不同地形樟子松天然林土壤呼吸特征及其影响因素
2020-01-15李岳泰满秀玲喻武陈婧王氏明演
李岳泰,满秀玲,喻武,陈婧,王氏明演
(1.西藏农牧学院, 西藏 林芝 860000;2.东北林业大学 林学院, 哈尔滨 150040)
0 引言
土壤呼吸是指未扰动的土壤向外界释放CO2的总和[1-2],通常包含土壤根呼吸(即自养呼吸)、土壤微生物呼吸(即异养呼吸)、含碳物质的化学氧化和土壤动物呼吸,后两者的作用比较弱,通常可以忽略不计[3-4]。据估计,每年通过土壤呼吸向外界排放的CO2约占全球CO2交换量的25%,是化石燃料的10倍以上,土壤呼吸已成为全球碳循环中最大通量之一[5],因此土壤呼吸的研究对目前日益严重的温室效应问题具有重要意义。森林作为陆地生态系统的主体,在全球的土壤呼吸通量中占据重要地位[6]。近年来,大量学者对不同森林类型的土壤呼吸进行研究,主要集中在土壤呼吸速率在不同时间尺度上的动态及其与环境因子的关系、全年呼吸量的估算等[7]。土壤呼吸受到土壤温度、土壤湿度、土壤养分、pH、土壤孔隙度、光照强度、地形、植被类型和人类活动等因素的影响[8-9]。Raich等[10]研究认为,温度和降水对土壤呼吸的时空变化产生了重要影响。相关研究表明[11-12],地形对土壤养分的积累及土壤与环境的物质能量交换密切相关,在不同坡位土壤的理化性质、水土流失状况也存在差异,进而对土壤呼吸产生影响。目前关于地形条件对森林土壤呼吸的影响研究较少,且结论不一,如卿明亮等[13]对不同坡位的油松林土壤呼吸研究发现,坡中的土壤呼吸年积累量低于坡上和坡下,坡位间存在显著差异;郑威等[14]对坡地红锥人工林土壤呼吸的研究表明,土壤呼吸有随坡位升高而增大的趋势。
大兴安岭林区位于我国最北端,是我国最大的寒温带针叶林区,也是我国天然樟子松林主要分布区。本研究以不同地形天然樟子松林(平地和坡地)为对象,对土壤总呼吸速率和各组分呼吸速率(自养呼吸、异养呼吸、枯落物呼吸和矿质土壤呼吸)进行研究,探讨樟子松林土壤呼吸特征及影响因素,为我国寒温带地区森林生态系统碳循环研究提供帮助,对研究寒温带森林碳释放有着重要意义。
1 研究区概况
研究地点位于黑龙江漠河森林生态系统国家定位观测研究站N 53°17′~53°30′、E 122°06′~122°27′,该区域为寒温带大陆性季风气候,气候特点夏季湿润多雨,冬季寒冷漫长,年平均气温-4.9 ℃,年平均无霜期80~90 d,年平均降水量350~500 mm,且集中在7—8月。土壤类型主要为棕色针叶林土,此外还有泥炭土和草甸土等非地带性土壤,是我国多年冻土主要分布区。森林植被主要有兴安落叶松(Larixgmelinii)林、樟子松(Pinussylvestrisvar.mongolica)林、山杨(Populusdavidiana)林和白桦(Betulaplatyphylla)林等类型。
2 研究方法
2.1 样地设置
