黄河三角洲潮坪环境中现代生物遗迹组成与分布特征
2020-01-18王媛媛王学芹胡斌
王媛媛,王学芹,胡斌
1.河南理工大学资源环境学院,河南焦作 454003
2.河南省生物遗迹与成矿过程重点实验室,河南焦作 454003
0 引言
现代沉积作用是一系列物理、化学及生物三者相互作用相互影响的复杂过程,是多种地质作用的产物。现代沉积中的现代生物遗迹对解读古土壤、古环境具有一定的启发性[1-6]。然而前人的研究大都着重于物理因素作为重要的沉积学标志,而忽略了生物成因沉积构造对沉积环境的解释作用。遗迹学研究的是现代和古代生物生活时在沉积物底质中(层面和层内)遗留下来的活动遗迹[7-8]。这种生物遗迹与其沉积底质、沉积环境、生态环境等有着密切的关系,对解释沉积环境有很大的启示意义[9-13]。
近十年来,国外学者对潮坪沉积中的现代生物遗迹进行了较为深入的研究。通过研究潮坪沉积中的现代遗迹,初步总结了潮坪沉积中的生物遗迹的形态、行为习性、分布特征,底层和环境因素等对生物遗迹分布的影响,并应用于相类似遗迹化石的对比,讨论了现代生物遗迹对潮坪沉积环境中微环境的指示和鉴定作用[14-18]。国内近几年也开展了现代生物遗迹的初步研究[19-23]。然而我国潮坪沉积现代生物遗迹的研究还处于起步阶段,并且有关黄河三角洲潮坪中现代生物遗迹,前人做的工作极少,而且从研究广度和深度方面还有待提高。
本次工作主要研究黄河三角洲潮坪环境现代生物遗迹的形态和行为习性特征,以及不同的微环境中现代生物遗迹的组成与分布的差异性,并研究其与沉积底层、沉积环境的关系,总结现代生物遗迹在潮坪环境中不同微环境的分布模式。
1 研究区概况

图1 研究区位置图和采样点(a)研究区在中国的位置;(b)研究区域;(c)潮坪环境的采样点;①潮上带;②泥坪;③混合坪;④砂坪Fig.1 Study area and sampling site
研究区位于渤海湾西岸黄河三角洲潮坪环境(图1)。黄河三角洲是受潮水影响的河控三角洲,属于半干旱季风气候[24]。该区域同时受河流、潮汐的影响,有机质较丰富,因此生物多样性很丰富。黄河是渤海西海岸主要补给性河流,流向为东西方向,潮汐为半日潮型。其全年温度一般为-8°C~36°C,平均年降水量大约为600 mm。按受潮水作用的不同,开发利用的差异,潮坪环境分为潮上带,泥坪,混合坪,砂坪和潮下带[24]。潮上带位于高潮位以上,沉积物主要是黄灰色泥质粉砂和一些生物碎屑,土壤熟化程度较高,有机质含量比潮间带高,常见植物残体。泥坪位于潮间带上部,在高潮线与平均高潮位之间,沉积淤泥质黏土和黏质砂土。混合坪位于平均高潮位线与平均低潮位线之间,沉积物主要是淤泥夹粉砂,上部淤泥层较厚,向下随着淤泥含量减少而粉砂增加。砂坪位于平均低潮位至低潮位线之间,上部沉积物为黏土质粉砂,中下部逐渐过渡为粉砂,有的地区甚至出现细砂、极细砂底质。潮下带指位于平均低潮线以下、浪蚀基面以上的浅水区域。此区域始终处于水下。
黄河三角洲现代沉积中发育丰富的沉积构造类型,反映其沉积环境和沉积过程的复杂多样性。黄河三角洲潮坪地形平缓,风的作用较大,潮坪环境的波痕发育,主要有削顶波痕,双脊波痕,舌形波痕和干涉波痕(图2)。
2 研究方法
2.1 取样
