Ds插入突变体的遗传学综合性实验设计与探讨
2019-12-24李楠李亚娟郭海滨张向前
李楠,李亚娟,郭海滨,张向前
插入突变体的遗传学综合性实验设计与探讨
李楠1,李亚娟1,郭海滨1,张向前2
1. 华南农业大学公共基础课实验教学中心,广州 510642 2. 华南农业大学林学与风景园林学院,广州 510642
学生创新能力的培养已成为中国高等教育的重要导向,加强综合设计性实验的应用是提高大学生创新能力的有效途径。文章以水稻插入突变体为实验材料,以突变体表型观测作为实验切入点,以遗传分析和转座检测为实验学习重点,重新设计了一个分子遗传学综合性实验项目。在此基础上,通过知识拓展环节引导学生利用开放实验平台学习TAIL-PCR技术,进一步进入研究性实验。通过综合性实验到研究性实验的递进式教学体系,能够使学生深入理解表型与基因的内在联系、跳跃基因的概念以及转座子在基因功能研究中的重要作用,加深学生对基因概念及遗传规律的认识和理解,提升学生理论知识与实验技能的整合运用能力。
综合性实验;实验教学;基因突变
遗传学是探究生物遗传和变异规律的理论科学,是生命科学领域的基础学科,研究内容丰富,小到基因结构、功能及表达,大到生物群落的遗传特征[1]。遗传学理论与实践关系密切,实验不仅是遗传学的重要基础,更是培养创新能力的重要环节[2]。为提升遗传学实验教学质量和促进创新人才培养,华南农业大学于2007年构建了“一般性实验—综合性实验—研究性实验”的递进式遗传学实验教学新体系,提出适合华南农业大学农科类专业的“创新型混合教学模式”[3]。2012年,华南农业大学实现了遗传学实验教学的独立设课,新编《遗传学综合性实验》教材和实验教学大纲,将遗传学实验从16学时的一般性实验调整至32学时的普通遗传学实验和48或64学时的遗传学综合设计性实验。
当前华南农业大学本科遗传学实验教学主要分为3大类综合性实验项目:细胞遗传学综合性实验、经典遗传学综合性实验及分子遗传学综合性实验。分子遗传学综合性实验从2012年应用至今已有7年,以籼粳水稻叶片为材料,利用特定SSR引物扩增籼粳水稻的DNA片段,实现对不同材料的区分。该实验主要以植物DNA提取、PCR扩增和凝胶电泳检测等分子标记相关技术的学习为主,已远不能满足创新型人才培养需要。
为提升遗传学综合设计性实验项目的质量,适应当代教学的需要,利用水稻插入突变体材料,以培养学生科研思维和研究方法作为实验教学目标,以遗传分析法和PCR检测技术为学习重点,重新设计了分子遗传学综合性实验。使学生通过本实验项目的学习,能够理解表型与基因的内在联系,了解跳跃基因的概念以及转座子的重要应用价值,掌握科学研究的思维方式与研究方法,提升对理论与实践、知识与技术的整合运用能力。
1 实验材料的背景
