新吉细毛羊5 个生殖激素受体基因多态性分析
2019-12-26吴翠玲赵云辉柴浩晨张明新王春昕
吴翠玲,谢 平,赵 卓,赵云辉,翟 博,柴浩晨,3,张明新*,王春昕*
(1.吉林农业大学动物科学技术学院,吉林长春 130000;2.吉林省农业科学院,吉林公主岭 136100;3.西北农林科技大学动物科技学院,陕西杨凌 712100)
家畜繁殖性状的高低决定了畜牧业的经济效益,其中产仔数是家畜繁殖性状的重要数量性状。但绵羊产羔数又是一个遗传力低、难以改良的经济性状。绵羊繁殖性能主要由下丘脑-垂体-性腺轴(HPGA)调节[1]。下丘脑产生的GnRH与其受体GnRHR结合后调节垂体前叶LH、FSH激素的合成和释放。增加GnRH的分泌和GnRHR的数量会引起LH释放高峰,而LH释放高峰能促进哺乳动物排卵。其中LH与FSH化学本质是糖蛋白,LH可促进胆固醇在性腺细胞内转化为性激素。FSH能促进母畜卵泡、颗粒细胞生长和雌激素合成。LH和FSH与受体结合后促进卵泡生长成熟,诱导排卵,促进黄体生成[2]。催乳素(PRL)在垂体嗜酸细胞中合成和分泌,可促进母畜乳腺组织发育,调控雌激素分泌合成,抑制排卵。PRL 与其受体PRLR 结合后,通过JAK2/STAT5 信号系统传导信息,保证目标基因的正常表达。雌激素主要由卵巢的颗粒细胞和卵泡膜细胞分泌,其受体ESR广泛存在雌性哺乳动物组织中,雌激素与ESR 结合后与FSH、LH协同调控动物发情。因此HPGA 中的生殖激素及其受体是影响绵羊产羔率的重要因素,可以作为影响绵羊产仔数的重要候选基因[3-4]。
新吉细毛羊是新疆畜牧科学院、新疆农垦科学院和吉林农业科学院联合培育的细毛羊品种,具有净毛率高、羊毛综合品质优异、适应性强、遗传性能稳定等优点,但新吉细毛羊产羔率只有115%~120%。提高新吉细毛羊繁殖力是目前待解决的关键问题。本研究对新吉细毛羊GnRHR、LHR、FSH、PRLR、ESR5 个基因多态性进行分析,旨在为新吉细毛羊育种工作中提高繁殖力提供理论依据。
1 材料与方法