经过野外勘察之后,根据不同沉积微环境选择 4个不同的采样点(① 118°94′E,37°57′N;②118°95′E,37°57′N;③118°96′E,37°57′N;④118°97′E,37°56′N)(图1c)。退潮后,观察层面的生物遗迹和垂直潜穴的形态,种类。对不同微环境的沉积物采样,每个采样点采集离层面20 cm深的样品,并由采样点中心与周围5个点的样品混合而成,一般为350 g左右。选择形态完整的,具有代表性的生物潜穴,用PVC管取芯。所用PVC管的直径有两种:3.2 cm和7.5 cm,其高度为10 cm。

图2 黄河三角洲潮坪中常见的波痕类型。(a)双脊波痕;(b)削顶波痕;(c)舌形波痕;(d)干涉波痕Fig.2 Common ripple mark in the tidal flat of the Yellow River delta
2.2 激光粒度分析
采集的沉积物样品带回实验室,先用烘箱在105℃下干燥24小时,将干燥后的沉积物按照四种不同的微环境装入保鲜袋内,每带样品50 g左右,用Rise 2008激光粒度分析仪做粒度分析实验。样品通过仪器的分选,得到多个粒度曲线,通过软件PADMAS颗粒粒度测量分析系统(Particle Diameter Measure&Analysis System)将测试数据做平均、统计、比较和模式转换等处理,得到分析结果(表1、图3)。
图3中,左边纵坐标为微分分布,右边纵坐标为累积分布,横坐标为颗粒物直径。微分分布又称为区间分布或频率分布,它表示一系列粒径区间中颗粒的百分含量。累计分布也叫积分分布,它表示小于或大于某粒径颗粒的百分含量。D50:一个样品的累计粒度分布百分数达到50%时所对应的粒径。它的物理意义是粒径大于它的颗粒占50%,小于它的颗粒也占50%,D50也叫中位径或中值粒径。D50常用来表示粉体的平均粒度。根据实验结果,得出以下结论:潮下带的沉积物最粗。潮下带的能量最强,水动力很大,研究的时候也最难。通过比较潮上带,泥坪,混合坪,砂坪和潮下带的沉积物粒度,可以发现从潮上带到潮下带,沉积物粒度逐渐增大(图3e),说明水动力状况也逐渐增强。
2.3 CT扫描
将取得的岩芯带到胜利油田(地化实验室)做CT扫描实验。CT扫描参数为:仪器型号是NanoVoxel 2000,电压150 keV,电流30 mA,因为没有用CCD相机,所以没有用物镜。用平板探测器扫描,旋转360°,单次采集1 440张,曝光时间,因样品而异,一般每秒钟拍n张,n张图片取平均值合成1张,n一般在6~10之间。分辨率73 000/850=85.88µm。
2.4 三维重构
获得的体数据(CT图像数据)后期三维可视化处理,包括体数据三维渲染、灰度值分割、定向数字虚拟切面、动画制作等使用德国Volume Graphics公司出品的三维可视化软件VGstudio Max2.1完成[25]。重构实验室为中国科学院南京古生物地质研究所X射线断层扫描实验室。
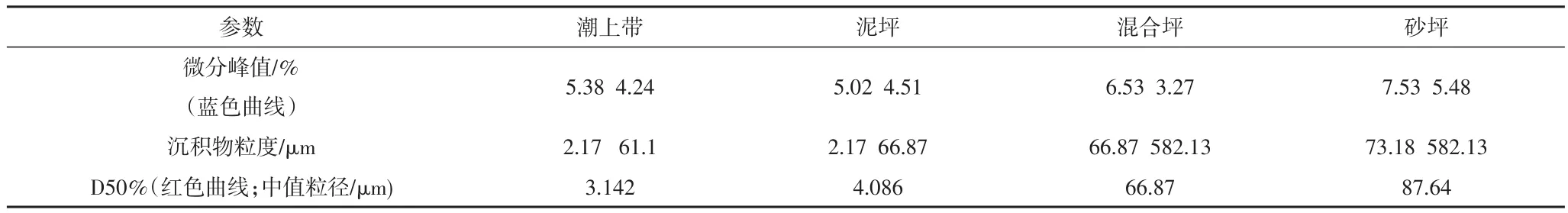
表1 四个沉积微环境的沉积物粒度的比较Table 1 Comparison of sediment grains of four sub-environments

图3 不同的沉积环境粒度分析曲线图(a)潮上带;(b)泥坪;(c)混合坪;(d)砂坪;(e)不同沉积环境的粒度分析比较。在图e中,黑色曲线是潮上带;红色曲线是泥坪;蓝色曲线是混合坪;绿色曲线是砂坪Fig.3 Grain size distribution in different sedimentary environments