由于天然樟子松林主要分布在大兴安岭地区的阳坡,平地也有部分分布,因此在前期踏查的基础上,于2018年5月初,选择平地和坡地樟子松林进行土壤特征研究。坡地樟子松林坡度为30°~40°,坡长为400 m左右,又分为坡下、坡中和坡上3个坡位,分别设置3块20 m×30 m的调查样地,在样地内随机布设3组内径10.4 cm、高度10 cm的PVC土壤呼吸环,每组为两个环,并去除其中一个环中的枯枝落叶层,以测定土壤总呼吸(RS)和去除枯落物的土壤呼吸(R1);另外在每个样地周围设置3个1 m×1 m的小样方,进行挖壕断根处理。具体做法如下:在每个小样方四周挖壕至植物根系分布层以下,以确保切断样方与外界的根系连接(本地区土层较薄,挖至30 cm深度即可),壕沟内用双层塑料薄膜进行隔离,并将土回填于薄膜内,定期清除样方内杂草,在每个小样方内设置3个呼吸环,以测定土壤异养呼吸(RH)。为排除根系的呼吸作用,根据以往相关研究[15-17],待挖壕完成1个月之后再进行异养呼吸的测定。观测期为2018年6月至10月,于每月中旬用LI-6400光合仪连接土壤呼吸室对样地内所有呼吸环进行测定。
2.2 呼吸组分的计算
土壤总呼吸(RS)、去除枯落物的土壤呼吸(R1)、土壤异养呼吸(RH)由仪器直接测得;土壤自养呼吸(RA)等于土壤总呼吸(RS)减土壤异养呼吸(RH);枯落物呼吸(RL)等于土壤总呼吸(RS)减去除枯落物的土壤呼吸(R1);矿质土壤呼吸(RM)等于土壤异养呼吸(RH)减枯落物呼吸(RL)。
2.3 影响因子的测定
樟子松林土壤10 cm温湿度测定:土壤温度(T10)的测定用LI-6400便携式CO2/H2O分析系统自带的土壤温度探针,土壤湿度(W10)的测定用TDR 100。
其他因子测定:每次测定土壤呼吸时,于各样地随机选取3个点取0~10 cm层的土壤样品,同一样地的土壤样品混合后,带回实验室风干、过筛备用。土壤总有机碳采用重铬酸钾-外加热法;土壤全氮采用半微量凯氏定氮法;土壤容重采用环刀法;土壤pH用pH计法。
2.4 土壤呼吸与土壤温、湿度的模型
土壤呼吸速率和土壤温度之间的关系采用指数模型[18]:
RS=αeβT。
式中:RS为土壤呼吸速率,μmol /(m2·s);α为土壤温度为0 ℃时的土壤呼吸速率,μmol /(m2·s);T为土壤温度,℃;β为指数模型中的温度反应系数。
土壤呼吸对温度的敏感性用Q10值表示,即温度升高10 ℃时土壤呼吸速率的变化倍数,计算式为:
Q10=e10β。
土壤呼吸速率和土壤湿度之间的关系采用线性模型[19]:
RS=aW+b。
式中:W为土壤湿度,%;a为水分反应系数;b为截距。
2.5 数据处理
用Microsoft Excel 2016进行数据整理及作图,用SPSS 24对不同立地条件土壤呼吸速率进行描述性统计分析和单因素方差分析,对土壤总呼吸和土壤温、湿度进行曲线回归分析,对土壤总呼吸与土壤总有机碳、全氮、碳氮比、pH进行pearson相关分析。
3 结果与分析
3.1 不同地形樟子松林土壤总呼吸变化特征