1.1 实验动物、试剂及仪器 在吉林省查干花种畜场采集新吉细毛羊119 只母羊血样各7~10 mL,-20℃保存。收集整理119 只新吉细毛羊头三胎产羔数据;血液基因组柱式小量提取试剂盒购自康为世纪生物技术有限公司,PCR Mixture 购自天根生物科技有限公司;主要仪器包括核酸蛋白检测仪(Quawell,美国)、电泳仪(北京六一)、PCR 仪(伯乐,美国)等。
1.2 实验设计和方法
1.2.1 PCR 扩增 用血液基因组柱式小量提取试剂盒提取DNA。根据NCBI 登录号利用primer 5 软件对5 个基因外显子区设计引物,在上海生工生物工程股份有限公司合成引物,引物序列及相关信息见表1。PCR 反应体系:PCR Mixture10 μL,上下游引物(10 mol/L)各0.5 μL,DNA 模板1.0 μL,加蒸馏水至20 μL。PCR 反应条件:95℃预变性3 min,95℃变性30 s,退火30 s,退火温度延伸1 min,34 个循环;72℃延伸5 min。4℃保存。PCR 产物用1.5%的琼脂糖凝胶电泳检测。
1.2.2 测序与统计分析 将每个基因PCR 产物分别混池送上海生工生物工程股份有限公司进行测序,用DNASTAR 软件对测序结果分析。对存在突变的基因片段PCR 产物分别测序。利用Excel 计算基因型频率。利用GraphPad prism 6 软件进行基因多态性与产羔数相关性分析。
2 结果与分析
2.1 DNA 和PCR 产物检测结果 提取的基因组DNA经核酸蛋白检测仪检测OD260/280 值为1.8~2.1。说明提取的DNA 纯度与质量满足后续实验要求。基因PCR产物结果见图1。由图1 可知,得到产物与预期片段大小一致,无杂带,符合后续实验的要求。

图1 5 个基因PCR 产物检测结果
