基于DNA条形码技术的浮游动物休眠卵种类鉴定:以洞庭湖流域常德柳叶湖为例*
2020-01-09于文波梁迪文杨宇峰崔宗斌
于文波,王 庆**,魏 南,梁迪文,杨宇峰,崔宗斌
(1:暨南大学生态学系,广州 510632) (2:中国科学院水生生物研究所,武汉 430072)
目前鉴定浮游动物休眠卵主要有传统形态学鉴定[9]和休眠卵萌发鉴定[10]两种方法. 由于浮游动物休眠卵分类学资料的匮乏,以及休眠卵体表形态的多样性,使得通过形态学鉴定浮游动物休眠卵存在一定难度而且准确率低,大多只能鉴定至属或科. 休眠卵的生理结构非常复杂,孵化成功与否取决于滞育终止的程度、卵的能量含量、非存活胚胎的数量以及环境因素,需要探索培养条件,萌发周期较长,成功率低,鉴定结果也因分类学知识的储备差别巨大. 因此,通过形态学鉴定浮游动物休眠卵存在一定难度而且准确率低. 随着DNA条形码发展为一种可靠的物种快速鉴定工具,许多学者将其应用至浮游动物种类鉴定中.
DNA条形码技术是使用一段标准化的DNA片段进行物种鉴定的方式,形似商品条形码,建立DNA序列信息与物种信息逐一对应的关系[11]. 针对不同类群的生物,通常采用不同基因作为条形码序列,例如生命条形码联盟(CBOL)植物工作组推荐叶绿体基因组的rbcL和matK片段作为植物鉴定的DNA条形码[12],真菌工作组推荐核糖体DNA内转录间隔区基因(ITS)为真菌首选DNA条形码[13],而动物物种的鉴定则采用线粒体细胞色素c氧化酶第一亚基编码基因 (cytochromecoxidase I gene, COⅠ)[14]. COⅠ基因不仅广泛应用于鸟类、蝴蝶、鱼类等物种鉴定中,而且由于使用DNA条形码鉴定时不受生物生活史阶段的限制,应用至微型浮游动物群落物种组成分析中发挥了巨大的作用,如王仁诚等对胶州湾夏季浮游动物物种组成的研究中,使用COⅠ基因成功鉴定出日本蟳、太平洋牡蛎等无脊椎动物的浮游幼体[15];张翔等使用COⅠ基因鉴定了太湖流域部分底栖无脊椎动物种类[16];Machida等使用DNA条形码技术,成功鉴定了密克罗尼西亚群岛的多种浮游生物[17];Bucklin等利用该方法,研究了北冰洋浮游桡足类种类组成[18];Baek等使用线粒体细胞色素COⅠ基因对133种桡足类个体成功进行了鉴定[19]. 本研究采用DNA条形码技术基于COⅠ基因序列对柳叶湖表层沉积物中浮游动物休眠卵进行了分子鉴定,同时对其形态信息进行采集,对建立浮游动物休眠卵图鉴,快速鉴定休眠卵种类有积极意义.
1 材料与方法
1.1 研究区域
柳叶湖(29°4′~29°8′N,111°40′~111°50′E)位于湖南省常德市境内,属于洞庭湖流域. 水域面积21.8 km2,容积为4.9×108m3,主要接纳周边山地、平原区地表径流和沟港来水,与多条河汇合后入沅江,再汇入洞庭湖,是一个典型的城市浅水湖泊.
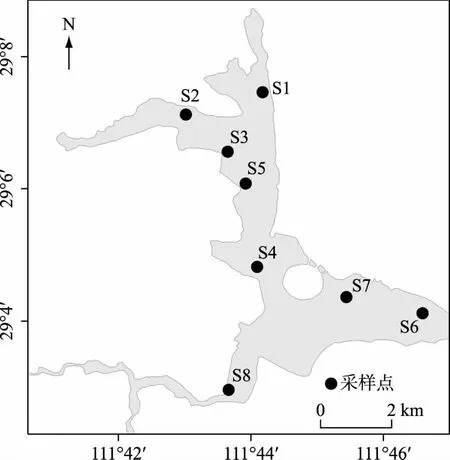
图1 洞庭湖流域常德柳叶湖表层沉积物采样点Fig.1 Sampling locations in Lake Liuye (Changde), Lake Dongting Basin, China
1.2 样品采集与处理
根据湖泊形状特征,共设8个采样点(S1~S8,图1),2018年6月使用采泥器(12 cm × 24 cm)采集柳叶湖表层5 cm沉积物,每个样点采集2个样品,共采集16个样品. 样品采集后置于4℃冰箱保存,再带回实验室后使用蔗糖浮选法[20]对休眠卵进行分离. 具体步骤如下:取20 cm3沉积物样品先后通过830 μm、64 μm筛网过滤初筛. 完成初筛后,沥净筛网中的水,用饱和蔗糖溶液(W蔗糖∶W水=1∶1)将滤渣转移到50 mL离心管中,上下颠倒使其混匀,在3000 r/min下离心20 min. 离心后将上清液再次转移至50 mL离心管,重复上述步骤至上清液无杂质,最后将上清液转移至30 μm筛网,用超纯水冲洗后转移至15 mL离心管中,4℃保存.