图4 潮上带日本大眼蟹潜穴层面概况(a)潮上带沼泽;(b)层面上日本大眼蟹潜穴形态概览Fig.4 The overview of burrows produced by Macrophthalmus japonicus in supratidal zone
3 黄河三角洲潮坪现代生物遗迹
3.1 潮上带生物遗迹
潮上带只有在特大高潮时才会被淹没,常形成沼泽、盐湖(图4a),裸露着松散的粉砂,黏土,主要的生物遗迹是日本大眼蟹的潜穴和爬行迹,潜穴形态较单一,爬行迹模糊不清(图4b),与泥坪中的部分日本大眼蟹遗迹类似,因此会在潮间带的泥坪中统一描述。
3.2 潮间带生物遗迹
3.2.1 泥坪生物遗迹
①日本大眼蟹Macrophthalmus japonicus营造的遗迹
日本大眼蟹Macrophthalmus japonicus主要分布在潮上带,泥坪和混合坪,并且泥坪中的日本大眼蟹数量最多。退潮后外出活动,具群聚性。日本大眼蟹造的遗迹主要是层面爬行迹,居住潜穴和排泄迹。爬行迹的形态主要有两种,一类是呈U形带状,两侧是轻微突起的砂脊,中间不平坦,有一条轻微突起的砂脊,爬迹两侧外缘有螃蟹的爪印(图5a)。另一类是紧凑的四列带状爬行迹,不连续,每隔3 mm断开,为螃蟹螯足留下的痕迹,略成弧形(图5b)。
日本大眼蟹的居住潜穴形态多样,穴口与层面垂直或斜交,由于沉积物含水率不同,粒度不同,潜穴口形态不一,一般为放射状,近同心圆状,鸟尾状,土丘状,围墙状。退潮后,日本大眼蟹不断从潜穴内向外探视,并爬出潜穴觅食,稍有动静就迅速钻入潜穴内,反复地探视和钻入潜穴,形成了线形放射状爬行迹。其长为5~25 cm,宽为1~3 cm(图5c),与生物大小有关。在含水率适中的沉积底层中,放射状爬迹清晰,在含水率较高的沉积底层中,放射状爬迹模糊不清(图5d)。在潮道两侧,围墙状的潜穴较多,潜穴口为圆形,直径为2~3 cm,围墙高为3~5 cm(图5i),造迹生物通过黏液瘤状构造用以加固潜穴,避免潜穴坍塌。海水退去,沉积物表层留下大量藻类和有机质,日本大眼蟹爬出潜穴摄食,产生很多食渣砂球,砂球大小不一,直径为5~10 mm。砂球有时分布较散乱,无规律性,有时排列成近同心圆形(图5e),主要是为了吸引异性进行交配。鸟尾状潜穴在泥坪中也很常见。日本大眼蟹爬出潜穴活动,从潜穴中挖出泥质沉积物。螃蟹数量较多,爬行过程中在泥质沉积物表面留下爪痕(图5f)。在平坦的泥坪上,发现较多小土丘状的潜穴,层面上为圆形开口,直径为5~10 mm(图5h),筑造这类潜穴的螃蟹个体较小,为了防止敌人侵袭。日本大眼蟹的潜穴内部构造较复杂,有Y形,U形,J形等管状潜穴,穴壁光滑,与表层沉积物颜色相同(图5g)。潜穴直径为2~4 cm,主穴道的穴口直径稍大,深度延伸20~40 cm。通过对日本大眼蟹潜穴进行CT扫描和三维重构,其形态结构更加直观和形象。从日本大眼蟹三维图像看出,潜穴形态为Y形,I形,U形,多分叉形,黄色的I形潜穴,上下直径不等,为3~9 mm,上部直径较大,呈喇叭状开口;绿色的U形潜穴基本等径,直径约为4 mm,U形竖直高度约为15 mm;紫色的Y形潜穴基本等径,直径约为3 mm,Y形整体与水平面斜交。细小的棕色潜穴为沙蚕幼虫潜穴,较分散,直径为1~2 mm,长为3~8 mm(图6a)。绿色分叉形潜穴主干直径稍大,为3.5 mm,分支直径为2.5~3 mm,有2~3个开口,主干开口最大,直径约为7 mm(图6b)。多分叉潜穴周围有细小的紫色潜穴,较密集,为沙蚕的潜穴,沙蚕寄居在螃蟹潜穴中,进食螃蟹潜穴中的有机质,螃蟹的潜穴也是砂蚕的临时庇护所。沙蚕潜穴有I形,弯曲型,长为2~17 mm,直径为1~3 mm。
