鹿茸间充质干细胞条件培养基对乳腺癌细胞系4T1的作用
2019-12-20孙胜楠孙铭泽杨春丁尚红张琦郭佳刘莹杨敏彭英华
孙胜楠,孙铭泽,杨春,丁尚红,张琦,郭佳,刘莹,杨敏,彭英华※
(1.中国农业科学院特产研究所,长春 130112;2.延边大学,吉林 延吉 133002)
间充质干细胞(Mesenchymal stem cells,MSCs)因具有多项分化、归巢、分泌多种细胞生长因子、分泌外泌体的能力,逐渐作为新的治疗手段而被广泛关注[1-2]。MSCs可直接通过细胞间接触或分泌旁分泌因子与癌细胞相互作用,这种干细胞疗法为癌症治疗提供了新的思路。研究表明,MSCs 所分泌的因子是其发挥作用的重要部分,MSCs 条件培养基内含有MSCs 分泌的各种因子,并被证实对癌症有治疗作用。MSCs 的条件培养基可下调肝癌和乳腺癌细胞NF-B 的表达,抑制肿瘤细胞的增殖[3]。骨髓来源MSCs 释放的可溶性因子可显著抑制黑色素瘤、肺癌和胶质瘤等的生长和转移。脐带血来源的MSCs 可通过上调肿瘤坏死因子相关凋亡诱导配体(TRAIL)的分泌,抑制多形性胶质细胞瘤的增殖[4]。将MSCs 与人肝癌细胞株共培养,MSCs 能够抑制肝癌细胞株增殖、促进其凋亡、下调Wnt 信号通路相关基因(Bcl-2、c-Myc、PCNA)表达。使用MSCs的条件培养基处理人肝癌细胞株时可获得相同的实验结果,这一研究证实,MSCs的旁分泌效应参与了肿瘤的调节[5]。尽管间充质干细胞具有多能性、趋向性等,但因其来源有限、在体外具有细胞活性较低等特点,限制了其在疾病治疗中的应用。
鹿茸是唯一能够每年完全再生的哺乳动物组织。研究表明,鹿茸的再生是基于干细胞的再生过程。随着鹿茸的生长,骨膜组织向间充质组织转化,其中的间充质细胞仍然具有干细胞的特征[6-7]。相比之下,鹿茸组织来源的间充质干细胞除了具有人源间充质干细胞的增殖快、分化能力强等特点之外,还具有易于获得、来源广泛等优点[8]。为了规避免疫原性,深入研究鹿茸间充质干细胞旁的分泌作用,本试验探讨了富含旁分泌因子的鹿茸间充质干细胞条件培养基对小鼠乳腺癌的作用,为其抗肿瘤应用提供实验依据。
1 材料与方法
1.1 主要试剂与仪器
胎牛血清(FBS)、青霉素-链霉素双抗、0.25%胰酶、磷酸缓冲盐溶液(Phosphate buffer saline,PBS)、胶原酶I(美国Gibco 公司);DMEM 培养基、DMEM/F12培养基(美国Corning 公司);总蛋白提取试剂盒、蛋白浓度测定试剂盒、ECL 发光试剂盒(上海碧云天生物技术有限公司);Bcl-xl、cleaved-caspase 3、caspase 3、-actin一抗及二抗抗兔(美国Cell Signaling Technology公司);CD44、CD90、CD105、CD31 一抗(美国 Biolegend 公司);MTT 细胞增殖及细胞毒性试剂盒(北京普洛麦格生物技术有限公司);CO2 恒温培养箱(美国Thermo公司);蛋白电泳仪、转膜仪(美国Bio-Rad 公司)。
1.2 细胞的分离与培养
1.2.1 鹿茸间充质干细胞的分离、培养 参考Li 等[9]的分离培养方法,取生长初期的梅花鹿鹿茸,消毒,切取5 cm 长的鹿茸尖部,挤血。从鹿茸中部纵向切开,75%乙醇清洗 3 遍,每次2 min,PBS 洗3 遍。剥离真皮层,在解剖显微镜下定位并切取鹿茸间质层,用含有双抗的PBS 清洗间质层组织3 次,将其切碎成沫状,PBS 清洗后,用0.1%胶原酶I 于37℃的CO2 培养箱中孵育2 h。消化后离心(1000 r/min 8 min)除去消化液,用细胞培养液清洗2 次后转移至培养瓶中,放入37℃的CO2 培养箱中进行培养,每隔2 d换液,细胞融合度达80%冻存即可。
1.2.2 小鼠乳腺癌细胞系4T1 的培养 使用含10%胎牛血清以及含100 U/mL 青霉素和100g/mL 链霉素的DMEM 培养基,于 37 ℃、5%CO2 培养箱内培养小鼠乳腺癌细胞4T1。
1.3 鹿茸间充质干细胞条件培养基的制备
用含10%FBS 的DMEM/F12 培养基培养鹿茸间充质干细胞,待细胞长至融合度为80%以上时,将培养液换为不含FBS 的DMEM/F12 培养基继续培养24 h后,收集培养上清,1 000 r/min 离心10 min,收取上清液,过0.22m 无菌滤器,80℃冰箱保存备用。
1.4 细胞处理
分别用0、50%、75%、100%的条件培养基处理4T1细胞24 h。
1.5 流式细胞术鉴定
取对数生长期的鹿茸间充质细胞,接种到6 孔板中,细胞融合度达到80%后,加入一抗(CD44、CD90、CD105、CD31),室温避光条件下孵育30 min,流式细胞仪检测阳性细胞率。
