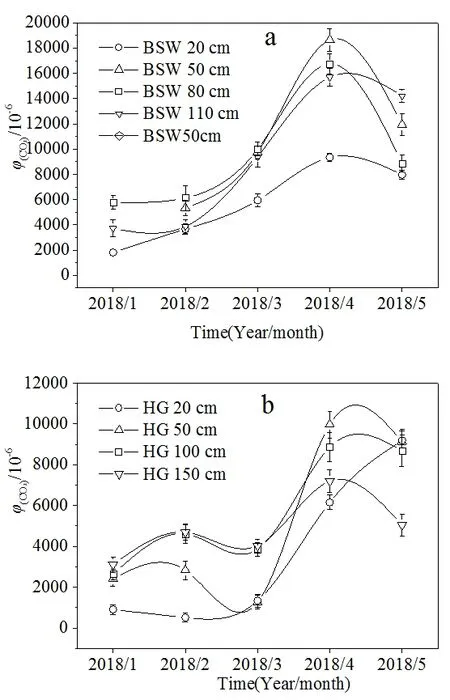
图3 土壤CO2浓度时间变化特征Fig. 3 Temporal variations of soil CO2 concentration
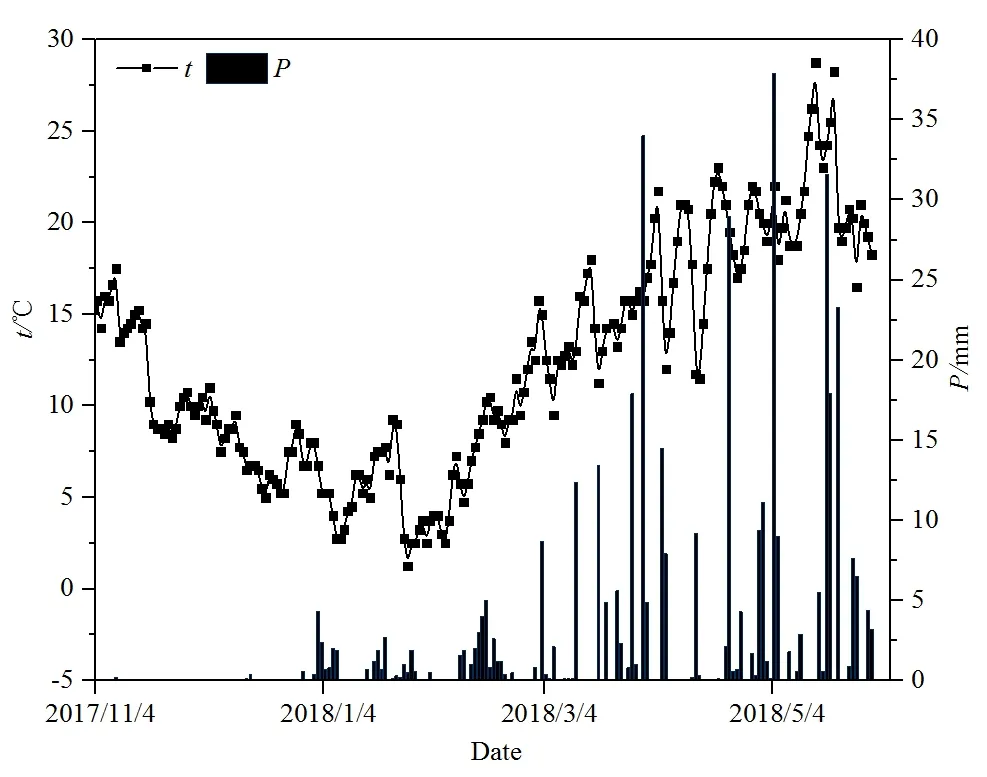
图4 气温(t)与降水量(P)Fig. 4 Temperature (t) and precipitation (P)
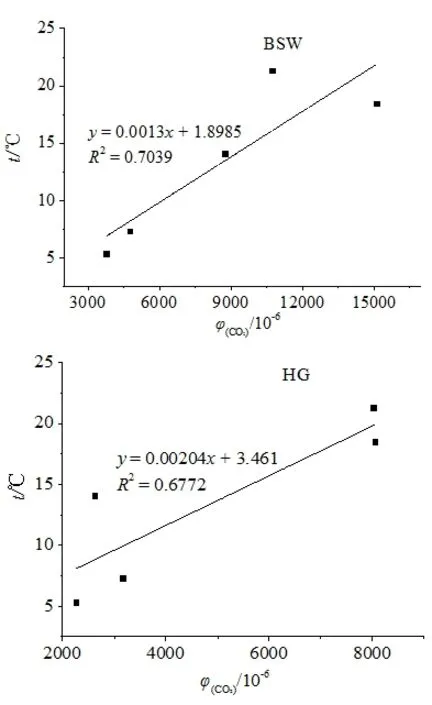
图5 气温与土壤CO2浓度相关性Fig. 5 Relationship between temperature and soil CO2 concentration
柏树湾各个深度的土壤CO2浓度在1—4月均呈现出逐渐上升的趋势,并在4月达到最大值(图3a)。后沟除20 cm处外,其他深度的土壤CO2浓度最大值也出现在4月(图3b)。研究期间气温逐渐升高(图4),气温升高一方面导致植物呼吸作用增强,根系向土壤中释放的CO2量增加,另一方面会刺激微生物活性,加强对土壤有机质和凋落物的分解。并且图5显示2个泉域土壤CO2浓度均与气温呈现一定的正相关性(图5),说明土壤CO2浓度的变化可能与气温有关。另外,不同深度土壤 CO2浓度变化范围的差异也说明了温度对土壤CO2浓度的重要影响。柏树湾20、50、80、110 cm处土壤CO2浓度变化范围分别为1812—9390、3716—15695、5338—18654、5795—16751 μmol·mol-1,其最大值分别为最小值的 5.16、4.22、3.49、2.89倍。后沟也表现出了相似的变化特征。其 20、50、100、150 cm处土壤 CO2浓度的变化范围分别为 507—9189、1264—9975、2630—8865、3105—7200 μmol·mol-1,最大值分别为最小值的 18.12、7.89、3.37、2.32倍。其他研究中也发现了相似的变化特征,即深部土壤CO2浓度变化具有一定的滞后性,而且随着土壤深度的增加,其滞后时间逐渐增长(Hendry et al.,1999;Bernhardt et al.,2006,Cannavo et al.,2006;Macpherson et al.,2008)。其原因可能是表层土壤对气温的响应较快,而深部土壤对外界环境变化的响应不及表层土壤敏感(Pavelka et al.,2007;殷超等,2017),从而导致了土壤CO2浓度升高的幅度随土壤深度的增加而减小。