粒重是决定水稻产量的3个主要因素(即每株有效穗数、每穗粒数和粒重)之一,是粒长、粒宽和粒厚的综合指标。而水稻粒长、粒宽及长宽比又称为水稻谷粒形状,是衡量稻米外观品质的重要指标之一,同时还影响稻米的商品品质和加工品质(糙米率、精米率和整精米率)。近年来,水稻粒形相关基因的研究取得了突破性进展。目前,至少已有93个水稻粒形相关基因被克隆,遍布水稻12条染色体[4~7]。本实验所用的突变体材料是一个转座引起的粒型突变体,其谷壳大小与野生型无异,而颖果显著缩小,该突变体表型为不完全显性遗传;利用转座子标签法,采用TAIL-PCR[8](thermal asymmetric interlaced PCR)技术已获知候选基因位于水稻第11号染色体,该基因属于类伸展蛋白基因家族,是一个控制水稻重要农艺性状的新基因。
该材料是由/双因子转座系统转化至粳型品种中花11号获得[9],其中用于转化的质粒为pDsBar1300,用于转化的质粒为pUbiTs (图1)[9]。pDsBar1300质粒的T-DNA区中携带转座子,的内部插入基因,能提供对Basta (商用除草剂,有效成分为膦丝菌素PPT)的抗性,在的旁侧插入潮霉素磷酸转移酶基因(hygromycin phostransferase,),能提供对潮霉素的抗性。将筛选得到的水稻转化纯合体和转化纯合体进行杂交,构建了多个杂交组合,通过转座酶的反式活化使发生转座[9]。
2 实验设计
2.1 设计思路
利用该材料进行综合性实验设计,先将以上科学研究的实验材料与野生型杂交获得F1,并自交获得F2分离群体,以该突变体、野生型、F1杂合体、F2群体的成熟谷粒及对应水稻叶片作为课程实验材料。首先让学生通过观察与考种实验发现突变体、野生型和F1杂合体在谷壳和米粒上的异同;其次提供F2群体米粒,通过米粒宽度测量及方差分析,引导学生对该材料进行表型观察和遗传分析;再次提供学生F2群体水稻叶片,利用PCR检测技术,分析突变体表型与插入之间的关系,验证表型调查结果。最后通过知识拓展环节,结合理论教学让学生深入了解转座因子在基因功能研究中的应用价值。并以突变体为例,探讨如何实现该类型材料(例如T-DNA或转座子插入突变体)的基因克隆及功能分析。该实验整个教学设计基于问题引导式教学模式(problem-based learning),以问题为导向,引导学生在实验中发现问题、分析问题、提出问题、研究问题并解决问题,最后得出科学研究结论,进而培养学生科学研究的思维方式,提升学生对理论与实践、知识与技术的整合运用能力和创新实践能力。
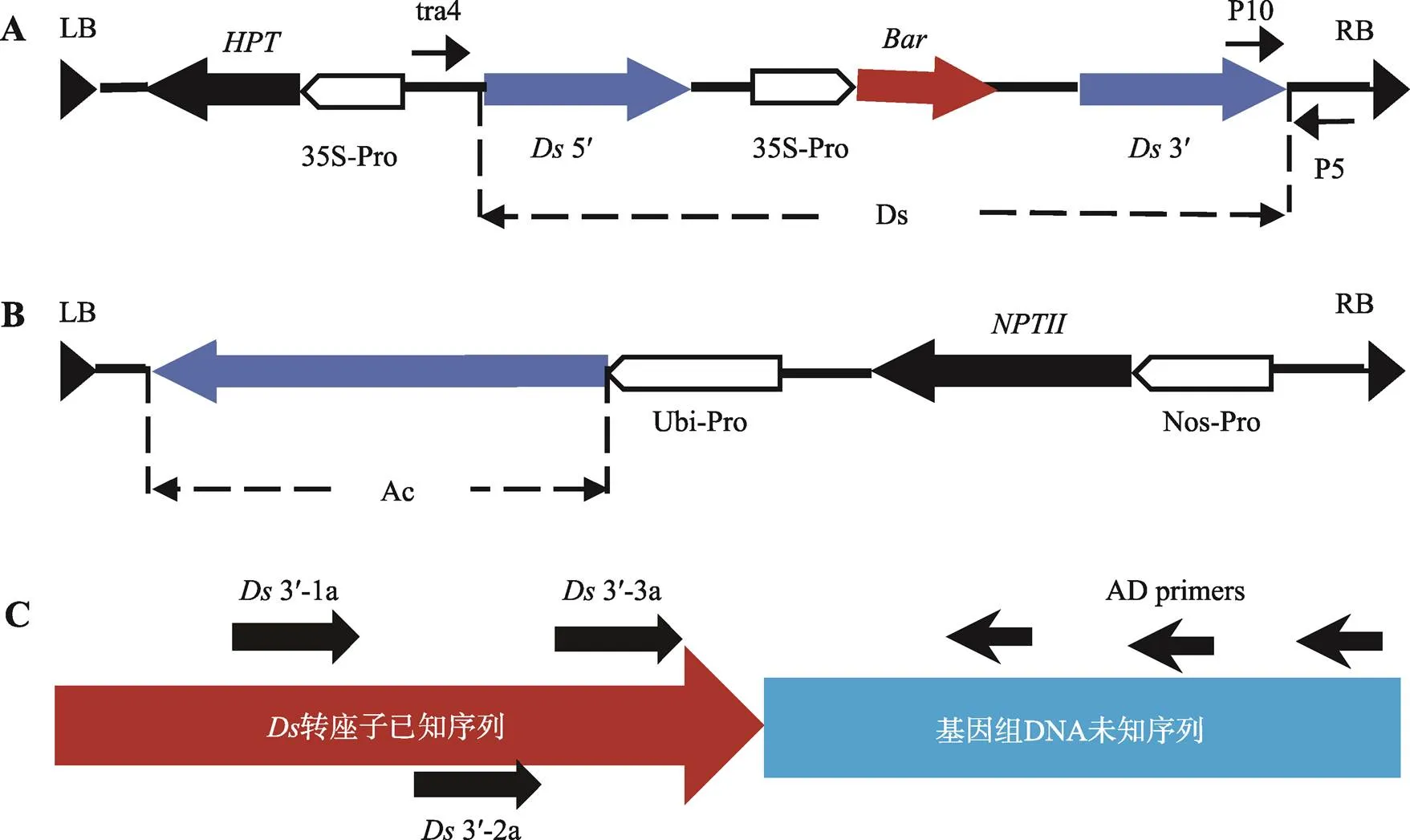
图1 质粒T-DNA区和TAIL-PCR引物所在位置示意图
A:质粒pDsBar1300中T-DNA区及PCR检测引物位置示意图。:潮霉素抗性基因;35S-Pro:35S启动子;:基因能提供对Basta的抗性;tra4、P5和P10:转座检测的3条引物。B:质粒pUbiTs中T-DNA区示意图。Ubi-Pro:Ubi启动子;Nos-Pro:Nos启动子;:基因编码新霉素磷酸转移酶,能赋予细胞抗卡那霉素的能力。LB和RB:T-DNA的左右边界。C:TAIL-PCR引物所在位置示意图。
2.2 实验涉及的主要理论知识与实验技术