1.3 休眠卵的图像采集
将保存休眠卵的离心管轻轻上下颠倒,充分混匀后用吸管转移至细胞培养板中,置于体视显微镜下观察,选择卵体完整的休眠卵使用单通道移液器吸取到载玻片上,在光学显微镜(OLYMPUS,BX51)下对共计191个休眠卵进行图像采集,测量长短径,观察记录形态特征,并对图片进行统一编号,随后将单卵转移到PCR管中,待提取DNA.
1.4 DNA提取、PCR扩增及测序
本研究采用“HotSHOT”法[21]对191个休眠卵进行单卵DNA提取. 首先向含有浮游动物休眠卵的PCR管中加入10 μL裂解液,在体式显微镜下用显微镊将卵体破碎,放置于干式恒温仪95℃裂解30 min后取出,然后置于-20℃冷却5 min,最后加入10 μL中和液并吸打混匀,即完成DNA提取.
使用引物ZplankF1_t1(TGTAAAACGACGGCCAGTTCTASWAATCATAARGATATT)和ZplankR1_t1(C ̄A ̄G ̄G ̄A ̄A ̄A ̄C ̄A ̄G ̄C ̄T ̄A ̄T ̄G ̄A ̄C ̄T ̄T ̄C ̄A ̄G ̄G ̄R ̄T ̄G ̄R ̄C ̄C ̄R ̄A ̄A ̄R ̄A ̄A ̄T ̄C ̄A)对总基因组DNA的COⅠ基因进行PCR扩增[16],扩增反应体系为25 μL,包含2×Gflex PCR Buffer (Mg2+, dNTP plus) 12.5 μL,引物各0.3 μmol/L,模板为5 μL,Tks Gflex DNA Polymerase 0.5 μL,ddH2O 补足至25 μL. 扩增程序为95℃预变性1 min,94℃变性40 s,45℃退火40 s,72℃延伸45 s,5个循环;94℃变性40 s,51℃退火40 s,35个循环,最后72℃ 5 min充分延伸. PCR完成后,扩增产物经琼脂糖凝胶电泳检测后,选择目标条带单一且明亮的PCR扩增产物送交上海美吉生物医药科技有限公司纯化并进行双向测序,测序引物为M13F(TGTAAAACGACGGCCAGT)及M13R(CAGGAAACAGCTATGAC).
1.5 序列分析与比对
将测序所得原始序列使用Chromas 2.6.4软件对峰图进行检查,并对碱基进行逐一校对. 由于样品采用双向测序,使用BioEdit 7.0.5软件进行序列拼接,以确保所用序列结果的准确性. 对所获得有效的序列在NCBI(National Center for Biotechnology Information)进行序列比对,同时下载相关的物种序列用于系统发育树构建. 序列比对时,参考Machida等[17]和Bucklin等[23]的研究分类标准,若序列相似度大于96%,则可鉴定至物种;若序列相似性介于83%~96%间,依据相似度的高低鉴定至属或科;若序列相似性小于83%,则归类至“门”或其他更高阶元.
1.6 系统发育分析
分别对桡足类、枝角类和轮虫3个类群物种序列开展系统发育分析. 使用MEGA X(10.0.5)分析所获得序列长度、碱基组成,采用 Kimura-2-Parameter双参数模型(K2P)计算遗传距离,使用邻接(Neighbor-Joining)法构建系统进化树,经1000次重复抽样(bootstrap)检验其置信度. 根据序列比对结果,选取每条序列相似度最高的序列作为参考序列,同时下载与比对结果相似度相近物种的序列及同源序列,其中桡足类47条、枝角类54条,轮虫38条序列分别构建进化树,进一步确定物种.