各地形樟子松林土壤总呼吸速率(RS)月动态均表现为单峰曲线(图1),平地、坡下和坡中的峰值出现在7月,坡上峰值在8月。平地RS的波动幅度最大(1.00~8.78 μmol/(m2·s)),坡中波动最小(1.32~6.19 μmol/(m2·s))。6—7月樟子松林土壤总呼吸大幅升高,且平地土壤总呼吸显著高于坡地(P<0.05),坡地各坡位间差异不显著(P>0.05);6月平地樟子松林RS最高(6.24 μmol/(m2·s)),分别比坡下、坡中、坡上高43.12%、72.38%、37.44%;7月平地樟子松林RS最高(8.78 μmol/(m2·s)),分别比坡下、坡中、坡上高30.27%、41.84%、30.85%,同时平地、坡下、坡中在7月达到峰值,分别为8.78、6.74、6.19 μmol/(m2·s)。8月平地、坡下、坡中RS开始出现缓慢下降,坡上则是缓慢上升;8月坡上达到峰值(7.23 μmol/(m2·s)),平地显著高于坡地(P<0.05)。9月均开始出现大幅下降,平地降速最快,下降了54.61%,坡下、坡中、坡上分别下降了34.67%、29.06%、19.01%。9月平地、坡下、坡中RS已非常接近,且显著低于坡上(P<0.05)。10月各地持续大幅下降,平地、坡下、坡中、坡上降幅分别为74.07%、69.75%、69.12%、67.05%,10月RS均达到最低值,分别为1.00、1.33、1.32、1.93 μmol/(m2·s),且无显著差异(P>0.05)。
由此可见,樟子松林RS的月动态呈单峰曲线,平地、坡下和坡中的峰值出现在7月,坡上峰值在8月。观测期内RS均值由大到小顺序为:平地、坡上、坡下、坡中,平地显著高于坡下和坡中(P<0.05),与坡上差异不显著(P>0.05)。平地RS在6—8月份均显著高于坡地(P<0.05),9月和10月各地形差异不显著(P>0.05)。6—8月本地区的降雨较为集中,且土壤温度较高,良好的外界条件提高了土壤中微生物的活性,同时植被根系生长迅速,根呼吸活跃,使得土壤呼吸速率较高[17]。坡地RS的变化较为一致,除7月以外,在6、8—10月由大到小均表现为:坡上、坡下、坡中。

图1 不同地形土壤总呼吸速率月变化Fig.1 Monthly variation of total respiratory rate of soil under different terrains
3.2 不同地形樟子松林土壤异养呼吸和自养呼吸变化特征
异养呼吸(RH)和自养呼吸(RA)在各地形均表现出相似的规律性(图2),RH的峰值均出现在7月,RA的峰值均在8月,月动态均呈单峰曲线。樟子松林RH在平地、坡下、坡中、坡上的波动范围分别为0.49~7.86、0.98~4.63、0.74~4.54、1.00~5.87 μmol/(m2·s),可见其在平地的波动要明显大于坡地各坡位。6—7月樟子松林RH处于快速上升期,平地RH显著高于坡地(P<0.01),7月坡上的上升速度远快于坡中和坡下,且7月均达到峰值(7.86、4.63、4.54、5.87 μmol/(m2·s)),坡上峰值显著高于坡下和坡中(P<0.05),平地显著高于坡地(P<0.01)。8月樟子松林RH开始出现下降,平地、坡下、坡中、坡上的下降幅度分别为16.15%、23.67%、9.22%、23.91%,8月平地RH显著高于坡地(P<0.01),坡地各坡位间无显著差异(P>0.05)。9月平地出现大幅下降,下降幅度为58.39%,坡地降幅较小;9月RH均降至较低水平,且无显著差异(P>0.05)。10月平地、坡下、坡中、坡上均大幅下降至最低值,降幅分别为82.02%、71.05%、72.37%、69.56%。