该实验涉及的理论知识点与实验技术主要包括孟德尔遗传定律、转座子及TAIL-PCR技术。分离定律是孟德尔遗传定律之一。在杂合子细胞中,位于一对同源染色体上的等位基因,在进行减数分裂时等位基因会随着同源染色体的分开而分离,分别进入两个配子中,独立地随配子遗传给后代,在F2代性状发生3∶1或1∶2∶1的分离[10]。
转座子(transposon)是染色体上可复制和位移的一段DNA序列。转座子可以通过切割、重新整合等一系列过程从基因组的一个位置“跳跃”到另一个位置。在各种功能基因组学研究方法中,利用转座子插入诱变的方法被认为是进行大规模基因功能鉴定的有力工具[11],目前已有多例用/转座系统成功分离水稻基因的报道[12,13]。
TAIL-PCR技术,即交错式热不对称PCR,通过利用已知的DNA序列设计一组特异性引物,并结合随机简并引物,采用高温特异性扩增与低温随机扩增相结合的方法,获得转座子插入侧翼区特异扩增片段,相应扩增片段可直接测序分析,筛选分离基因。
2.3 实验材料与方法
2.3.1 实验材料
将突变体与野生型杂交,并将F1自交获得的F2群体,以突变体、野生型、F1杂合体及F2群体的成熟谷粒,F2群体谷粒对应的水稻叶片作为课程实验材料。所有材料均种植于华南农业大学农场,单株种植,常规管理。
2.3.2 实验方法
2.3.2.1 突变体的鉴定和遗传分析
一个实验班人均约30人,将学生以5人为一组,提供给每组同学野生型、突变纯合体、突变杂合体谷粒各1袋,每袋100粒。F2群体210株对应收获种子210袋,每袋100粒。考种方法为10粒为1组,3次重复,整齐摆放,配置游标卡尺与分析天平测量。利用ImageJ软件获取考种数据,并运用SPSS18.0软件进行卡方检验和t检验。
2.3.2.2转座的PCR检测
转座的PCR检测参照刘芳等[9]方法。因子切离的PCR检测利用引物P10、Tra4和P5同时扩增。如果未发生切离,称为满载供体位点(full donor site, FDS),通过引物P10和P5可以扩增出约400 bp的特异带;如果从T-DNA上切离,称为空载供体位点(empty donor site, EDS)引物Tra4和P5可以扩增出约870 bp的特异带;如果既存在EDS又存在FDS,则利用3条引物可以同时扩增出两条特异带。
具体实验步骤包括基因组DNA提取、PCR扩增、电泳分离、数据统计与分析等。每年在田间种植两亲本各20株、F120株和F2群体210株。待上课前1周取F2群体每株上部叶片置于超低温冰箱中保存备用。
水稻DNA提取:取2~4 cm长的叶片用液氮研磨至粉末,加入TPS抽提液(100 mmol/L Tris-HCl pH8.0,10 mmol/L EDTA,1 mol/L KCl)1000 μL,75℃水浴30 min;13 000r/min离心12 min,吸上清约500 μL转入1.5 ml离心管中,加入等体积遇冷的异丙醇,−20℃放置10 min,13 000 r/min离心5 min,弃上清,干燥沉淀,加200 μL灭菌水溶解,4℃冰箱冷藏备用。