2.2 测序结果分析 测序发现FSHR基因不存在SNP 突变位点。GnRHR、LHR、PRLR、ESR基因突变位点统计结果见表2,4 个基因分型的测序结果如图2~5。GnRHR基因存在2 个SNP 为cDNA 区发生的A505G、G720C突变,氨基酸分别发生G720C、R240S 改变。LHR基因存 在5 个SNP 为cDNA 区发生的T1262G、A1991G、C2012T、A2032T、T2041A 的突变,氨基酸分别发生D421A、Y664C、S671L、T678S、L681M 改 变。PRLR基因存在2 个SNP 为cDNA 区发生的A1263G、C1544G突变,氨基酸分别发生K387E、S480R 改变。ESR基因存在4个SNP为cDNA区发生的A106T、C181T、G612A、C735T 突变,其中A106T 突变导致氨基酸发生I36F 改变。后3 个为同义突变,未导致氨基酸的改变。
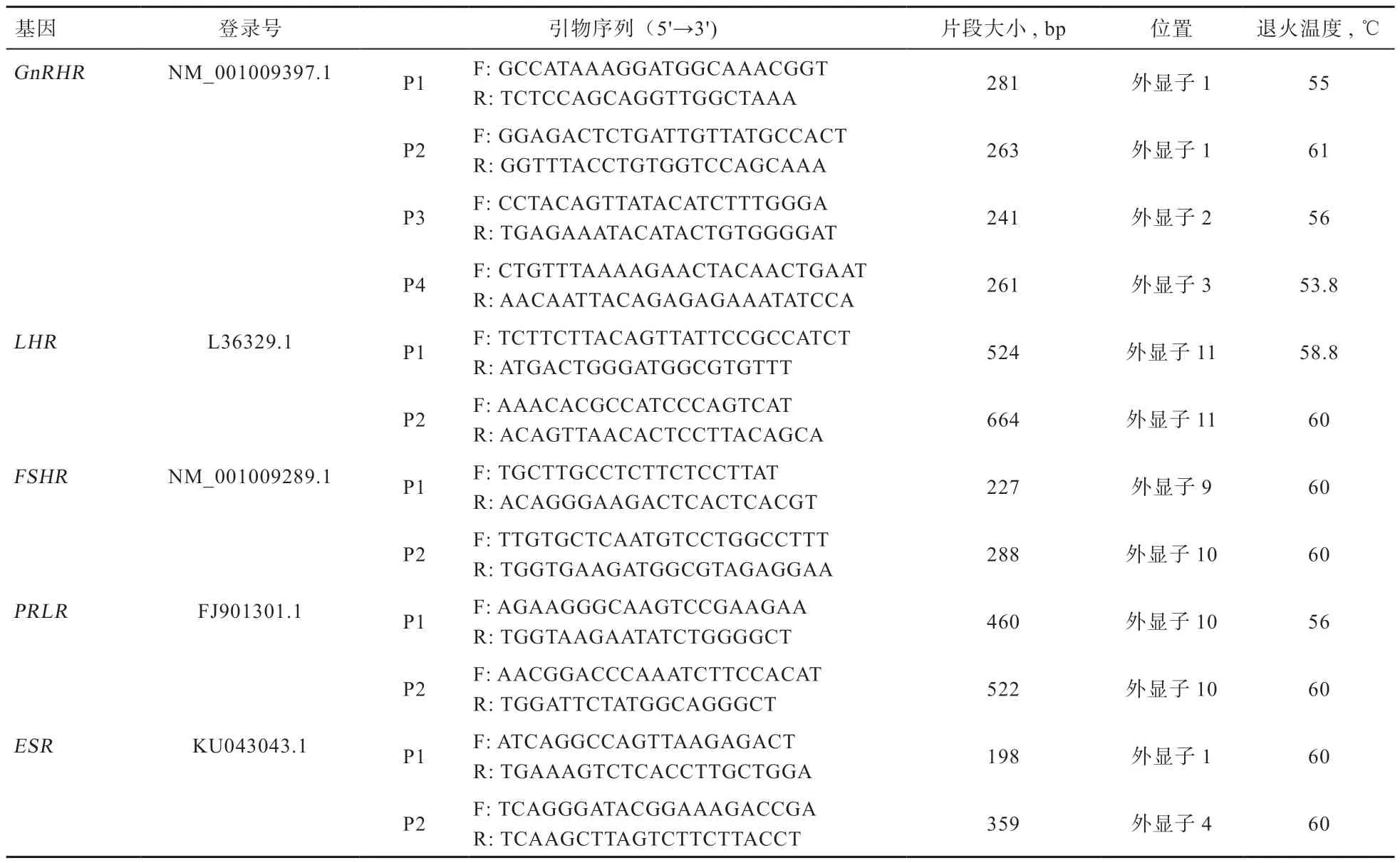
表1 引物信息表
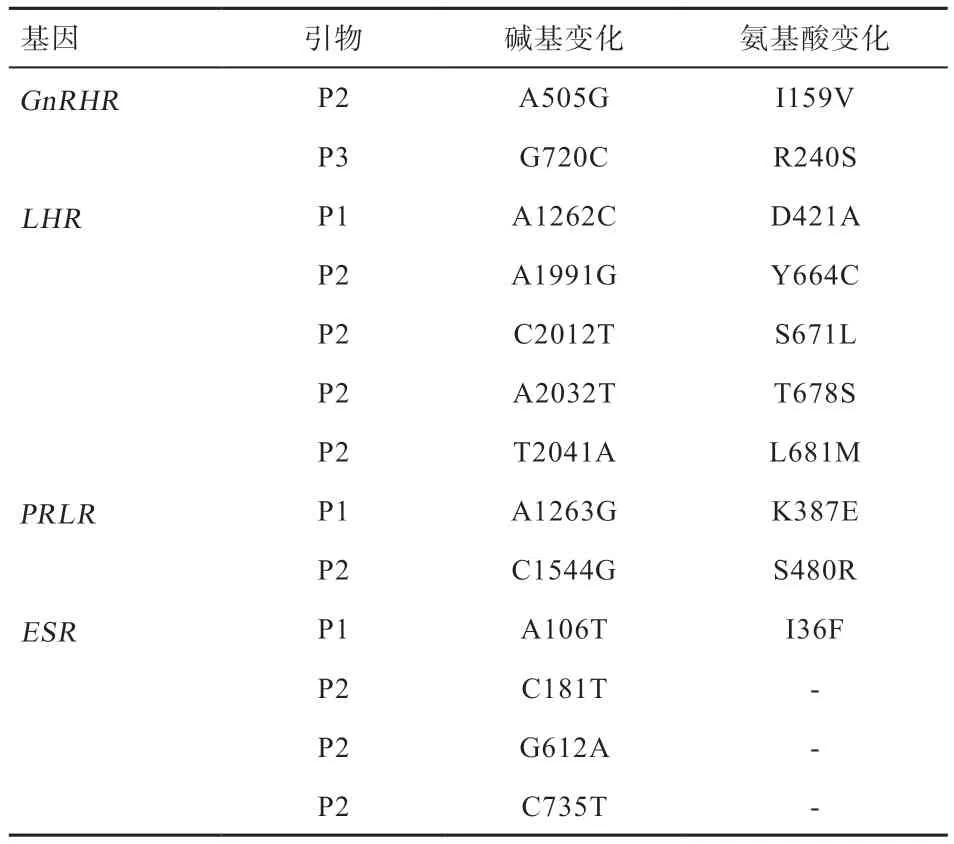
表2 4 个基因突变位点信息表
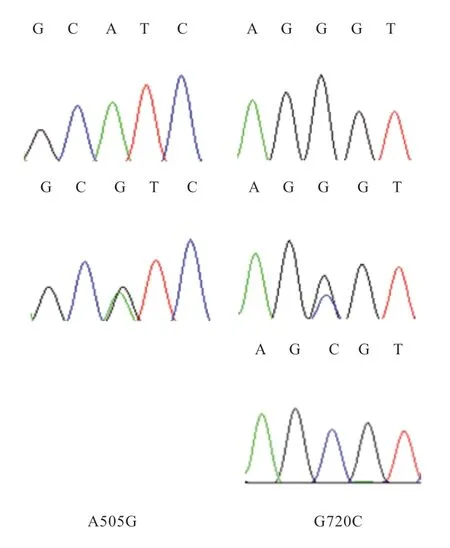
图2 GnRHR 基因测序结果

图3 LHR 基因测序结果

图4 PRLR 基因测序结果
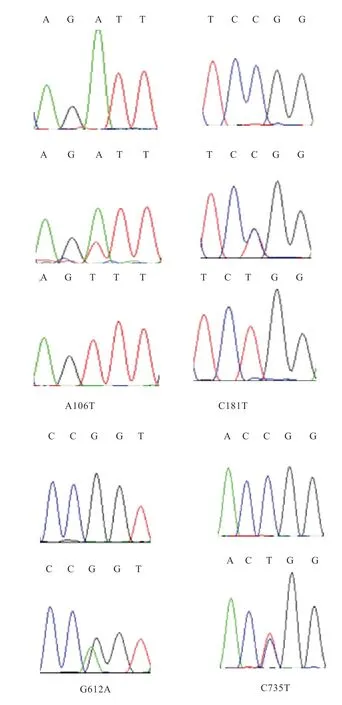
图5 ESR 基因测序结果
2.3 统计分析 新吉细毛羊GnRHR、LHR、PRLR、ESR4 个基因多态位点的基因型、基因型频率计算结果以及不同基因型对新吉细毛羊产羔数的影响见表3。由表3 可知,除了GnRHR基因A505G 突变,LHR基因C2012T、A2032T 的突变,PRLR基因A1263G 突变,ESR基因C735T 突变只有2 种基因型,其他突变位点有3 种基因型。在新吉细毛羊群体4 个基因多态性与产羔数相关性分析结果中,只有ESR基因A106T 突变对新吉细毛羊产羔数影响显著。其中 AA、AT、TT 3 种基因型个体的平均产羔数分别为1.391、1.101、1.417。AA 基因型、TT 基因型平均产羔数比AT 基因型分别多0.290、0.316 个(P<0.05)。AA 型、TT 基因型平均产羔数之间差异不显著(P>0.05)。