可见,不同地形下樟子松林RH的波动范围在0.49~7.86 μmol/(m2·s)之间,6—10月呈单峰曲线变化趋势,峰值均出现在7月。平地和坡地RH在6—8月份差异较大,这段时期平地RH显著高于坡地(P<0.01),9—10月份平地和坡地无显著差异。观测期内各立地条件樟子松林RH均值由大到小顺序为:平地、坡上、坡下、坡中,平地分别比坡下、坡中、坡上高50.80%、54.28%、30.64%,平地RH显著高于坡下和坡中(P<0.05)。
樟子松林坡上和坡下的RA波动范围大于平地和坡中(图2)。6月平地、坡下、坡中、坡上的RA分别为0.44、1.23、0.52、1.10 μmol/(m2·s),坡下和坡上RA显著高于平地和坡中(P<0.05),6—8月平地、坡下、坡中逐渐上升,上升幅度分别为425.00%、260.98%、353.85%;坡上呈波动上升,上升幅度为251.01%,可见平地上升速度快于坡地;8月各地形均达到峰值,平地和坡中的峰值比较接近,分别为1.87、1.84 μmol/(m2·s),坡下、坡上峰值较高,分别为3.21、2.76 μmol/(m2·s)。9月平地和坡下出现大幅下降,降幅为41.28%和66.28%,坡中、坡上降幅较小,坡下的下降速度要远快于其他立地条件;9月坡上RA显著高于其他地形(P<0.05),平地、坡下、坡中较为接近。10月持续大幅下降,均降至较低水平,坡下、坡上出现最小值,分别为0.35、0.93 μmol/(m2·s)。

图2 不同地形异养呼吸和自养呼吸速率的月变化Fig.2 Monthly variation of heterotrophic respiration rate and autotrophic respiration rate under different terrains
由此可见,不同地形樟子松林RA的波动范围在0.35~3.21 μmol/(m2·s)之间,峰值出现在8月,最低值出现在6月或10月,变化趋势表现为6—8月波动上升,8—10月逐渐下降。6—8月坡下RA最高,9—10月坡上RA最高,平地在整个观测期内均处于较低水平。观测期内各地形RA均值由大到小顺序为:坡上、坡下、坡中、平地,平地RA均值显著低于坡下和坡上(P<0.05)。
3.3 不同地形樟子松林枯落物呼吸和矿质土壤呼吸变化特征
不同地形樟子松林土壤枯落物呼吸速率(RL)呈单峰曲线变化趋势,坡下和坡上峰值出现在8月份,平地和坡中则出现在9月份(图3)。在观测期内樟子松林RL波动范围在0.19~2.22 μmol/(m2·s)之间。6月平地和坡下枯落物呼吸(RL)显著高于坡上(P<0.05);7—8月呈上升趋势,8月坡下、坡上RL均达到峰值,分别为1.81、2.22 μmol/(m2·s),坡中RL显著低于平地、坡下、坡上(P<0.01)。