②泥螺Bullacta exarata营造的遗迹
泥螺Bullacta exarata主要分布在泥坪和混合坪沉积物表面,摄食藻类和有机碎屑。退潮后,大量泥螺在沉积物表面缓慢爬行,有时也会隐藏于层面以下1~2 cm深处。泥螺爬行时在沉积物表面留下拖迹,为条带状痕迹,为直型、微弯曲型、交叉弯曲型,长为15~30 cm,宽为0.5~1 cm,因泥螺大小而异。在含水率适中的底层中,拖迹较浅,中间较平滑,两侧形成稍高于表面的砂脊(图7a)。一般来说,泥螺喜欢含水率高的沉积底层,其爬行留下的遗迹很难保存下来。退潮后,大量生物出来活动,一些鸟类也会在泥坪觅食,因此,为了自我保护,泥螺用头盘掘起泥砂与身体分泌的黏液混合,包被在身体表面,就像一个泥团(图7b)。
③红带织纹螺Nassariussuccinctus营造的遗迹
红带织纹螺Nassariussuccinctus主要分布在泥坪中,涨潮时潜入沉积物表面以下2~3 cm深处;退潮后,爬出沉积物表面进行觅食,留下觅食迹。觅食迹为U形带状遗迹,弯曲型,有多个转弯,多个爬行迹交叉缠绕,宽为0.5 cm,长为15~35 cm。由于红带织纹螺个体较小,即使成虫个体也很小,而且相对泥坪上的螃蟹和跳跳鱼等生物来说,数量也很少,因此在野外不容易观察。在含水率适中的底层中,遗迹中间留下清晰的蹼状构造(图8a),在含水率较高的底层中,两侧有明显的凸起的砂脊,中间的蹼状构造模糊不清(图8b)。
④跳跳鱼Periophthalmus cantonensis营造的遗迹

图5 日本大眼蟹营造的遗迹(a)日本大眼蟹的爬行迹及造迹生物;(b)日本大眼蟹的足辙迹;(c)固底中日本大眼蟹的星射状遗迹;(d)汤底中日本大眼蟹的星射状潜穴;(e)日本大眼蟹的粪球粒;(f)日本大眼蟹的鸟尾状遗迹;(g)纵剖面上日本大眼蟹的潜穴形态;(h)成对的土丘状日本大眼蟹潜穴;(i)围墙状日本大眼蟹潜穴Fig.5 Trace produced by Macrophthalmus japonicus
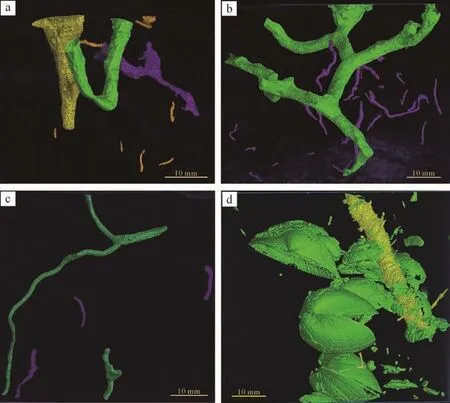
图6 双齿围沙蚕、日本大眼蟹、四角蛤潜穴的三维形态(a)日本大眼蟹的U形,I形,Y形潜穴形态;(b)日本大眼蟹的多分叉形潜穴;(c)双齿围沙蚕的Y形潜穴;(d)四角蛤的倾斜潜穴Fig.6 Three-dimensional morphology of burrow of Macrophthalmus japonicus,Perinereis aibuhiteusis and Mactra veneriformis Reeve

图7 泥螺营造的遗迹(a)泥螺的拖迹(黄色箭头指示的为造迹生物:泥螺);(b)泥螺头盘掘起泥砂包被在表面(黄色箭头指示的为造迹生物:泥螺),右上角为造迹生物Fig.7 Trace produced by Bullacta exarata