PCR扩增及琼脂糖凝胶电泳检测:学生使用自己提取的基因组DNA为模板进行PCR扩增,每个模板使用3条引物(P10、Tra4和P5)同时扩增。扩增后的PCR产物用1%的琼脂糖凝胶进行电泳,在凝胶成像系统上观察电泳结果,记录样品基因型。
2.3.2.3 突变体基因克隆
利用水稻的/双因子转座系统,筛选转座插入纯合体植株为材料,采用TAIL-PCR[8]的方法扩增元件旁侧的水稻基因组序列,分析插入位点旁邻序列,通过序列比对确定该基因在水稻染色体上的具体位置,并对候选基因进行了分析。TAIL-PCR反应所用引物见表1,其位置示意图如图1所示,特异扩增得到的第3轮PCR产物用来测序。在水稻基因组中的插入位置运用BLAST (http://www.ncbi.nlm.nih.gov/BLAST)分析工具完成。因子插入所在位点相关基因预测信息可以在水稻基因组注释数据库获得(http://rice.plantbiology.msu. edu/)。
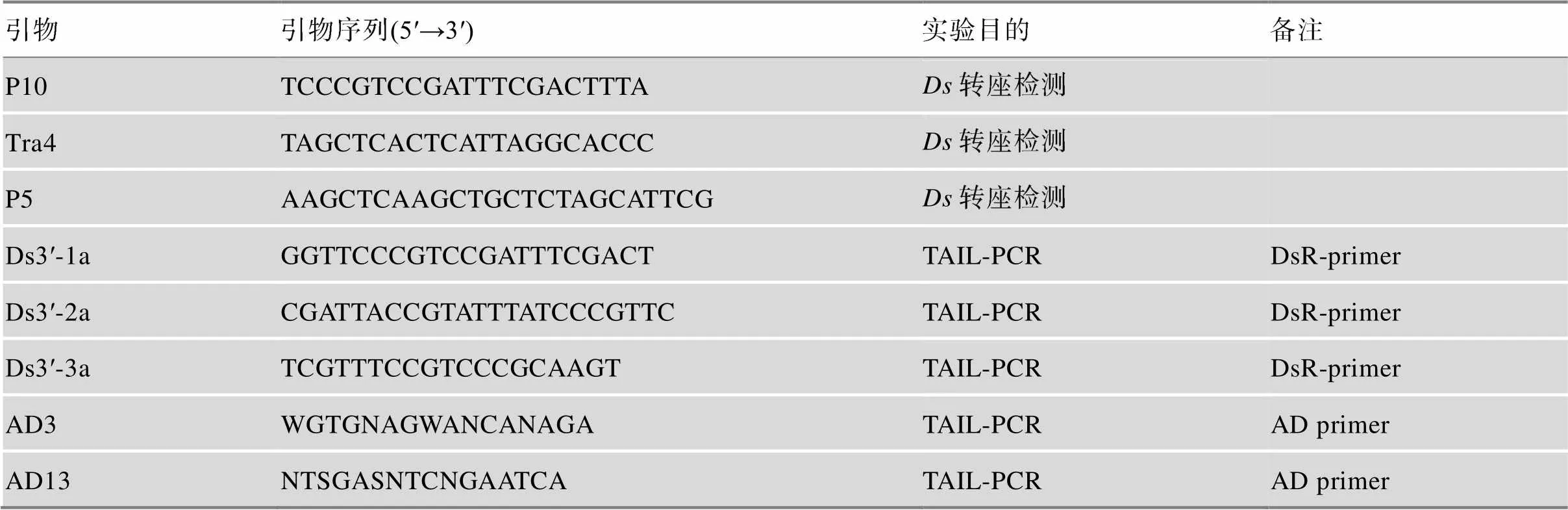
表1 实验所用引物
DsR-primer:元件3′末端特异的嵌套引物;AD primer:熔解温度较低的随机简并引物;W:“A/T”碱基;N:“A/C/G/T”碱基;S:“C/G”碱基。
2.4 实验结果与讨论
2.4.1 突变体鉴定与遗传分析
水稻是1个转座子插入引起的颖果明显变小,而谷粒大小没有改变的突变体(图2A)。进一步分析表明,突变纯合体颖果(20.25±0.26 mm,10粒)比野生型颖果宽(27.10± 0.43 mm,10粒)减少了25.3%,而其百粒重较野生型减少了33.6%;杂合体颖果比野生型宽度减少了18.5%,百粒重减少了25.2%。
为了研究突变体表型遗传特性,我们构建了F2分离群体。在F2群体中,不同植株的颖果表型明显不同,其中60株颖果宽度类似野生型,103株颖果宽度达22.09± 0.21 mm (10粒),47株颖果宽度则与突变纯合体无异。卡方测验表明其分离比符合1∶2∶1 (2= 1.68<20.05,2= 5.99),说明突变表型由1对不完全显性基因调控。
2.4.2 突变性状与共分离分析
为了分析突变体表型与之间的关系,并验证表型调查结果,对F2分离群体做了转座的PCR检测(图2B)。在F2代的210个单株的分离群体中,60株野生型植株均为未转座类型,47株突变体植株为转座纯合类型,其余为转座杂合类型,三者之间的比率同样符合1∶1∶2。根据上述结果可以认为,该突变体的产生是由于转座的插入所引起的,遗传行为符合1对不完全显性基因的分离模式,表明该突变表型与插入共分离,与F2群体颖果表型调查结果相吻合。
3 实践教学中的课堂组织方式
3.1 以科研思路为导向,创新实验内容、教学方法与组织方式