9月各地形变化趋势有所不同,平地和坡中持续上升至峰值,分别是1.83、1.60 μmol/(m2·s),上升了4.38%、72.40%,坡下和坡上开始下降,分别降低了36.58%、49.17%;9月平地和坡中RL显著高于坡下和坡上(P<0.05)。10月均出现大幅下降,平地、坡下、坡中、坡上分别降低了81.20%、83.48%、72.14%、59.17%;10月除坡上外,平地、坡中、坡下均降至最小值,分别为0.34、0.19、0.45 μmol/(m2·s),坡上最小值在6月,大小为0.44 μmol/(m2·s)。
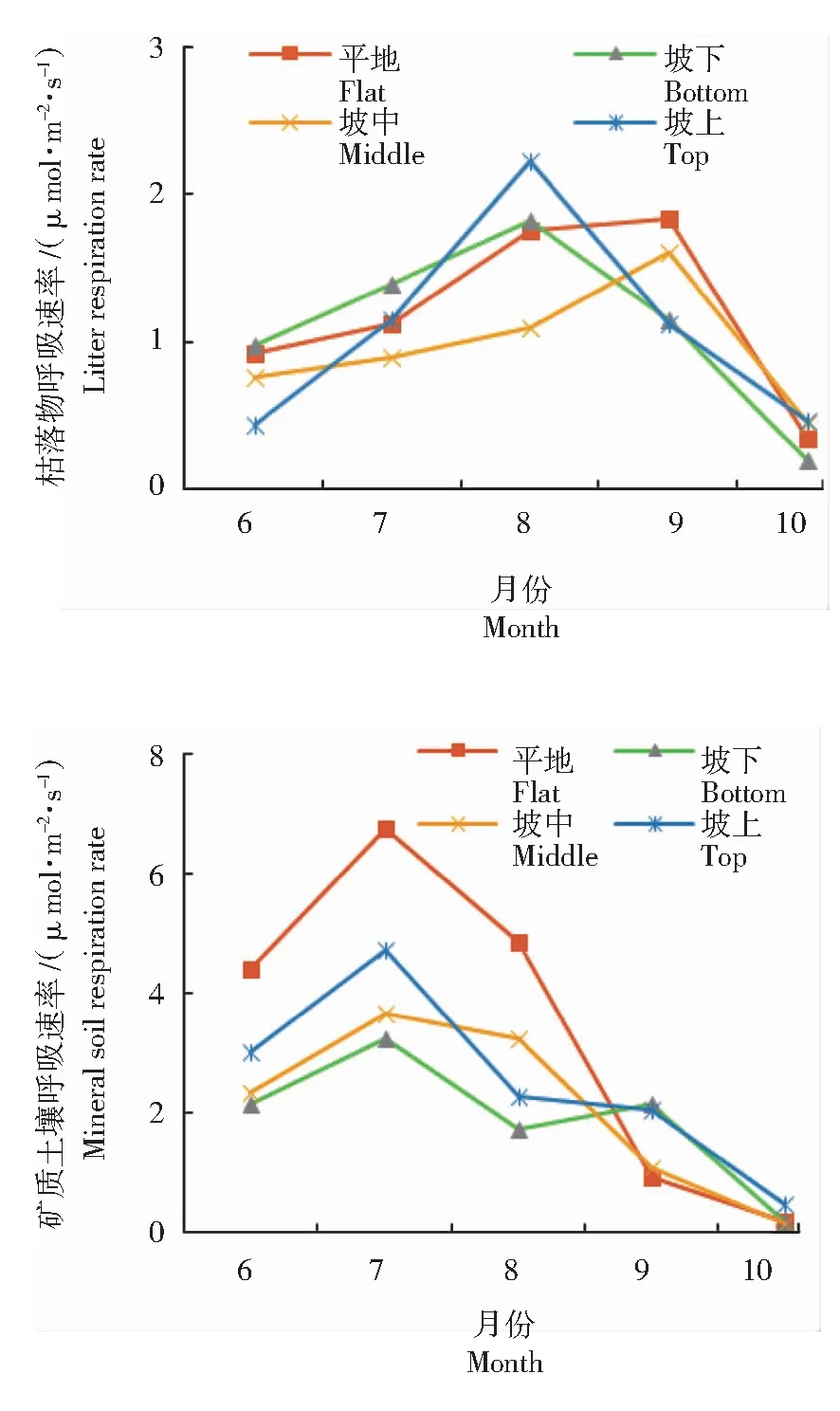
图3 不同地形枯落物呼吸和矿质土壤呼吸速率的月变化
综上可知,樟子松林土壤枯落物呼吸速率(RL)在生长季呈单峰曲线变化,坡下和坡上峰值在8月,平地和坡中峰值在9月。RL变化趋势为坡下和坡上在6—8月逐渐上升,8—10月逐渐下降;平地和坡中在6—9月逐渐上升,9—10月逐渐下降。除10月无显著差异外,6—9月樟子松林RL在不同立地条件下均存在显著差异(P<0.05),且在8月坡中RL与平地、坡下、坡上差异达极显著水平(P<0.01)。
矿质土壤呼吸(RM)的变化趋势与总呼吸和异养呼吸相似(图3),各地形的最大值均出现在7月,最小值均在10月。樟子松林RM总体波动范围在0.13~6.74 μmol/(m2·s)之间,平地的波动范围最大,坡地各坡位波动范围较小且比较接近。6月平地显著高于坡地(P<0.05),坡地不同坡位间无显著差异(P>0.05);7月均快速上升至峰值,平地、坡下、坡中、坡上峰值分别为6.74、3.25、3.24、4.73 μmol/(m2·s),平地RM显著高于坡地各坡位(P<0.01),坡上显著高于坡中和坡下(P<0.05)。8月均开始出现大幅下降,平地、坡下、坡中、坡上分别降低了28.10%、46.97%、17.24%、52.40%;8月平地RM显著高于坡地(P<0.01),坡中显著高于坡下(P<0.01)。9—10月持续下降,均降至10月的最小值,分别为0.16、0.16、0.13、0.47 μmol/(m2·s),且无显著差异(P>0.05)。
综上可见,矿质土壤呼吸(RM)的波动范围在0.13~6.74 μmol/(m2·s)之间,峰值出现在7月,变化趋势为6—7月均表现为逐渐上升,7—8月均逐渐下降,8—10月为波动下降,最小值均出现在10月。6—8月平地RM显著高于坡地(P<0.01),9月坡下和坡上RM显著高于平地和坡中(P<0.01),10月平地和坡地无显著差异(P>0.05)。观测期内RM的均值由大到小顺序为:平地、坡上、坡中、坡下,平地显著高于坡下和坡中(P<0.05)。
3.4 土壤呼吸影响因素分析
3.4.1 土壤总呼吸速率(RS)与土壤10 cm温度(T10)的相关性
樟子松林土壤10 cm温度(T10)的月动态呈单峰曲线(图4),峰值出现在8月,各地形的T10无显著差异(P>0.05)。土壤总呼吸速率与土壤温度的回归分析结果表明(表1),樟子松林土壤总呼吸速率(RS)与土壤10 cm温度(T10)存在极显著的指数相关(P<0.01),且坡地的相关性要强于平地。T10解释了樟子松林土壤总呼吸季节变化的69.3%~83.8%。温度敏感性系数Q10表示温度每升高10 ℃土壤呼吸所增大的倍数,各立地条件Q10由大到小顺序为:坡下、坡上、坡中、平地,表明坡地对土壤温度变化的敏感程度要大于平地。

