东海带鱼资源变动及其栖息地驱动因子研究进展
2020-01-16陈全震李尚鲁颜云榕叶文建俞存根
杜 萍,陈全震,李尚鲁,颜云榕,叶文建,俞存根
(1.国家海洋局海洋生态系统与生物地球化学重点实验室//2.自然资源部第二海洋研究所,浙江 杭州 310012;3.浙江海洋大学,浙江 舟山 316022;4.广东海洋大学,广东 湛江 524000;5.浙江省海洋监测预报中心,浙江 杭州 310007)
带鱼(Trichiurus lepturus)是世界重要的渔获种类,主要分布在西北太平洋、印度洋和大西洋。带鱼分布区内,以我国产量最高[1]。带鱼广泛分布于我国四大海区,是我国近海最重要的一种传统底层经济鱼类。全国带鱼年产量变动趋势为1956-l974年为迅速增长期,从17×104t增至58×104t;1974-1988年在波动中下降,至1988年仅为37×104t;20世纪90年代开始又快速增长,1995年突破100×104t,2006年达到历史最高记录142×104t;之后10年保持在110 × 104t左右,2017年、2018年较低,分别为101× 104t、94 × 104t[2-3]。根据带鱼地理分布和形态特征,一般将分布在我国的带鱼分成黄渤海种群、东海种群和南海种群,但仍有分歧。其中,南海种群资源数量调查研究数据不足;黄渤海种群已于20世纪60年代严重衰退;东海种群是分布在我国的带鱼主群,冬汛嵊山渔场曾是我国最大的带鱼生产渔场和渔汛区[4-8],即东海是带鱼主要产区,产量约占全国总产量的87%。
带鱼于20世纪30年代随着嵊山渔场的开发迅速跻身东海“四大渔业”行列;东海带鱼产量在20世纪50年代初至1974年间一直呈上升趋势,1974年达到最高,年产量为53 × 104t;之后波动下降,1988年仅29 × 104t;此后由于捕捞能力快速增长,带鱼捕捞产量大幅增加,2000年达到91 × 104t;2000年后又开始波动下降,但一直能维持渔业生产,2018年上海、浙江和福建3省的捕捞产量和约为53 × 104t[3,9](20世纪80年代后我国带鱼统计数据中有时包含来自东南亚和西非沿岸国家底拖网的带鱼,因此产量统计数据可能会偏高)。带鱼作为东海“四大渔业”之一,也是我国单鱼种产量最高的一种经济鱼类,无论是过去还是现在,对东海乃至全国海洋渔业生产都具有十分重要作用[7]。渔业资源结构是在特定环境条件中长期形成的,过度捕捞是造成渔业资源衰退的最主要原因,但渔业种群变化也与栖息地环境变化和资源本身的特性有关。带鱼是我国海洋鱼类资源中资料积累最丰富、研究最系统、全面的种类,我国围绕带鱼生态习性、生活史特征、洄游分布、种群数量动态、繁殖保护等研究已有60多年的历程。根据已有文献,东海带鱼主要从栖息地区域和范围改变、种群数量年间波动、低龄阶段生长加快和性成熟提前几个方面表现出主动或被动对栖息地环境变化的适应。栖息地变动主要是带鱼对温盐和饵料生物空间变化的主动适应;种群数量年间波动则是带鱼对栖息地环境年间变化的被动适应;低龄阶段带鱼生长加快和性成熟提前等生物学特性变化则是长期的生物学适应性表现,该变化是对自身资源量减少的适应,也是对栖息地环境变化的适应。基于此,本研究从栖息地选择、种群数量年间波动、生物学变化等角度阐述东海带鱼对栖息地环境变化的适应性表现,从而为从栖息地角度解析东海带鱼种群变动原因的研究明晰方向,进而为东海带鱼资源保护和生境修复提供科学依据。