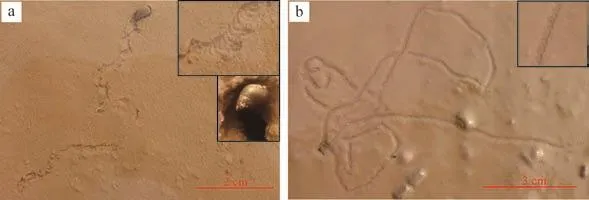
图8 红带织纹螺营造的遗迹(a)带蹼状构造的红带织纹螺觅食迹(右上角为蹼装构造及造迹生物);(b)不带蹼状构造的红带织纹螺觅食迹Fig.8 Trace produced by Nassarius Succinctus
弹涂鱼(Periophthalmus cantonensis)主要分布在泥坪中,营造的遗迹主要是层面的爬行迹和居住潜穴。退潮后,跳跳鱼在层面上爬行、跳跃。当跳跳鱼运动时,在层面上留下一条拖痕,拖痕为犁沟状,长20~90 cm,宽2~5 mm。拖痕两边为对称的弧形痕迹,是生物的胸鳍和尾鳍留下的,外侧是胸鳍痕迹,胸鳍压痕中留下明显的纹理(图9c),长0.5~1.5 cm,宽0.3~0.5 cm;内侧是尾鳍痕迹,长0.2~0.5 cm,宽0.1~0.3 cm。当沉积底层粒度较细、含水率适中时,其痕迹清晰;当沉积底层含水率较高时,痕迹模糊(图9a)。跳跳鱼的潜穴形态一般是I形或U形,有一两个潜穴口,穴口倾斜,附近有跳跳鱼的爬迹(图9b),潜穴内部有分叉的通道,短的为死通道。研究表明,潜穴构造与跳跳鱼的大小有密切的关系,跳跳鱼将潜穴作为觅食、居住、逃逸以及产卵的场所[26]。跳跳鱼进入潜穴的身体方向有一定的意义,比如,跳跳鱼的头先进入潜穴,表明潜穴为逃逸潜穴,如果尾巴先进入潜穴,表明潜穴为觅食潜穴[27-28]。由于沉积物较软,其潜穴一般难以观察和保存。当弹涂鱼稍受惊动,马上跳回水中或躲进螃蟹潜穴中。因此螃蟹的潜穴和跳跳鱼的潜穴很难区分,不过,一般来说,潜穴口周围有跳跳鱼的爬行迹,即为跳跳鱼的潜穴,否则为螃蟹的潜穴。
3.2.2 混合坪生物遗迹
①双齿围沙蚕Perinereisaibuhitensis营造的遗迹
双齿围沙蚕Perinereisaibuhitensis(Grube)主要分布在混合坪中,幼虫以浮游生物为食,成虫以腐殖质为食。沙蚕造的遗迹主要有觅食迹、居住迹和排泄迹。觅食迹的形态为直型,弯曲型。长为3~10 cm,宽为1~2 mm,觅食迹两端一般与两个潜穴口相连。如果砂蚕的觅食迹较密集,它们可能会相交。在爬行过程中,沙蚕的疣足可能留下细小的足迹。在含水率适中的沉积底层中,两侧的沙脊底部出现疣足的痕迹(图10d);在含水率较高的沉积底层中,沙蚕疣足的痕迹模糊不清(图10a)。居住迹主要是垂直管状的居住潜穴,潜穴口在表层呈圆形或次圆形,直径1~2 mm,觅食时沙蚕伸出穴口,摄食层面上的藻类、有机碎屑,稍有动静立即钻入潜穴,沙蚕反复伸出穴口觅食,在穴口形成几条长度不一的痕迹,长1.5~3 cm,沿穴口呈星射状分布,形成星射状觅食迹(图10c)。潜穴口周围有粪球粒(图10e),粪粒直径1~2 mm左右,呈圆锥状粪丘分布于穴口周围。潜穴形态为Y形,U形,潜穴壁较光滑(图10f,g),等径,直径1~2 mm,下潜深度为5~8 cm。沙蚕有时会寄居在螃蟹潜穴内,推测是由于在爬行过程中,突遇敌人躲藏在螃蟹潜穴内。或在沉积物表面觅食时,由于层面有机质较少,钻进螃蟹潜穴内进行觅食(图10b)。通过对双齿围沙蚕潜穴做CT扫描及三维重构,可以清晰地观察到沙蚕层内潜穴的三维形态,为Y形,I形(图6c),较大的绿色Y形潜穴为成虫潜穴,上部U形与下部弯曲型柱状基本等径,直径约为2 mm,下方弯曲型柱体延伸较深,约为55 mm。较小绿色Y形,紫色I形为幼虫潜穴,较小绿色Y形潜穴直径约为3 mm,整体较短,向下延伸至10 mm;紫色潜穴较小,直径为1~3 mm 不等,长为8~12 mm 不等,较分散。