如图9(b)所示,新型扩孔钻头领眼段出现严重失效,主要有两方面的原因:其一,地层不均质性强,地层砾石含量高,钻头承受了较高冲击载荷,钻头心部布齿密度较低,个别切削齿失效从而导致钻头心部区域失效现象迅速发生;其二,扩孔钻头的横向不平衡力仍然存在,为了平衡这一钻头载荷所产生的侧弯矩,钻头回转中心与理想几何中心会产生一个偏转角度(如图10所示),这一偏转角度会使钻头心部切削齿产生额外的吃入深度Δh,其切削载荷大幅升高,最终导致钻头心部切削齿快速磨损。
2 结果
2.1 柳叶湖浮游动物休眠卵序列特征及比对结果
本研究共对191个休眠卵进行了分子鉴定,对测序结果进行拼接后成功获取101条COⅠ基因部分片段序列,单次成功率为52.88%. 扩增的COⅠ基因部分序列长度约为649 bp,序列无插入和缺失. 在所有序列比对结果中核苷酸T、C、A、G的平均含量分别为35.22%、17.78%、27.91%、18.98%,且A+T含量(63.13%)远高于C+G含量(36.76%),碱基均出现明显的偏倚性,符合线粒体碱基组成特点. 常德柳叶湖浮游动物休眠卵获得的DNA序列比对结果见表1. 本研究在191个休眠卵中所获得101条序列中,成功鉴定出17种休眠卵,其中11种鉴定至种,5种鉴定至属,1种鉴定至科;同时对成功鉴定的休眠卵进行分类并逐个编号,其中桡足类为CP1、枝角类为C1~C13、轮虫为Ro1~Ro3,同时记录该种休眠卵出现的样点及检出的个体数(表1、图2).
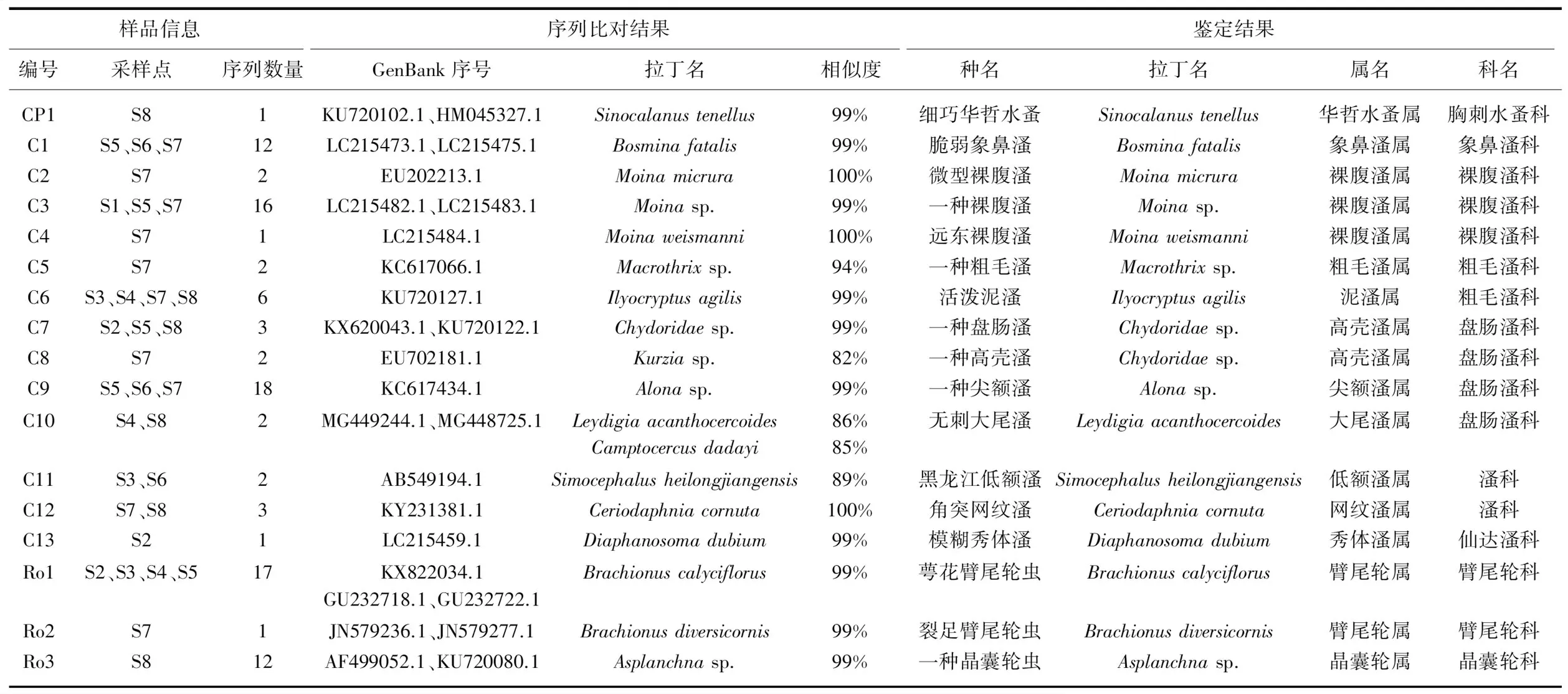
表1 条形码序列NCBI比对结果与样品鉴定结果的比较1)
1)单次成功率依次为:桡足类:47%;枝角类:57%;轮虫类:39%.