1 栖息地选择
栖息地是生物赖以生存和繁衍的空间和环境,关系着生物的食物链和能量流[10]。鱼类关键栖息地包括越冬场、产卵场、索饵场和洄游通道。带鱼的越冬场、产卵场和索饵场范围广阔,其在不同生活阶段,具有远距离洄游习性。据研究[1,7,11],东海带鱼种群的重点分布区在浙江近海,北部可达34°N周围的黄海中部,南可到达粤东近海;主要越冬场位于30°N以南的浙江中南部水深60~ 100 m海域,越冬期为1-3月;每年三四月起,随着水温升高,越冬鱼群大致以每2个月1个纬度的速度逐步北上,5-7月经鱼山渔场进入浙江中北部的舟山渔场和长江口渔场;产卵后主群继续北上,8-10月分布在南黄海南部索饵,索饵群体前锋可达35°N附近,与黄渤海群相混栖,但是20世纪80年代中期以后,随着带鱼资源的衰退,索饵场的北界明显南移,主要分布在东海北部至吕泗、大沙渔场的南部;10月份以后,随着冷空气南下,鱼群向南进行越冬洄游,并在浙江北部沿岸低盐水系和外海高盐水系交汇的混合水区短暂栖息,形成著名的冬季嵊山带鱼渔场;之后随着水温继续下降,小部分带鱼游向外海越冬场,大部分带鱼继续南下洄游到浙江中南部外侧海区的越冬场。福建和粤东近海的越冬鱼群2-3月即开始北上,3月即有少数鱼群在福建近海产卵繁殖,4-6月产卵后进入浙江南部,并随台湾暖流继续北上,秋季分散在浙江近岸索饵。
带鱼属暖水性中下层鱼类,栖息环境相对稳定,自身具有昼沉夜浮的习性。影响底层鱼类聚集的非生物因子较多,如温度、盐度、水深、水团、溶解氧、底质类型等[7,12]。由于带鱼养殖难度大[13],至今尚没有通过控制实验方法掌握环境因子对其不同发育阶段的影响,过去较多的是利用东海带鱼栖息地环境因子的实测数据分析东海带鱼栖息地空间分布及其变化。以往研究显示,东海带鱼聚集特征具有年代际变化,该变化与气候、流场、风场、温盐及生物等环境变化有关[14-17]。水温通过影响鱼类代谢强度、体温和性腺发育而直接影响其摄食、产卵和越冬区域;盐度通过影响鱼类呼吸代谢、消化系统和渗透压调节而直接影响其空间分布;同时,温盐等理化条件又通过影响天然饵料(浮游动物和小型鱼类)数量和分布而间接影响带鱼栖息地选择[16]。东海带鱼基本是沿着沿岸水系、台湾暖流和黄海冷水团的混合水域分布;鱼群聚集区一般位于台湾暖流舌锋附近;中心渔场位置与黑潮强度密切相关[9,16-20]。东海带鱼越冬洄游路线时空变化主要受台湾暖流进退和消长的影响[11]。东海带鱼分布区域很大程度上受到水温和盐度变化的影响[9,16-23]。海表温度降低,带鱼聚集区南移;海表温度升高,聚集区北移,均靠近黑潮北分支左侧水域[11]。盐度影响着带鱼的聚集和中心渔场的移动[19],长江径流量大的年份,中心渔场一般偏向东南;径流量小的年份,一般偏向西北[24]。幼带鱼数量峰值通常出现在水温22~ 26℃、盐度26~ 28条件下[25]。从带鱼自然分布的数据看,低盐高温有利于东海浮游植物的生长和带鱼的洄游、索饵、聚集并形成渔场[17-18]。陈亚瞿等利用1960-1981年东海区浮游生物的分布资料分析东海带鱼饵料基础与渔场的关系,发现浮游动物生物量及磷虾等的数量分布同东海带鱼渔场的位置变化和鱼群移动关系密切,该关系在冬、春季浙江南部近海渔场和夏、秋季长江口及舟山等渔场尤为明显[26]。