图9 跳跳鱼营造的遗迹(a)汤底中跳跳鱼的爬行迹;(b)跳跳鱼的潜穴;(c)固底中跳跳鱼的遗迹(右上角为造迹生物)Fig.9 Trace produced by Periophthalmus cantonensis

图10 双齿围沙蚕营造的遗迹(a)汤底中双齿围沙蚕的爬行迹;(b)螃蟹潜穴口旁边的双齿围沙蚕的爬行迹;(c)双齿围沙蚕的星射状觅食迹;(d)弯曲型双齿围沙蚕的爬行迹(右上角为造迹生物);(e)双齿围沙蚕潜穴口旁的粪球粒(右上角为粪球粒);(f)双齿围沙蚕的U形潜穴;(g)双齿围沙蚕的Y形潜穴Fig.10 Trace produced by Perinereis aibuhitensis
②托氏昌螺Umboniumthomasi营造的遗迹
托氏昌螺分布在潮间带混合坪中,退潮后在层面上爬行,有时聚集成群,食藻类。托氏昌螺的遗迹主要是运动迹。研究区观察到的托氏昌螺壳体大小1~2 cm,其运动迹为条带状爬行迹,长为3~9 cm,宽为1 cm,两侧为稍高于沉积物表面的砂脊,中间有一条极细的沙脊。在含水率较低的底层中,运动迹表面出现规则的羽状纹理,沿中间的细沙脊呈对称分布,纹理与两侧沙脊成一定角度,大约为30°(图11a)。在含水率较高的底层中,运动迹表面较光滑,无纹理(图11b)。
③四角蛤Mactra veneriformisReeve营造的遗迹
四角蛤(Mactra veneriformisReeve)常潜于沉积物表层往下5~6 cm深处,以海水中的浮游生物,藻类和各种残渣为食。主要栖息于潮间带混合坪和砂坪,底内穴居。四角蛤造的遗迹主要是居住迹,潜穴与层面垂直,由两部分组成:上面的虹吸管穴道和下面的壳体潜穴。从沉积物表面看,四角蛤潜穴为圆形,直径为2~5 mm,虹吸管潜穴近似圆柱形,长2~3 cm,直径约0.3 cm。下面的壳体潜穴长3.1~3.3 cm,宽0.5~1 cm,表面光滑(图12a)。有时潜穴轻微倾斜,不与层面垂直(图12b)。这一点与四角蛤层内潜穴的三维图像恰好对应(图6d),黄色倾斜柱体为四角蛤潜穴,长为35 mm,直径为4.5 mm,周围的绿色壳体已钙化,并且可以观察到托氏昌螺的壳体,表明托氏昌螺和四角蛤栖息在同一沉积环境中。从底层表面的潜穴开口来看,四角蛤潜穴与双齿围沙蚕的潜穴有明显差异,一般来说,四角蛤潜穴在沉积底层表面的开口为单孔,直径约3 mm,潜穴口周围没有粪球粒。而双齿围沙蚕潜穴一般较密集,且在沉积物表面的开口明显小于四角蛤的穴口,直径约1~2 mm,部分穴口处有粪粒或星射状分布的觅食拖迹。
④豆形拳蟹Philyra pisumde Haan营造的遗迹
豆形拳蟹Philyra pisumde Haan主要分布在砂坪中,数量较少,以底栖生物和泥砂中的有机质为食。涨潮时下潜到沉积物以下,退潮后爬出层面觅食,豆形拳蟹造的遗迹主要是居住迹和爬行迹。退潮后,豆形拳蟹在沉积物表层活动,形成三列线形爪痕,长为10~35 cm,三列爪痕紧凑,不连续,由螯足的凹坑隔开,近似等间距,断开的线形痕迹长为1~2 cm,宽为3 mm(图13b)。居住潜穴穴口与底表斜交,周围有零星食渣砂球和爪痕,砂球直径为2~5 mm,有的被潮水冲刷崩塌。潜穴内部构造复杂,有Y形,J形,U形等,延伸15~30 cm,穴壁光滑,有时有食渣砂球。豆形拳蟹的居住潜穴与日本大眼蟹的居住潜穴有一定的区别。豆形潜穴的潜穴口形态较单一,食渣砂球也较分散,由于砂坪受潮水影响较大,沉积物粒度较粗,活动留下的爪痕和爬行迹模糊不清(图13a)。