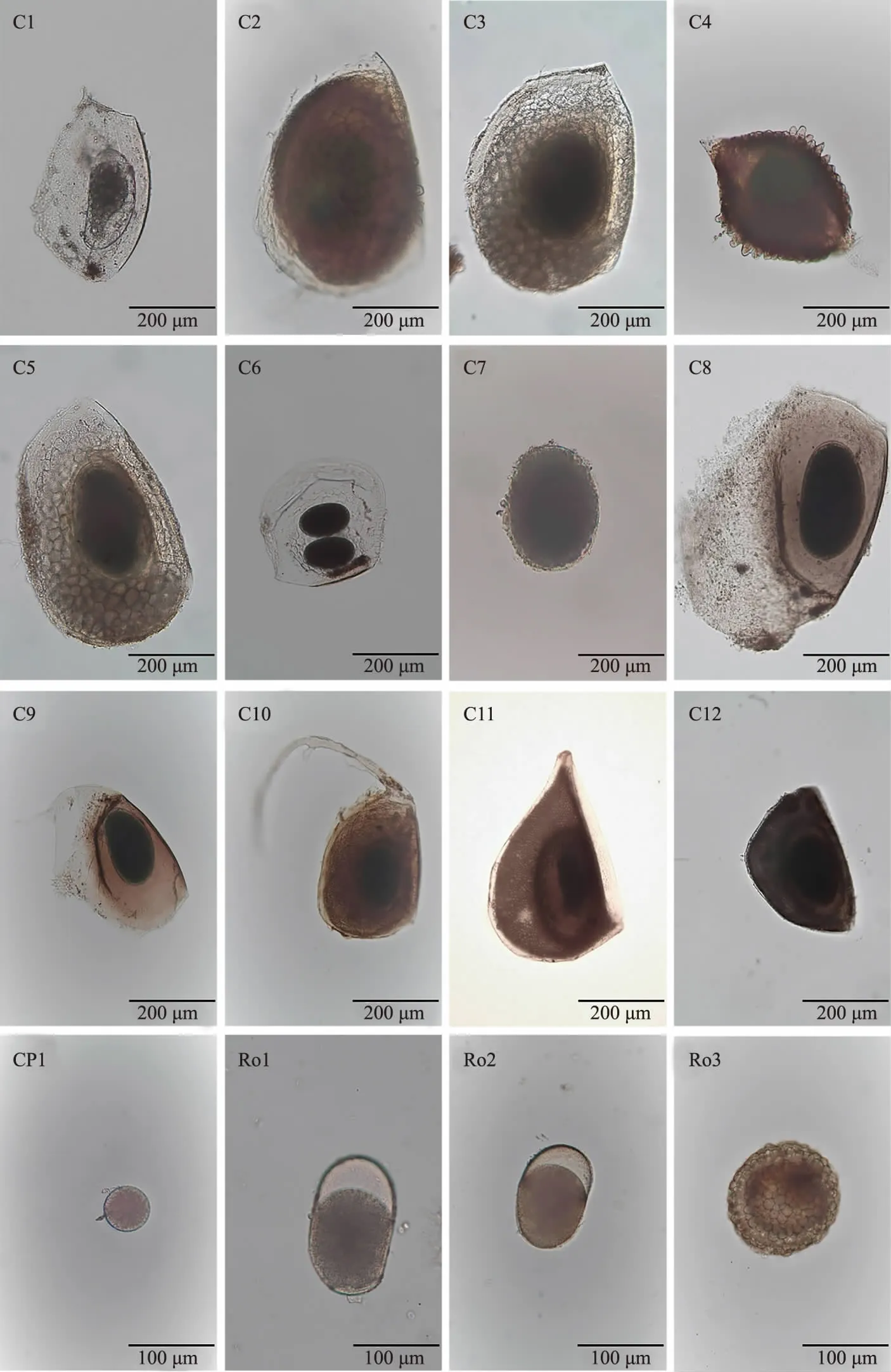
图2 休眠卵形态学信息Fig.2 Morphological information of resting eggs
样品C1、C2、C4、C6、C12、C13、CP1、Ro1、Ro2通过NCBI数据库比对可鉴定至种(表1);样品C3、C5、C9、Ro3可鉴定至属(表1);样品C7和C8鉴定至科,其中样品C8可能为高壳溞(Kurziasp.,序列相似度为82%);样品C10序列比对结果有两种,无刺大尾溞(Leydigiaacanthocercoides)和Camptocercusdadayi,相似度分别为85.9%和85.8%,同属盘肠溞科;样品C11比对结果为低额溞(Simocephalussp.),可能为黑龙江低额溞(S.heilongjiangensis,相似度88.5%).