近40年来,东海鱼类栖息地受自然变化和人类活动影响而出现自然岸线阻断、海水变暖和富营养化等变化。岸线变化可能阻断和改变鱼类的洄游路线;水温变化直接影响鱼类的代谢、摄食、产卵及洄游等行为;营养盐输入和分布变化则会通过影响初级生产力和浮游生物结构而改变鱼类饵料供给;捕捞过度不仅直接破坏渔业资源量,同时也通过降低鱼群抵御环境变化的能力,从而放大栖息地环境变化对鱼类的影响[27-30]。因而,东海海域的栖息地环境变化很可能是东海带鱼资源变化的原因之一。然而,由于我国之前对历史数据的挖掘较少,利用东海实测数据,分析近40年来东海带鱼栖息地变动的研究仍比较有限,现有必要整合东海近40年的水文、化学和饵料生物实测数据,揭示东海带鱼关键栖息地内温度、盐度和饵料生物的历史变化,揭示其与东海带鱼栖息地区域和范围变动的相关性。
2 种群数量年间波动
评价鱼类种群数量年间波动最直观参数为渔获量,较客观参数为单位捕捞努力量渔获量(CPUE)[31]。东海带鱼历年渔获量统计数据显示,1951-1974年,渔获量随捕捞努力量的增长而不断提高;1974-1988年,渔获量却随着捕捞努力量的增长而下降,说明近海带鱼资源处于衰退状态;1988-2000年,随着作业渔场由沿岸和近海向外海转移,渔获量又开始大幅回升;2000年后,沿岸、近海和外海渔场的带鱼资源均出现明显衰退,渔获量开始逐步下降[9,16,32],但依然可以保持一定产量(表1),是我国单鱼种产量最高的一种经济鱼类。
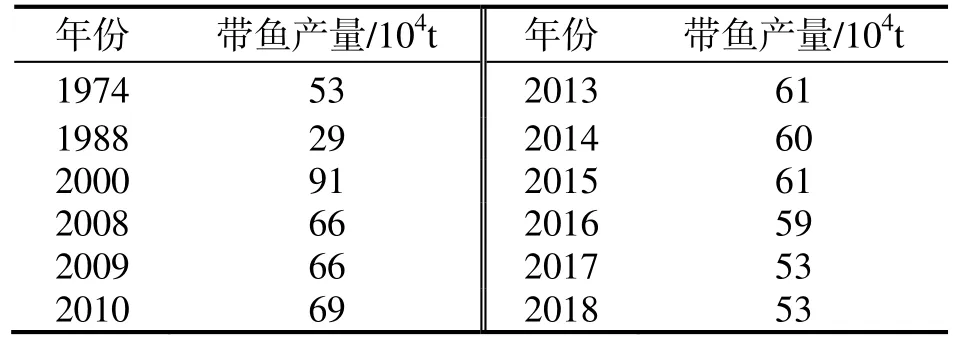
表1 东海带鱼产量统计 [3,9]Table 1 Yields of Trichiurus lepturus in the East China Sea
带鱼渔获量变化包括变化趋势和年间变动,变化趋势主要受捕捞影响,排除捕捞效应后,带鱼CPUE的年间波动与气候因素和栖息地环境因素密切相关。多数研究阐述了温盐对带鱼渔获量有重要影响[14,17,33-34]。何发祥等[35]研究浙江近海1961-1982年冬汛带鱼渔获量变化与Elnino的关系,发现了厄年和准厄年,冬汛带鱼产量高,反之则低。Elnino年台湾暖流和南黄海冷水皆强,初夏沿岸水强或者较强,使中心产卵场位置偏北偏向岸方向,中心产卵场面积大,冬汛产量则高;反之,台湾暖流或南黄海冷水弱,则中心产卵场可能偏南偏外,中心产卵场范围小,冬汛产量则低。同时,Elnino年5-6月(带鱼生殖高峰期),长江径流骤增带来大量营养盐和有机物质,使东海渔场水质肥沃、饵料生物繁多;而闽浙沿岸夏季上升流强度弱,使中上层鱼类减少,食物丰富和竞争者少均有利于带鱼渔获量增加。俞存根[36]报道了浙江近海冬汛带鱼渔获量与太阳黑子、气象要素关系,认为降水多的年份,带鱼渔获量往往偏低;降水少的年份,渔获量往往偏高。