本实验作为分子遗传学综合性实验项目以培养学生创新思维与方法作为教学目标,将研究中发现的水稻插入粒型突变体作为实验材料,以遗传分析和PCR检测技术为实操学习重点,并通过以下4步来完成实验教学:(1)引导学生发现问题;(2)通过表型调查和方差分析分析问题;(3)将表型观测与PCR检测技术相结合,对突变体进行鉴定与遗传分析提出问题;(4)通过知识拓展环节,结合转座因子内容,以该材料为例深入讲解转座子的特征及应用价值,加深学生对跳跃基因概念的理解,进一步研究问题。
本实验可分4次课完成,每周1次课,每次课4个学时,分为以下3大部分:
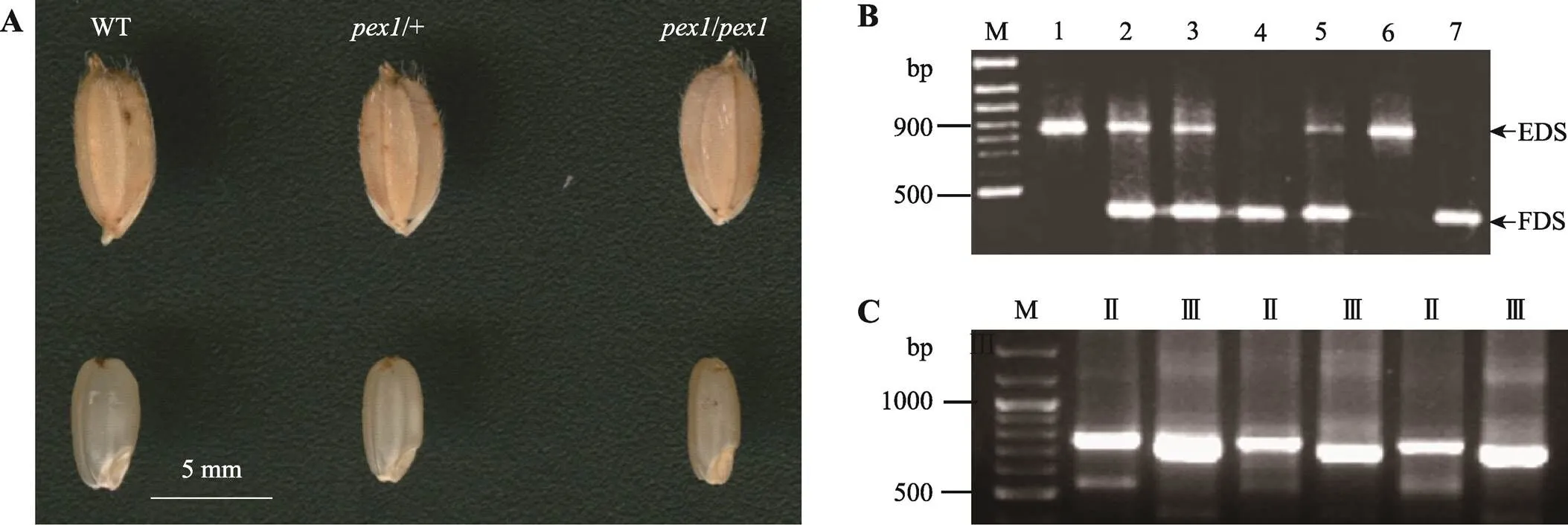
图2 水稻pex1突变体表型及其分子检测
A:水稻突变体谷粒和颖果表型。WT:野生型;:突变杂合体;:突变纯合体。B:转座的PCR检测。M:100 bp DNA Ladder;1~7:检测植株,其中植株4、7的未发生转座,仅存在FDS,1和6为转座纯合体,仅存在EDS;2, 3, 5为转座杂合体,既存在EDS又存在FDS。C:插入突变体TAIL-PCR产物琼脂糖电泳分析。M:100 bp DNA Ladder; Ⅱ, Ⅲ:分别为TAIL-PCR第二、三轮产物。
(1)遗传分析实验部分1次课。1~2学时:首先提供给学生野生型突变体谷粒材料,引导学生对谷壳和米粒进行仔细观察并发现问题。教师再讲解考种的方法,指导学生用统计方法研究突变体表型特征。3~4学时:提供给每组F2群体谷粒,要求学生测量谷粒表型,利用卡方检验对突变体进行遗传分析。在该节课中,所有学生均表现出对实验的浓厚兴趣,并能按时完成实验。
(2)转座的PCR检测部分2次课,第一次课的实验内容为提取水稻叶片DNA;第二次课的实验内容为PCR扩增及电泳,拍照记录带型,要求学生课后对实验结果进行统计分析。该部分实验内容相对复杂,会有个别学生需要通过重复实验才能完成。
(3)实验结果讨论与教学延伸部分1次课。首先组织学生汇报实验结果,了解学生对科学研究的思路、方法及结果分析的掌握情况。其次,教师结合理论知识讲解转座子对其他基因表达调控及其在基因组进化和基因功能研究方面的重要应用价值。并以突变体为例,与学生探讨D插入突变体的基因克隆技术和功能分析方法,拓展学生遗传知识的深度与宽度。通过对实验结果的讨论不仅有助于提升学生实验总结能力,更能激发学生进一步探索的兴趣,课后一般会有超过半数的同学主动联系老师进行研究性实验或创新训练项目拓展。
3.2 引导学生进行研究性实验
针对部分对该突变材料和基因克隆感兴趣的同学,引导他们利用开放实验平台,自行开展材料的基因克隆研究。教学方法为:
首先老师与学生通过课堂讨论和文献资料查阅了解插入突变体材料的基因克隆方法,与基因的图位克隆方法相比较,并设计实验方案。图位克隆又称定位克隆,用该方法分离基因的实质是寻找与目的基因紧密连锁的分子标记,无需预先知道基因的DNA顺序,但因需要构建遗传分离群体而耗时较长。就插入突变体而言,由于插入序列是已知的,可以作为插入位点的分子标签采用其他途径克隆目的基因。在此基础上,要求学生进一步查阅资料、阅读文献拿出插入突变体基因克隆可供选择的多种方案(例如反向PCR和TAIL-PCR等),通过比较分析,选用其中一种方法开展研究。在此过程中,教师需及时跟进、解疑答惑,并做好引导。教学实践表明,几乎所有学生均会优先考虑选用TAIL-PCR方法进行该突变体的基因克隆。