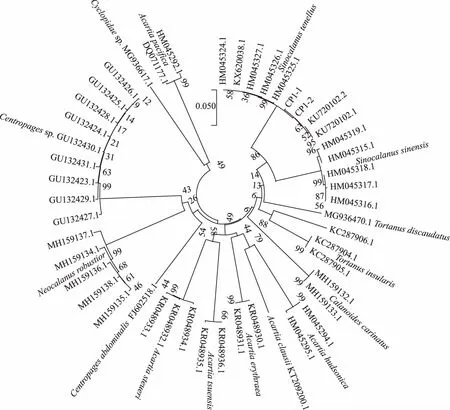
图3 基于邻接法构建的桡足类休眠卵COⅠ基因系统发育树Fig.3 Neighbor-Joining tree resulting from analysis of COⅠ gene data of copepod resting eggs
2.2 系统进化分析
本研究涉及3个类群,分别基于49个、69个、48个单倍型构建3条系统发育树(图3~5),其中桡足类系统发育树中鉴定序列为2条,下载相关同源序列为47条;枝角类系统发育树中鉴定序列为15条,下载相关序列为54条;轮虫系统发育树中鉴定序列为9条,下载相关序列为38条. 观察构建的Neighbor-Joining系统发育树显示柳叶湖浮游动物不同种类休眠卵间的界限明显,置信度较高(99%~100%)(图3~5). 同属物种均可聚类为一支,如:细巧华哲水蚤(Sinocalanustenellus)先于参比序列(HM045324.1、HM045326.1)聚为一支后与同属的中华华哲水蚤(S.sinensis)聚为一支,微型裸腹溞(Moinamicrura)首先与其参比序列(LC215484.1)聚为一支后与同属的远东裸腹溞(M.weismanni)聚为一支;萼花臂尾轮虫(Brachionuscalyciflorus)先与其参比序列(GU232718.1、GU232722.1)独立聚为一支,后与同属的裂足臂尾轮虫(B.diversicornis)聚为一支. 序列比对相似度较低的休眠卵C10、C11分别与无刺大尾溞和黑龙江低额溞的参比序列聚类至一起,可确定为相同物种.

图4 基于邻接法构建的枝角类休眠卵COⅠ基因系统发育树Fig.4 Neighbor-Joining tree resulting from analysis of COⅠ gene data of cladocera resting eggs
3 讨论
3.1 使用DNA条形码技术鉴定休眠卵的可行性及局限性
由于休眠卵传统鉴定较为困难,本研究用DNA条形码技术对柳叶湖表层沉积物中休眠卵种类进行初步鉴定,通过NCBI数据库比对,在物种水平上准确鉴定柳叶湖浮游动物休眠卵11种,构建了Neighbor-Joining系统发育树,发现所有物种均聚为独立分支,以属为分类阶元亦可在系统发育树中成功聚类,表明DNA条形码技术可成功应用于柳叶湖浮游动物休眠卵的种类鉴定. 研究过程中鉴定出许多其他物种的休眠卵或休眠体,如斑条透明苔虫(Hyalinellapunctata)、平角涡虫(Planocerareticulata)、长跗摇蚊(Tanytarsussp.)等,同时所测序列中未鉴定出原生动物.