降水多的年份,一方面大陆径流增强,沿岸水系势力增大,分布偏外,水平混合区扩大,另一方面大量降水致使表层盐度偏低,上下层海水混合减弱,带鱼垂直移动受到限制,从而导致渔场分布偏外,鱼群稀疏,难以捕获大网头;反之,鱼群密集,中心渔场明显,渔获量往往较大。同时,9月份以后的气温变化也要影响带鱼渔获量,暖冬年及冷冬年往往均产况欠佳,渔获量偏低;而冬汛前期气温偏低,后期气温偏高的年份,则带鱼渔获量往往较高。王跃中等[9,16]详述了气候变动(陆地降水和径流、季风风速、海表水温和热带气旋)与移除捕捞努力量影响后的东海带鱼渔获量年间变动间的密切相关性,认为气候因素对渔获量年间变动的影响途径主要是通过改变栖息地营养盐分布和饵料生物供给,如陆地降雨和径流会影响营养盐输入量,季风会影响营养盐的扩散、循环及利用效率,水温则影响带鱼性腺发育、成熟和饵料供应。然而,利用实测数据分析和验证营养盐及饵料生物变动与东海带鱼CPUE年间变动关系的研究尚未见报道。
3 生物学变化
带鱼的生活史选择型,从单位补充量产量曲线看,为中间偏k-选择型;但从生物学特性看,世代过程较短,性成熟迅速,出生后第二年即可全部达到性成熟而自然产卵,且具有再次或多次性产卵习性,自然死亡率较高,寿命相对短,具有r-选择型特性[24]。因此,带鱼能长期承受较高强度的捕捞压力,对捕捞的适应能力较大黄鱼、小黄鱼等k-选择型鱼类高。此外,带鱼食性广,且不同年代、季节、海区的食物组成有差异[26,37-39],表明带鱼具有很强的摄饵可塑性和就地摄食特点。
带鱼“衰而不竭”的原因,除与其偏r-选择型的生活史特征和摄饵可塑性强的习性有关外,也与其生物学适应性调节有关。东海带鱼衰退的生物学特征包括小型化、低龄化、生长加快、性成熟提前、产卵时间延长、繁殖力提高而卵径变小[2,40-47]。带鱼小型化和低龄化是过度捕捞的直接结果[48],同时也与成体带鱼的重要食物数量剧烈波动有关[32,49];性成熟提前、繁殖力提高是带鱼对资源量减少而增加补充量的一种适应;而低龄阶段生长加快和性成熟提前的变化则可能主要与栖息地温度、饵料数量和质量变化有关[43,46]。
温度升高有利于促进鱼类生长和性腺发育,同时促进饵料生物生长。东海海域自1976年开始,海表温度升高明显,冬季平均升温1.0~ 1.5℃,夏季平均升温0.5 ℃[50]。研究发现21世纪初的1龄以内东海带鱼生长速度较20世纪几个年代均高[46],春夏季同样肛长的带鱼,从20世纪50、60年代到90年代有逐渐增重的趋势;带鱼亲体初次性成熟年龄提前,使带鱼产卵亲体数量增加和产卵时间延长,以至于东海带鱼在近海、外海几乎全年都在产卵[43-45]。分析认为,海水温度持续上升是东海带鱼生长加快和性成熟提前的原因之一[44-45]。徐汉祥等还发现东海带鱼补充群体数量也与水温正相关[45]。