确定采用TAIL-PCR方法后,师生共同讨论、制定并修改完善实验方案,根据实验方案开展基因克隆研究,实验过程中教师实时跟进指导。虽然TAIL-PCR扩增核酸的基本原理与普通PCR无异,但是需要研究者对基本的PCR扩增原理和影响因素有深刻的理解和把握。TAIL-PCR需3轮扩增,前一轮产物稀释后用作后一轮扩增的模板,而且每轮的扩增尤其第一轮PCR扩增程序比普通PCR复杂;同时TAIL-PCR引物又有退火温度较高的嵌套式特异性引物和退火温度较低的兼并引物之别(图1)。这些都是有别于普通PCR的地方,也是该研究型实验教学的重点和难点。
TAIL-PCR 扩增3′旁邻序列如图2C所示,特异扩增的第三轮PCR产物可直接用于测序。获得测序结果后,可指导学生利用公共核酸数据库(National Center for Biotechnology Information)进行初步的生物信息学分析,例如BLAST比对等便可获取目的基因的基本信息。本例的目的基因位于水稻第11染色体,属于类伸展蛋白基因家族。至此,插入突变体基因分离最关键的一步已完成。但是在实际教学中发现大多数学生需要重复多次才能获得理想扩增,在此过程中老师耐心指导尤为重要。实验最后,老师可结合TAIL-PCR电泳结果(如图2C)提出如何判断TAIL-PCR产物是否为特异扩增产物(由于特异引物是嵌套式的,第三轮产物比第二轮产物要小,可据此判别是否为特异扩增产物),分析研究结果及问题,撰写研究论文及研究报告。当然,后续有关基因功能的研究还有很多方面可以引导学生作进一步的思考。
4 教学中的思考与探索
该综合性实验融合了种子科学、经典遗传学的孟德尔遗传规律、基因组学的转座因子和统计学方差分析等理论知识内容,结合了表型观测、PCR检测技术、TAIL-PCR技术、基因克隆技术和方差分析法等实验内容。该综合性实验运用“综合性实验–研究性实验”的递进式教学体系,融合了“问题引导式—传授式—分组合作式—讨论分析式”混合教学法[14,15],以培养学生创新思维和方法为教学目标,让学生学会如何在科学研究中发现问题、分析问题、提出问题、研究问题,最后得出科学研究结论;让学生明白知识之间的联系性和整体性,进而实现知识融会贯通;有助于学生在团队协作中提升解决问题和创新实践的能力,这也是科研成果转化为本科实验教学的一个有益探索。
但在教学实践中,要使本课程达到最佳教学效果,还需要思考以下两点:首先,本实验所使用的材料来源于科学研究,与其相关的理论背景知识涉及不完全显性遗传与基因突变中的转座因子,然而这部分理论知识在遗传学理论教学中不是重点讲解内容,容易被学生忽视。因此在实验教学的过程中,需要将这部分理论知识整合进去,不仅让学生通过实验可以理解,更希望能引导学生将显性与不完全显性遗传、跳跃基因与转座子、转座因子的分类及其与基因突变进行对比分析,并将它们融会贯通;其次,综合性实验和研究性实验在教学中是层层递进式关系,在培养学生综合创新能力上发挥着不可替代的作用。