该技术也存在一定局限性,例如本研究中只鉴定出一种桡足类休眠卵,原因可能是桡足类休眠卵个体小,形态种类多变,在体式显微镜下挑选单个体休眠卵进行分子鉴定时易与杂质混淆,造成鉴定遗漏. 另外轮虫种类休眠卵差异细微,无特殊形态特征,在对本研究中所挑选的未知休眠卵进行分子鉴定时,鉴定结果大多为萼花臂尾轮虫,其他种数量较少. 除部分浮游动物休眠卵体形易区分外,大部分休眠卵与沉积物颗粒或杂质相似,显微镜下不容易挑取.
3.2 研究中出现的问题与改进
首先,本研究休眠卵单次鉴定成功率约为52.88%. 镜检下许多有正常卵胚胎结构的完整卵体,由于产物浓度不达标、无扩增产物或PCR产物为假阳性,从而导致在后期PCR过程中鉴定失败. 究其原因,可能主要是个别物种COⅠ序列与引物某些位点不契合致使成功率偏低,因此有学者改进设计了兼并引物jgLCO1490/jgHCO2198[24]或针对某些特定种群设计巢式PCR引物[25];卵体处于不应期内,代谢速率较低,胚胎内线粒体较少,无法提取到足够的线粒体DNA;卵体表面携带大量病毒、细菌等[26-27],干扰COⅠ基因的特异性扩增,导致扩增产物主要为细菌DNA,从而表现为假阳性. 因此,当破碎枝角类的卵时应当将卵鞍剔除,以防止其携带的细菌或病毒对后续实验产生影响,或对于个体较大休眠卵DNA提取时可以采用“玻璃乳提取法”[28],以提高DNA提取的纯度及成功率. 不同样点休眠卵鉴定成功率不同,靠近湖心的样点成功率较高(50%~60%),而近岸的样点成功率较低(20%~30%). 可能是由于近岸表层沉积物中休眠卵受到某些环境因子影响使DNA受到降解,降低卵体DNA质量,例如Jiang等研究证实沉积物中的重金属含量会使太平洋纺锤水蚤(Acartiapacifica)休眠卵萌发率显著降低[29],并且表层沉积物中石油烃含量的增加会使太平洋纺锤水蚤休眠卵的存活率下降[30].
此外,若出现无法比对到具体物种的序列,可通过进一步系统发育树分析确定结果,如休眠卵C10、C11样品,样品通过系统发育分析分别与无刺大尾溞和黑龙江低额溞聚类至一支,可初步确定为相同物种. 本研究中休眠卵C3、C5、C9、Ro3样品鉴定至属,C7、C8仅能鉴定至科,并且研究中所获取多条序列在数据库比对中无相关信息返回,原因主要是NCBI数据库中无相似度高的参比序列,因而无法进行比对. 由于目前传统形态学鉴定仍是浮游动物鉴定的主要手段,但从事传统形态学鉴定的专家较少,鉴定主观性较强. 若将形态学鉴定和DNA条形码相结合,即在分类学专家帮助下,建立本地DNA条形码数据库,这必将极大地提升物种鉴定、群落动态监测的准确性[31-33].
目前国内针对浮游动物休眠卵鉴定工作开展较少,往往都是通过单一方式鉴定. 不同鉴定方式有不同的优缺点,应当将多种方式结合起来准确鉴定浮游动物休眠卵,以建立较完整的浮游动物休眠卵鉴定图库,方便进一步开展休眠卵研究工作. 此外NCBI数据库中浮游动物信息不够完善,在保证精确度的同时,应加大对NCBI数据库的上传力度. 随着测序技术的进步和DNA条形码技术的普及,我们相信DNA条形码技术将在物种鉴定中发挥越来越重要的作用.
4 结论
本研究采用DNA条形码技术对柳叶湖表层沉积物休眠卵种类进行鉴定,成功获取17种物种信息,初步建立了柳叶湖浮游动物休眠卵图谱. 在目前休眠卵传统形态学分类资料匮乏的情况下,COⅠ基因可作为浮游动物休眠卵鉴定的一种可靠、快捷、有效的方式. 但NCBI数据库对于物种的收录依旧不够完整和准确,导致本研究个别样品无法鉴定至种. 因此,比对数据库准确性和完整性亟待加强完善.
致谢: 感谢曾悦和刘陈在论文写作中给予的帮助.