3 讨 论
数量性状的表型值是由基因型值和环境值共同作用的结果。遗传效应是产生表型变异的内因,环境效应是表型变异的外部原因。近年来,为提高绵羊产羔数,给养羊业生产实践、选种选配提供理论依据。国内外从遗传效应方面开展了大量多胎候选基因多态性的研究[5]。本实验研究了5 个关键的生殖激素受体基因:GnRHR、LHR、FSHR、PRLR、ESR。目前研究发现GnRHR基因对黄淮山羊、济宁青山羊、古蔺马羊、川东山羊、西农萨能奶山羊、贵州白山羊产羔数影响显著[6-7];FSHR基因对波尔山羊、贵州黑山羊、辽宁绒山羊、萨能奶山羊产羔数影响显著[8];PRLR基因对中国美利奴(新疆型)多胎品系、小尾寒羊、关中山羊、波尔山羊、济宁青山羊[9-10]产羔数影响显著;ESR基因对小尾寒羊、湖羊、济宁青山羊产羔数影响显著[11]。
3.1GnRHR基因多态性分析GnRHR是G 蛋白耦联受体家族成员,在垂体表面、淋巴细胞、乳腺、卵巢和前列腺中表达。GnRHR基因是分子标记的研究热点之一。国内外对GnRHR基因多态性进行了大量的研究[7,12]。本研究对GnRHR基因的全部外显子共设计了4 对引物,在外显子2、外显子3 分别发现一个错义突变。其中G720C 突变位点只有一个样本为CC 型,虽产羔数为2,但不具有统计学意义。其结果可能存在偶然性,CC 型也有可能是新吉细毛样双羔性状的标记,需加大样本量,进一步验证。
3.2LHR基因多态性分析 目前的研究中,对绵羊LHR基因多态性研究较少,大多集中在LHβ基因上。因为LH基因的β亚基具有激素和物种特异性,识别、结合靶组织的特异受体,决定激素的生理特异性。但激素发挥作用必须依赖受体。LHR基因外显子10、外显子11 组成编码区,本实验中主要对LHR基因外显子11 多态性研究,发现了5 个SNP 错义突变位点,均与新吉细毛羊产仔数无显著相关。王利红[13]利用PCRSSCP 技术对湖羊、晋中绵羊和陵川半细毛羊LHR基因外显子11 的多态性进行了研究,虽发现4 个突变位点,其中3 个突变位点A1991G、A2032T、T2041A 与新吉细毛羊上发现的相同,但均未对绵羊产仔数产生显著影响,其结果与本实验研究结果相似。推测LHR不是新吉细毛羊产羔性状的主效基因。