然而,在本研究期间,5月气温最高,2个泉域土壤CO2浓度在5月却出现下降趋势(后沟20 cm处除外)。另外,与柏树湾CO2浓度逐渐上升不同,后沟50、100、150 cm处土壤CO2浓度在3月也出现了下降的趋势(图3b)。根据气象资料可以发现,在整个研究期间5月降水量最大,达到175.5 mm(图4)。尽管降水量增加可以促进植物根部呼吸作用和微生物活性,但降水量增加导致土壤含水量过高时,土壤中氧气的供应与扩散受到限制会对植物和微生物产生抑制作用,并导致土壤 CO2浓度降低(Liu et al.,2013;Gabriel et al.,2014)。另外,大量降雨入渗使土壤含水量饱和,形成壤中流补给坡面流或地下径流,径流量和径流速的增大加速了土壤 CO2随径流的迁移和循环,这也可能导致土壤CO2浓度出现陡然降低(蓝芙宁等,2017)。同理,后沟50、100、150 cm处土壤CO2浓度在3月出现降低的趋势可能也与3月降水增多有关。而柏树湾3月各深度的土壤CO2浓度并没有出现降低趋势。这可能是因为马尾松根系较发达,在经过长时间的干旱后,3月降水增加对根部呼吸的刺激作用大于其产生的抑制作用。并且发达的根系也导致土壤孔隙度较高,3月降水量不足以导致植物呼吸因缺氧而受到抑制。由于后沟表层土壤中混入的大量煤铁残渣使土壤孔隙度增加,土壤含水量对植物呼吸和土壤微生物产生的限制作用减弱。这导致后沟20 cm处土壤CO2浓度在3月和5月并没有出现降低的现象。并且5月后沟杂草生长旺盛,其较浅的根系集中在表层土壤中,因此后沟20 cm处土壤CO2浓度的最大值出现在5月。
3.3 土壤CO2垂向变化
除上述因素外,由于柏树湾和后沟均位于岩溶地区,土下碳酸盐岩溶蚀也可能会影响土壤CO2浓度。因此,本研究进一步分析了2个泉域土壤CO2浓度及其δ13C的垂向变化特征。