综合性实验是学生进入科学研究领域前的传送带,旨在激发学生的研究兴趣,并使其掌握基本的研究方法和实验技能。研究性实验是学生进入科学研究领域的缩影,旨在让学生经历科学研究的过程,了解科学研究方法,进而培养学生创新思维及解决问题的能力。在综合性实验设计与教学过程中,如何加强综合性实验与研究性实验的关联性与整体性,激发学生研究的兴趣,引导学生进入研究性实验,一直是我们教学过程中需要不断思考与创新的地方。
[1] Chen DF, Lu DR, Zhang FX, Zhang GF. The development of genetics teaching in China in the last four decades and its future prospect., 2018, 40(10): 916–932.陈德富, 卢大儒, 张飞雄, 张根发. 中国遗传学教学40年发展及展望. 遗传, 2018, 40(10): 916–932.
[2] Li N, Guo HB, Li YJ, Lao MM. Design and practice of comprehensive experiment for cytogenetics., 2019, 38(9): 200–203, 227.李楠, 郭海滨, 李亚娟, 劳媚媚. 细胞遗传学综合性实验的设计与实践. 实验室研究与探索, 2019, 38(09): 200–203, 227.
[3] Liu ZQ, ZhaoYX, Fu XL, Li N. Design and exploration of genetic experiments for non-mendelian segregation., 2019, 41(3): 262–270.刘自强, 赵苑秀, 傅雪琳, 李楠. 偏孟德尔分离的遗传学实验设计与探讨. 遗传, 2019, 41(3): 262–270.
[4] Qi P, Lin YS, Song XJ, Shen JB, Huang W, Shan JX, Zhu MZ, Jiang LW, Gao JP, Lin HX. The novel quantitative trait locus GL3.1 controls rice grain size and yield by regulating Cyclin-T1;3., 2012, 22(12): 1666– 1680.
[5] Segami S, Kono I, Ando T, Yano M, Kitano H, Miura K, Iwasaki Y. Small and round seed 5 gene encodes alpha-tubulin regulating seed cell elongation in rice., 2012, 5(1): 4.