3.3FSHR基因多态性分析 国内外大量研究表明FSHR基因多态性对动物产羔数具有重要影响。本研究主要对FSHR基因外显子9、外显子10 多态性分析,共设计3 对引物,其中对FSHR基因外显子10 设计2对引物,第一对引物PCR 产物全长1 271 bp,覆盖全部外显子。由于PCR 产物过长,未在实验结果图1 中表示。测序结果表明3 对引物的PCR 产物均未发现突变位点。除了FSHR基因的外显子9、外显子10 以外,外显子7 与5'端调控区也是该基因的研究热点[14]。为最大挖掘新吉细毛羊多胎基因,应对FSHR基因其他区域进行深入研究。
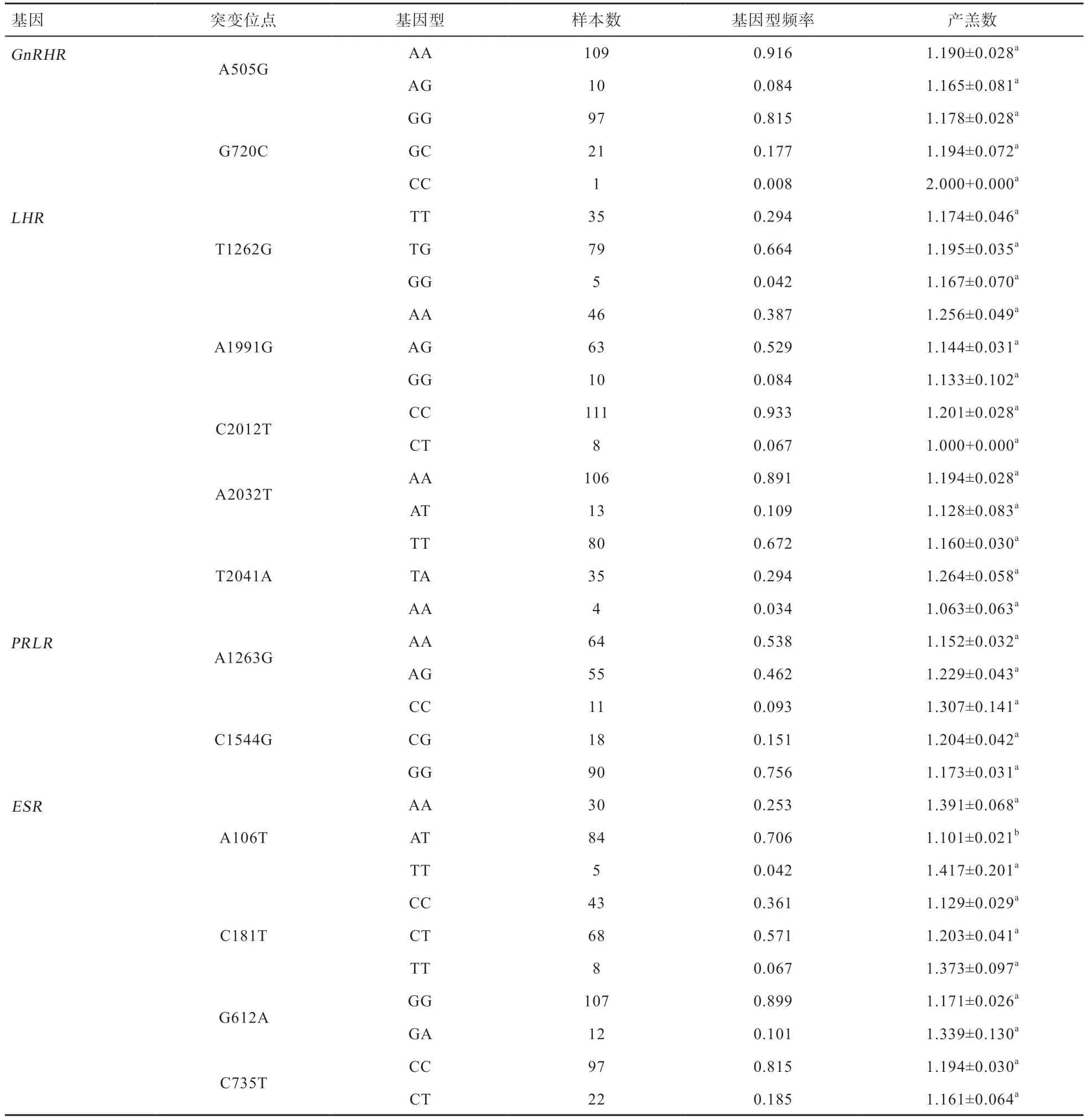
表3 新吉细毛羊4 个基因的基因型频率、对产羔数影响
3.4PRLR基因多态性分析PRLR是细胞因子受体家族成员。目前国内外研究将PRLR基因作为猪繁殖性能的一个主效基因。猪的PRLR基因存在Alu I 酶切位点,其等位基因A 公认为猪产仔性状的优势基因型[15]。除此之外,PRLR基因与FSHβ、ESR、RBP4基因互作对猪繁殖性能有显著影响。本研究对新吉细毛羊的PRLR基因外显子10 多态性进行了分析,共设计两对引物,在引物P1、P2 的PCR 产物中各发现一个错义突变:A1263G、C1544G。在A1263G 突变中,不存在GG 基因型。张向楠利用PCR-SSCP 技术检测4 个绵羊群体PRLR基因外显子10,发现A1263G 突变存在GG 基因型,其基因型频率极低。该结果与本研究结果相似,不同基因型对产羔数影响差异不显著[16]。在C1544G 突变中,虽然不同基因型对新吉细毛羊产羔数差异不显著。但产羔数趋势为CC 型>CG 型>GG 型。吴洪宾利用PCR-SSCP 技术在中国美利奴羊中发现相同突变位点,其GG 型3 月龄出生重比CC 型大约重1.7 kg。但产羔数趋势与本研究结果一致,CC 型产羔数比GG 型约多0.3 只[17]。推测C 等位基因是绵羊多胎性状优势基因。