本研究期间,土壤CO2浓度在垂向上的单向梯度和双向梯度均被发现(图 6)。1—3月,柏树湾和后沟土壤CO2浓度均表现为单向梯度,最大值均出现在土壤底部;在 4、5月,柏树湾表现为双向梯度,土壤CO2浓度最大值分别出现在了80 cm和50 cm处(图6a);后沟4月出现了双向梯度,土壤CO2浓度最大值出现在50 cm处;5月表现为单向梯度,但最大值出现在20 cm处,即土壤CO2浓度随深度的增加而降低(图6b)。
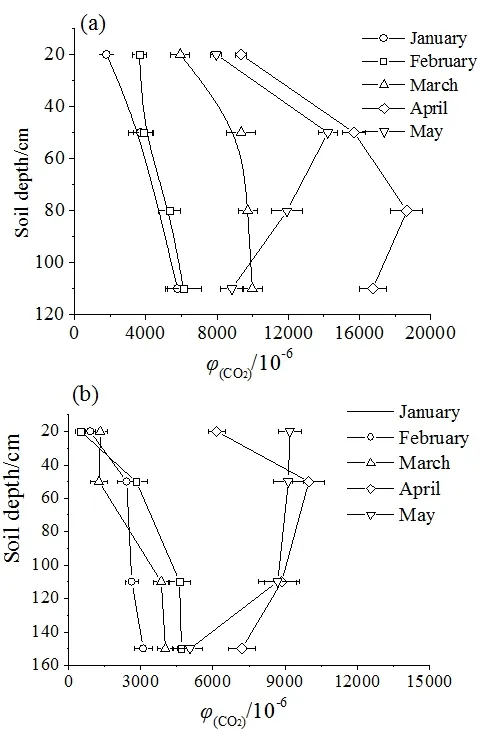
图6 土壤CO2浓度垂向变化Fig. 6 Vertical change of soil CO2 concentration
之前的研究曾发现柏树湾和后沟在旱季呈现双向梯度(Zhao et al.,2019),但本研究中1—3月柏树湾和后沟均呈现单向梯度,这可能与本研究期间降水量过少有关。如后沟在2017年11月—2018年2月以及2018年4月发生断流,断流时间较之前研究时期明显增加(Zhao et al.,2019)。缺少水分参与限制了土下岩溶作用对底部土壤 CO2的消耗,土壤CO2浓度垂向变化主要受气体传导率影响。由于表层土壤气体传导率较高,表层土壤中的CO2更容易通过土-气界面进入大气,而底部气体传导率较低,土壤CO2不容易扩散。因此,柏树湾和后沟土壤CO2浓度在垂向上的变化与非岩溶区相似,即随土壤深度的增加而升高。但随着降水增加,土下岩溶作用增强。岩溶作用对底部土壤CO2的消耗导致柏树湾和后沟底部土壤CO2浓度降低,土壤CO2浓度的最大值不再出现在土壤底部。值得注意的是,尽管3月降水量明显增加,达到了104.7 mm,但由于之前经历了长期的旱季,并且最大的降水事件出现在取样前几个小时,土壤CO2可能还未有充足时间参与岩溶作用,因此3月土壤CO2浓度依然表现为单向梯度。相反,尽管4月降水较少,但经历了3月后期较多的降水,土壤含水量增加,底部土壤CO2有充足时间参与岩溶作用,从而导致底部土壤CO2浓度降低。土壤CO2浓度在3月呈现单向梯度而在4月呈现双向梯度,表明降水通过岩溶作用对底部土壤CO2产生的影响具有一定滞后性。
之前监测结果显示,雨季后沟泉域 H2SO4和HNO3溶蚀碳酸盐后产生的CO2脱离了水体,并且土壤CO2浓度在雨季表现为单向梯度(Zhao et al.,2019)。为验证 H2SO4和 HNO3溶蚀碳酸盐岩后是否向土壤中释放了CO2以及其是否对土壤CO2浓度的垂向变化产生了影响,土壤 CO2的 δ13C用以示踪土壤CO2来源。