[6] Cho SH, Yoo SC, Zhang H, Pandeya D, Koh HJ, Hwang JY, Kim GT, Paek NC. The rice narrow leaf2 and narrow leaf3 loci encode WUSCHEL-related homeobox 3A (OsWOX3A) and function in leaf, spikelet, tiller and lateral root development., 2013, 198(4): 1071–1084.
[7] Jin J, Shi JL, Liu B, Liu YC, Huang Y, Yu Y, Dong AW. MORF-RELATED GENE702, a reader protein of trimethylated histone H3 lysine 4 and histone H3 lysine 36, Is involved in brassinosteroid-regulated growth and flowering time control in rice., 2015, 168(4): 1275–1285.
[8] Liu Y, Whittier RF. Thermal asymmetric interlaced PCR: automatable amplification and sequencing of insert end fragments from P1 and YAC clones for chromosome walking., 1995, 25(3): 674–681.
[9] Liu F, Zhang XQ, Zhang ZM, Chen ZG, Zhu HT, Wang J, Zhang JL, Zhang GQ. Transpositional behaviour of theelement in the/system in rice., 2007, 52(20): 2789–2796.
[10] Liu XD, Li YJ. General Genetics Comprehensive Experiment. Beijing: China Agriculture Press, 2011.刘向东, 李亚娟. 普通遗传学综合性实验. 北京: 中国农业出版社, 2011.
[11] Jeon JS, An G. Gene tagging in rice: a high throughput system for functional genomics., 2001, 161(2): 211–219.
[12] Park SH, Kim CM, Je BI, Park SH, Park SJ, Piao HL, Xuan YH, Choe MS, Satoh K, Kikuchi S, Lee KH, Cha YS, Ahn BO, Ji HS, Yun DW, Lee MC, Suh SC, Eun MY, Han CD. A-insertion mutant of OSH6 (Homeobox 6) exhibits outgrowth of vestigial leaf-like structures, bracts, in rice., 2007, 227(1): 1–12.
[13] Zhu QH, Dennis ES, Upadhyaya NM. Compact shoot and leafy head 1, a mutation affects leaf initiation and developmental transition in rice (L.)., 2007, 26(4): 421–427.
[14] Qiang GR, Wang H, Yang ZP, Wang HB, Fang WJ. Research and practice on teaching methods of college organic chemistry experiment., 2017, 36(03): 200–202, 211.强根荣, 王红, 杨振平, 王海滨, 方文军. 大学有机化学实验教学方法研究与实践. 实验室研究与探索, 2017, 36(03): 200–202, 211.
[15] Zu Q, Liu JB, Sun YM, Hu K. Reform of experimental teaching method in colleges and universities., 2015, 13(03): 88–90.祖强, 刘加彬, 孙岳明, 胡凯. 大学本科实验教学方法改革初析. 实验科学与技术, 2015, 13(03): 88–90.
Design and exploration of genetic comprehensive experiments based oninsertion mutants
Nan Li1, Yajuan Li1, Haibin Guo1, Xiangqian Zhang2
The cultivation of innovative abilities has become an important guide for higher education in China. Strengthening the integrated knowledge to design experiments is an effective way to improve undergraduate students’ innovative abilities. Herein we designed a comprehensive experiment for molecular genetics by utilizing a riceinsertion mutant identified previously in our research project. In the comprehensive experiment, we adopt the method of scientific research as the main line of teaching and take the interesting phenotype of the rice mutant as the breakthrough point to reform and innovate genetics laboratory teaching. On the basis of this, we combined the progressive teaching method and guided the students to learn the TAIL-PCR skill and conduct an innovative experiment through expanding their knowledge. The comprehensive experiment will deepen students’ understandings of the relationship between genotypes and phenotypes, help them master the effective way of thinking and technologies for scientific research to further improve their ability of the integrated application capability of theory and practices.
comprehensive experiment; experimental teaching; gene mutation
2019-09-26;
2019-11-01
国家自然科学基金面上项目(编号:30900884,31671594)和华南农业大学2017年教学改革与研究项目(编号:JG17093)资助[Supported by the National Natural Science Foundation of China (Nos. 30900884, 31671594) and Teaching Reform and Research Projects of SCAU in 2017 (No. JG17093)]
李楠,硕士,实验师,研究方向:遗传学实验教学与管理。E-mail: 523155900@qq.com
张向前,博士,副教授,研究方向:植物分子育种。E-mail: aacrav@163.com
10.16288/j.yczz.19-149
2019/11/7 14:34:00
URI: http://kns.cnki.net/kcms/detail/11.1913.R.20191107.1151.002.html
(责任编委: 陈德富)