3.5ESR基因多态性分析ESR基因被公认为猪繁殖性状的主效基因。ESR基因的Pvu II 多态位点与猪的产仔数存在紧密联系[18]。绵羊中对ESR基因研究主要集中在外显子1 和外显子4 上。因此本研究对ESR基因外显子1 和外显子4 多态性进行分析,共设计2 对引物。在P1 的PCR 产物中发现一个错义突变A106T。P2的PCR 产物中发现3 处同义突变:C181T、G612A、C735T。在A106T 的研究中发现AA 基因型与TT 基因型比AT 基因型产羔数约多0.3 只(P<0.05),推测AA 型和TT 型为优势基因型。
综上所述,在5 个生殖激素受体基因中发现的突变位点对新吉细毛羊的产羔数无显著影响。本实验室前期研究发现新吉细毛羊BMPR-1B基因、BMP15基因、GDF9基因多态性对产羔数也无显著影响[19-20]。同时单一基因的辅助育种效率低、世代间隔长,已经不能满足目前养羊业育种的需求。为提高新吉细毛羊产羔率,将进一步在引进多胎基因、基因联合、幼雏超排[21]等方面提高产羔率。
4 结 论
本研究发现FSHR基因不存在突变位点,其余4 个基因存在13 个突变位点。GnRHR基因存在A505G、G720C 突变,LHR基因存在T1262G、A1991G、C2012T、A2032T、T2041A 突变,PRLR基因存在A1263G、C1544G突变,ESR基因存在A106T、C181T、G612A、C735T突变。在新吉细毛羊群体4 个基因多态性与产羔数相关性分析结果中,只有ESR基因A106T 突变对新吉细毛羊产羔数影响显著。其中AA、AT、TT 3 种基因型个体的平均产羔数分别为1.391、1.101、1.417。AA 基因型、TT 基因型平均产羔数比AT 基因型分别多0.290、0.316个。AA 基因型、TT 基因型平均产羔数差异不显著。