图4 不同地形10 cm土壤温度和土壤湿度的月变化Fig.4 Monthly variation of soil temperature and soil moisture of 10 cm under different terrains

表1 不同地形土壤总呼吸速率与 土壤10 cm温度(T10)的回归关系
3.4.2 土壤总呼吸速率与土壤10 cm湿度(W10)的相关性
樟子松林土壤10 cm湿度(W10)的变化趋势呈单峰曲线(图4),峰值在8月。观测期内坡上W10显著低于其他地形(P<0.05),平地在8—9月显著高于坡地(P<0.01)。土壤总呼吸速率与土壤10 cm湿度回归分析结果表明(表2),在坡中和坡上,樟子松林土壤总呼吸与土壤10 cm湿度(W10)存在显著相关性(P<0.05),而平地和坡下的相关性不显著(P>0.05),且R2有随坡位升高而增大的趋势,表明随着坡位的上升,土壤总呼吸与W10的相关性越来越强。W10解释了樟子松林土壤总呼吸季节变化的6.4%~41.5%,可见土壤总呼吸与土壤湿度的相关性较土壤温度弱。

表2 不同立地条件土壤总呼吸速率与土壤10 cm 湿度(W10)的回归关系Tab.2 The correlation between total soil respiration and soil moisture (W10) of 10 cm under different terrains
3.4.3 土壤总呼吸速率与土壤养分的相关性
将土壤总呼吸速率与0~10 cm层的土壤总有机碳、全氮、碳氮比(C/N)和pH进行pearson相关分析,结果显示不同地形樟子松林土壤总呼吸速率与土壤养分的关系有所差异(表3)。平地、坡地土壤总有机碳和土壤总呼吸速率均呈极显著正相关(P<0.01),且随着坡位的升高,坡地樟子松林土壤呼吸速率与总有机碳相关性有变强的趋势;全氮与土壤总呼吸速率呈正相关关系,且在坡中与土壤总呼吸速率的相关性达极显著(P<0.01);碳氮比在平地和坡上与土壤总呼吸速率呈极显著正相关(P<0.01),其余地形相关性不显著;pH在坡上与土壤总呼吸速率呈极显著正相关(P<0.01)。

表3 不同地形土壤总呼吸速率与土壤养分、pH的相关性Tab.3 The correlation between soil total respiration and soil nutrient and pH under different terrains
注:*.在0.05级别(双尾)相关性显著;**.在0.01级别(双尾)相关性显著。
Note: *.Correlation is significant at the 0.05 level (two-tailed); **. Correlation is significant at the 0.01 level (two-tailed).