饵料生物充足可能也是带鱼生长加快和性成熟提前的一个重要原因。带鱼的摄食强度较高,生长对饵料生物依赖性强[26,39,51]。同时,东海带鱼是一种主食磷虾等甲壳动物和小型鱼类,兼食头足类和毛颚动物等的广食性凶猛鱼类,摄饵可塑性和食物竞争能力强。带鱼在生长发育过程中,食性由浮游动物食性逐步转变为游泳动物食性,体长200 mm以下带鱼主要以磷虾、糠虾为食[1,26,39,51];且随着东海渔业资源的整体衰退,鱼类的营养级普遍降低[52],加之带鱼个体小型化,磷虾类和浮游幼虫等饵料生物在东海带鱼食物中的相对重要性(%IRI)在提高[39]。因此,索饵季节,低年龄阶段带鱼的生长状况可能与磷虾等浮游动物数量密切相关。自20世纪80年代至21世纪初,东海海域溶解无机氮(DIN)入海总量由8.0×105t/a上升至26×105t/a;磷酸盐(PO4-P)入海总量由1.7×104t/a上升到7.0×104t/a[53]。随着营养物质输入的增加和水温的升高,浮游植物数量增加明显,并经常形成藻华[54-55],大量藻类的沉降为底层浮游动物和鱼类的生长提供了丰富的饵料基础,从而可能有利于带鱼幼体生长和发育加快[43,46]。空间分布上,幼带鱼主要分布在浙江北部15 m等深线、南部10 m等深线以东的东海海域,北部多于南部;不同水深均有分布,中上层较多;表层个体较大,而底层个体较小[25]。50年来,东海渔业资源量减少、群落结构发生变化,中、上层鱼类、虾、蟹和水母类比例升高,底层鱼类比例下降[27,56-58],可见,带鱼的食物和空间竞争对象在减少。此外,与小型鱼类相比,磷虾和浮游幼虫生长快速,快速生长的生物通常具有较高比例的磷含量(C∶P 和 N∶P比较低),以满足核糖核酸转录和蛋白质合成需要[59],而带鱼摄入较高磷含量的食物,可能也是带鱼生长和发育加快的原因之一。虽然已有研究表明浮游动物(尤其是磷虾、糠虾等甲壳类浮游动物)作为带鱼幼鱼的饵料生物,可能对低龄阶段带鱼生长加快和性成熟提前至关重要,但是目前还未见带鱼索饵场饵料浮游动物历史变化与带鱼生物学变化关系的研究报道。因此,分析东海饵料浮游动物数量的空间分布和历史变化,验证饵料生物充足与带鱼生长加快和性成熟提前之间的关系,有助于更全面地从栖息地适应角度解析东海带鱼种群生物学变化的机制。
4 结语
近些年来,我国陆续建立东海产卵带鱼保护区和东海带鱼国家级水产种质资源保护区,并实施伏季休渔制度,以加强对带鱼种群的养护与管理,减缓其资源衰退。东海带鱼的衰退不仅与捕捞活动有关,也与东海海域的气候变化、物理环境和饵料生物演变有关,而东海带鱼能够衰而不竭除了与带鱼偏r-选择型的生活史特征、摄饵可塑性强的习性有关外,也与其较强的栖息地适应性有关。本综述显示,东海带鱼较强的栖息地适应性主要表现在:(1)对温盐响应敏感,且可远距离洄游,因此可及时调整其栖息地区域和范围,选择温盐适宜的栖息环境,以提高种群存活率;(2)对饵料变化的响应敏感,浮游生物的空间和时间变化会影响带鱼的栖息地选择和种群数量变化;(3)通过改变生物学特征适应环境温度、饵料数量和质量的变化,如加快生长速度和性成熟提前等。解析栖息地环境演变与带鱼种群变动间的相互作用机理对理解和预测东海带鱼资源量变动具有重要意义。根据目前的研究情况,笔者认为现阶段应该重点从栖息地海流(温度、盐度)和关键饵料生物历史变化等角度揭示东海带鱼的适应性特征,为揭示带鱼等东海渔业资源衰退原因和生境修复提供可靠的科学依据。