H2SO4和HNO3参与岩溶作用产生的 CO2全部来自于碳酸盐岩。由于碳酸盐岩的δ13C远偏正于土壤有机质和植物的 δ13C(Jiang,2013),若 H2SO4和 HNO3溶蚀碳酸盐岩后释放的CO2若进入土壤,将导致底部土壤CO2的δ13C偏正。2个泉域深部(50 cm以下)土壤CO2的δ13C显示,除1月外,底部土壤CO2的δ13C出现了一定的偏正。但柏树湾110 cm处土壤CO2的δ13C偏正于80 cm处0.29‰—0.48‰,而后沟150 cm处土壤CO2的δ13C偏正于100 cm处0.05‰—0.73‰。相对于柏树湾,后沟H2SO4和HNO3输入量更大,后沟底部土壤CO2的δ13C偏正幅度却小于柏树湾。另外,不同深度土壤CO2的δ13C具有相似的季节变化特征(表2),说明不同深度的土壤CO2具有相同的来源。因此,之前发现的后沟土壤 CO2浓度在雨季呈现单向梯度,可能是降水下渗过程中将表层土壤CO2向下运移,使土壤CO2聚集在土壤底部造成的,而H2SO4和HNO3溶蚀碳酸盐对土壤CO2浓度的影响并不明显。后沟CO2脱离水体则是因H2SO4和HNO3溶蚀碳酸盐后产生的CO2以CO2(aq)形式进入地下水中,地下水出露后受 pCO2影响发生脱气造成的(West et al.,2005)。由于雨季水岩接触时间减少,H2SO4和HNO3溶蚀碳酸盐后产生的CO2更容易通过此过程脱离水体,这也解释了为什么在之前监测中发现雨季后沟CO2的净消耗量要小于旱季(赵瑞一等,2015b)。
4 结论
(1)上覆植被是2个泉域土壤CO2浓度出现差异的主要原因。柏树湾土壤 CO2浓度大于后沟,这可能是柏树湾泉域岩溶碳汇量大于后沟的重要原因。
(2)受温度影响,2个泉域土壤CO2浓度在研究期间呈现升高的趋势,并且土壤CO2浓度升高的幅度随土壤深度的增加而减小。同时,土壤CO2浓度也受降水量影响,降水过多可能会对土壤CO2产生抑制作用。
(3)降水量过少导致土下岩溶作用弱,土壤CO2浓度表现为单向梯度;降水量增加促进岩溶作用对底部土壤CO2的消耗,从而导致土壤CO2浓度表现为双向梯度。另外,土壤 CO2的 δ13C表明,上覆植被是土壤CO2的主要来源,H2SO4和HNO3溶蚀碳酸盐岩产生的CO2对土壤CO2的贡献较小,可以排除其对土壤CO2浓度垂向变化的影响。