4 讨论
4.1 不同地形土壤呼吸组分的差异
由分析可知,樟子松天然林土壤总呼吸月动态规律呈单峰曲线,峰值出现在7—8月份,符合以往相关研究[18-20]。观测期内土壤总呼吸速率的均值变化范围为4.28~5.66 μmol/(m2·s),与段北星等[21]研究兴安落叶松林土壤总呼吸速率范围4.71~7.41 μmol/(m2·s)较为接近。土壤总呼吸在平地的均值显著高于坡下和坡中,与坡上差异不显著(P>0.05)。郑威等[14]研究坡地红锥人工林土壤呼吸发现,土壤总呼吸随着坡位的升高而上升,与本研究结果存在差异,樟子松林坡下的土壤呼吸要高于坡中,这可能是地区和林型的不同而造成的。大兴安岭地区土层较薄,不同坡位间土层厚度差异比较明显,平地和坡下土层相对较厚,土壤养分含量更多,植被生长更为茂盛,有效地促进了土壤呼吸。观测期内异养呼吸、矿质土壤呼吸在平地显著高于坡下和坡中(P<0.05),异养呼吸平地均值分别比坡下、坡中高50.80%、54.28%,矿质土壤呼吸平地分别比坡下、坡中高出81.38%、63.16%。可见平地的土壤总呼吸高于坡地,主要是由于平地拥有较高的异养呼吸和矿质土壤呼吸。本研究中不同立地条件土壤总呼吸、异养呼吸和矿质土壤呼吸的月变化趋势较为相似,峰值均集中在7月或8月,最小值在10月,且在6—8月平地RS、RH、RM均显著高于坡地(P<0.01),在9—10月平地和坡地的差异不显著(P>0.05),呼吸速率表现出明显的季节变化,符合相关研究[22-24]。土壤呼吸具有明显季节变化的原因主要是:夏季高温多雨,植被根系生长旺盛,微生物代谢活动较强,此外枯落物在夏季湿热条件下分解更快,可为土壤提供大量养分[25];冬季植被进入休眠期,环境条件恶劣,植物及微生物活动性减弱。大兴安岭地区纬度较高,9—10月份已步入冬季,温度迅速下降,各土壤呼吸组分均降至较低水平,在不同立地间无显著差异。
自养呼吸在平地的均值显著低于坡下和坡上(P<0.05)。从时间动态上看,坡上自养呼吸在9—10月份明显高于其他立地条件。可能是9—10月土壤温度迅速下降,樟子松林样地位于阳坡,坡上由于地理条件的优势,光照更加充足,9—10月土壤温度高于其他立地条件(图4),有效地促进了土壤呼吸。郑威等[14]研究显示,在温度较低的冬季,坡上红锥人工林土壤呼吸要高于其他坡位,主要原因也可能是太阳辐射。枯落物呼吸在整个观测期内均处于较低水平,各地形差异不显著(P>0.05)。
4.2 影响因子分析
土壤温度通常与土壤呼吸的关系极为密切,两者均表现出明显的季节变化[26]。土壤湿度影响着植被及土壤微生物的生理活动、土壤的通透性,是土壤呼吸的一个重要影响因子[27]。在本研究中,不同地形樟子松林土壤总呼吸与土壤10 cm温度(T10)均存在极显著的指数相关(P<0.01),T10可以解释不同地形下樟子松林土壤总呼吸月变化的69.3%~83.8%,说明土壤温度是影响本地区樟子松林土壤呼吸的主要因子之一。这与很多人的研究结果一致[21,28]。Q10的模拟结果显示平地的温度敏感性指数低于坡地,不同坡位的Q10差异较小。相关研究表明在湿润多雨的年份Q10较低,干旱年份Q10较高,说明土壤湿度的升高会降低Q10[29]。平地可能由于土壤湿度高于坡地,因此Q10相对较低,符合上述研究。本研究中坡中和坡上的土壤总呼吸与土壤10 cm湿度(W10)存在显著相关性(P<0.05),且R2有随坡位升高而增大的趋势,表明了随着坡位的上升,樟子松林土壤总呼吸与W10的相关性变强。W10解释了樟子松林土壤总呼吸季节变化的6.4%~41.5%,可见土壤总呼吸与土壤湿度的相关性比土壤温度弱。相关研究也表明,大兴安岭地区土壤湿度对土壤呼吸的影响小于土壤温度[21,29],原因可能是本研究区域蒸散量大,土壤持水能力较低,而排水能力较强等特点也可能降低了土壤湿度对土壤呼吸的影响[30]。对土壤养分的研究显示,不同地形樟子松林土壤总呼吸与0~10 cm土壤总有机碳均呈极显著正相关(P<0.01),全氮、碳氮比、pH与土壤总呼吸呈正相关,且在部分地形相关性达极显著水平(P<0.01)。宋启亮等[31]对大兴安岭地区5种林型土壤呼吸的研究显示,土壤呼吸速率与土壤有机碳呈显著正相关,而与全氮相关性均不显著。大量研究[32-34]也显示土壤呼吸与总有机碳相关性显著。土壤有机质是微生物代谢活动的重要物质基础,是影响土壤呼吸的一个重要因素。本研究中平地、坡上土壤呼吸与总有机碳相关性较强,且随着坡位的升高,坡地樟子松林土壤呼吸与总有机碳相关性有变强的趋势。