图11 托氏昌螺营造的遗迹(a)有纹理的托氏昌螺的爬行迹(右上角为造迹生物);(b)无纹理的托氏昌螺的爬行迹Fig.11 Trace produced by Umbonium thomasi

图12 四角蛤营造的遗迹(a)垂直层面的四角蛤潜穴(黄色箭头指示潜穴);(b)斜交层面的四角蛤潜穴(黄色箭头指示潜穴)Fig.12 Trace produced by Mactra veneriformis Reeve
3.2.3 砂坪生物遗迹
①竹蛏Solen gouldi营造的遗迹
竹蛏(Solen gouldi)主要分布于砂坪中沉积物以下3~8 cm深处,以其锚形斧足营直立生活。竹蛏造的遗迹主要是层内居住迹,为垂直潜穴,在沉积底层表面有开口,穴口直径约2 mm左右,无堆积物。潜穴上下等径,纵切面与壳体宽度和高度一致(图14a),潜穴下潜深度为10~15 cm,穴壁较光滑。竹蛏居住潜穴与底层表面近似垂直,整体形态为“I”形(图14b)。穴道横切面与生物横切面形状一致,一般宽约1.5 cm,厚约1 cm,壳体穴道长度大于壳体长度,一般长度约6~8 cm。
②棒锥螺Turritella bacillumKiener营造的遗迹棒锥螺Turritella bacillumKiener分布于砂坪中,以藻类为食。退潮后,在沉积物表面缓慢爬行进行觅食,觅食迹形态一般为直线型或微弯曲型(图15a)。一般来说,拖迹宽为3~10 mm,长为10~20 cm。拖迹的密度随动物觅食活动分散或密集。由于生物的壳体较尖锐,运动的时候,随着壳体方向变化而变化,壳尾位于两侧砂脊中间,形成较深的犁沟状痕迹,两侧砂脊倾斜向上,中部深深下凹。
③秀丽织纹螺Nassauriusfestiva营造的遗迹
秀丽织纹螺Nassauriusfestiva分布在潮间带砂坪中,易群聚,以沉积物表面的藻类为食。秀丽织纹螺造的遗迹主要是运动迹。运动迹形态为不规则的U形带状迹,为直型,弯曲型,交叉型,宽为5~10 mm,长为5~20 cm。遗迹中间平坦光滑,两侧为稍高于中间的沙脊(图15b)。秀丽织纹螺的觅食迹与泥螺的爬行迹形态类似,但是秀丽织纹螺的觅食迹比泥螺的爬行迹浅。通过多次的野外观察发现,秀丽织纹螺生活在含水率较高的沉积底层中,并且,砂坪比泥坪的沉积物粒度粗,因此留下的痕迹较浅,不清晰。

图13 豆形拳蟹营造的遗迹(a)豆形拳蟹的潜穴;(b)豆形拳蟹的爬行迹(右上角为造迹生物)Fig.13 Trace produced by Philyra pisum de Haan

图14 竹蛏的潜穴(a)竹蛏的潜穴(黄色箭头指示造迹生物:竹蛏);(b)纵剖面上的竹蛏潜穴Fig.14 Burrows produced by Solen gouldi

图15 棒锥螺和秀丽织纹螺营造的遗迹(a)棒锥螺的爬行迹(右上角为造迹生物);(b)秀丽织纹螺的爬行迹(右上角为造迹生物)Fig.15 Traces produced by Turritella bacillum Kiener and Nassaurius festiva
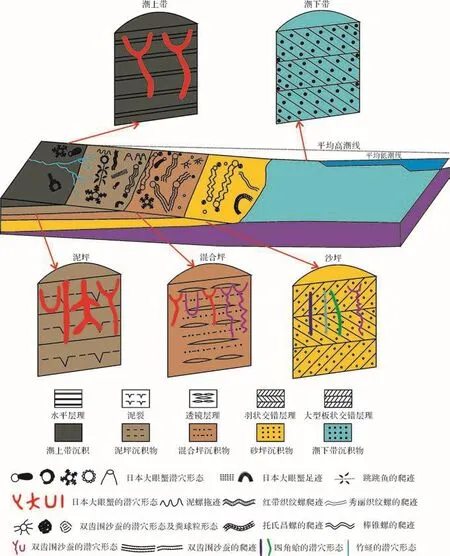
图16 黄河三角洲潮坪环境现代生物遗迹组成及分布模式图Fig.16 Composition and distribution mode of neoichnological assemblage in tidal flat of the Yellow River delta
3.3 潮下带生物遗迹
潮下带始终处于水下,为高能的环境,受潮水影响较大,生物遗迹也极少(图16)。另外鉴于观测不便,本文不做描述和研究。
4 结论
综上所述,黄河三角洲潮坪环境中现代生物遗迹在潮间带的分布丰度和分异度最高。潮上带底层沉积物以泥质为主,分选好,位于平均高潮线以上,受潮水影响较小,有机质较少,所以生物遗迹较少,主要是日本大眼蟹Macrophthalmusjaponicus的层面觅食迹和层内U形,Y形及多分叉的居住潜穴。潮间带中的泥坪沉积物为泥质黏土,分选较好,受河流和潮水的共同影响,营养物质较丰富,生物遗迹较丰富,有日本大眼蟹Macrophthalmusjaponicus的层面觅食迹和层内U形,Y形居住潜穴;泥螺Bullacta exarata的层面拖迹;红带织纹螺Nassarius Succinctus的层面觅食迹;跳跳鱼Periophthalmus cantonensis的层面爬行迹和居住潜穴;双齿围沙蚕Perinereis aibuhitensis的层面爬行迹,觅食迹和垂直的Y形、U形居住潜穴;混合坪沉积物为泥质和砂质的混合组分,分选中等,生物遗迹比泥坪要少,主要是双齿围沙蚕Perinereis aibuhitensis的爬行迹和居住迹;豆形拳蟹Philyra pisumde Haan的层面爬行迹和居住潜穴;托氏昌螺Umbonium thomasi的层面爬行迹;四角蛤Mactra veneriformisReeve的垂直居住潜穴。砂坪沉积物较粗,为黏土质粉砂,细砂,分选较差,水动力较强,生物遗迹较少,主要是棒锥螺Turritella bacillumKiener的层面爬行迹,秀丽织纹螺Nassauriusfestiva的层面爬行迹;竹蛏Solen gouldi的垂直居住潜穴。潮下带始终处于水下,水动力较强,无法观察生物遗迹。
黄河三角洲潮坪沉积中的现代生物遗迹的组成与分布与沉积底层的粒度,含水量,水动力条件及食物来源等有着密切的关系,造迹生物灵敏的反应了这些沉积环境和生态环境指标,营造了不同形态和生态属性的生物遗迹。潮上带受潮水影响较小,营养物质较少,生物遗迹种类较少。潮下带始终处于水下,为高能的环境,受潮水影响较大,生物遗迹也极少。而潮间带环境中的生物遗迹的丰度和分异度最高,保存潜力极高。其沉积物粒度中等,水动力条件适宜,又有丰富的有机质和食物来源,从泥坪到混合坪再到砂坪,生物遗迹的丰度和分异度呈逐渐减少的趋势。
致谢 感谢中石化胜利油田勘探开发研究院地化实验室在CT扫描中的帮助,感谢中国科学院南京古生物地质研究所殷宗军副研究员,吴素萍老师在三维重构中的帮助,感谢波兰雅盖隆大学地质科学研究所的Alfred Uchman教授,瑞士巴塞尔大学地质研究院Andreas Wetzel教授及南京古生物地质研究所的殷宗军和罗茂副研究员提供的具有启发性的意见和建议。