1.6 细胞增殖检测
取对数生长期的4T1 细胞,均匀接种于96 孔板中,每孔1 104个,加入终浓度分别为0、50%、75%、100%的鹿茸间充质干细胞条件培养基,设置3次重复,增殖检测严格按MTT 说明书操作。
1.7 Western Blot检测
使用预冷的PBS清洗处理后的细胞,蛋白裂解液裂解30 min。5%的浓缩胶电泳0.5 h 后,12%分离胶电泳1 h,使用湿转的方法将蛋白转膜至PVDF 膜上,5%的脱脂奶粉室温封闭1 h,一抗4℃过夜孵育,每隔5 min 使用 TBST 清洗 1 次,共洗 5 次,二抗室温孵育1 h,TBST 洗涤 5 次,显影。
1.8 统计学处理
使用图像分析软件Image J,对Western blot 图像条带进行灰度分析量化表达的蛋白。使用 GraphPad Prism 5.0 软件对数据分析,并用two-tailed unpaired ttest 及Tukey 检验法进行多重统计学分析。
2 结果
2.1 鹿茸间充质干细胞的分离
使用倒置光学显微镜,观察培养不同阶段的细胞形态,消化1 d 后的组织块均匀贴壁于培养瓶底部,3~4 d可见组织块周围爬出少量细胞,5~7 d贴壁细胞形成群落,组织块四周细胞呈放射状向周边生长。连续培养14 d 后,细胞融合可达80%。经传代培养,细胞成典型的单层长梭样生长。传代培养12 h 后即可完全贴壁,2 d左右细胞即可长满整个培养瓶底面,并可继续传代扩增。P5 和 P7 代鹿茸间充质干细胞呈鱼群样生长,且细胞生长状态良好。结果见图1。
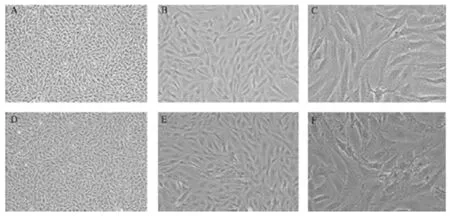
图1 鹿茸间充质干细胞形态Figure 1 Morphology of antler mesenchymal stem cells
2.2 鹿茸间充质干细胞的鉴定
为确定所分离的间充质细胞为实验所需的鹿茸间充质干细胞,对所分离的细胞进行干细胞表面抗原鉴定。经流式细胞术检测,鹿茸间充质细胞高表达间充质干细胞标志蛋白为CD44(83.23%)、CD90(83.08%)、CD105(97.64%),不表达白细胞特异性标志蛋白为CD31(1.59%)。结果表明,实验过程中所获得的细胞符合间充质干细胞的鉴定标准(图2)。
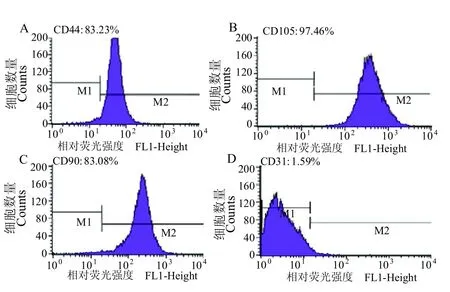
图2 流式细胞术检测鹿茸间充质干细胞表达表型Figure 2 Flow cytometry was used to detect the expression phenotype of antler mesenchymal stem cells
2.3 鹿茸间充质干细胞条件培养基对4T1细胞增殖情况的影响
取对数生长期的4T1 小鼠乳腺癌细胞均匀接种于96 孔板中,分别使用0、50%、75%、100%的鹿茸间充质干细胞条件培养基处理4T1 小鼠乳腺癌细胞24h。MTT检测结果显示,条件培养基对4T1 细胞活性具有抑制作用,并且随着条件培养基的浓度增加抑制作用增强。0、50%、75%、100%的鹿茸间充质干细胞条件培养基相对应的细胞存活率为100%、81.1%、73.7%、59.6%。结果见图3、表1。

图3 MTT 法检测鹿茸间充质干细胞条件培养基对4T1 细胞增殖的影响Figure 3 MTT was used to detect the effect of antler mesenchymal stem cells conditioned medium on 4T1 cell proliferation

表1 鹿茸间充质干细胞条件培养基对4T1 细胞增殖的影响Table 1 The effect of antler mesenchymal stem cells conditioned medium on 4T1 cell proliferation
2.4 鹿茸间充质干细胞条件培养基对4T1细胞凋亡信号通路的影响
不同浓度的鹿茸间充质干细胞条件培养基处理4T1 小鼠乳腺癌细胞24 h 后,Western blot 方法检测凋亡相关蛋白 Bcl-xl、Cleaved-caspase 3、Caspase 3 的表达水平。结果显示,随着条件培养基处理浓度的增加,4T1 细胞中Bcl-xl 蛋白表达水平呈浓度梯度下降,而Cleaved-caspase 3 蛋白表达水平呈浓度梯度升高。这一结果提示,4T1 小鼠乳腺癌细胞的增殖能力下降,是通过影响细胞中Bcl-xl、Cleaved-caspase 3 凋亡通路的 蛋白表达实现的。结果见图4。
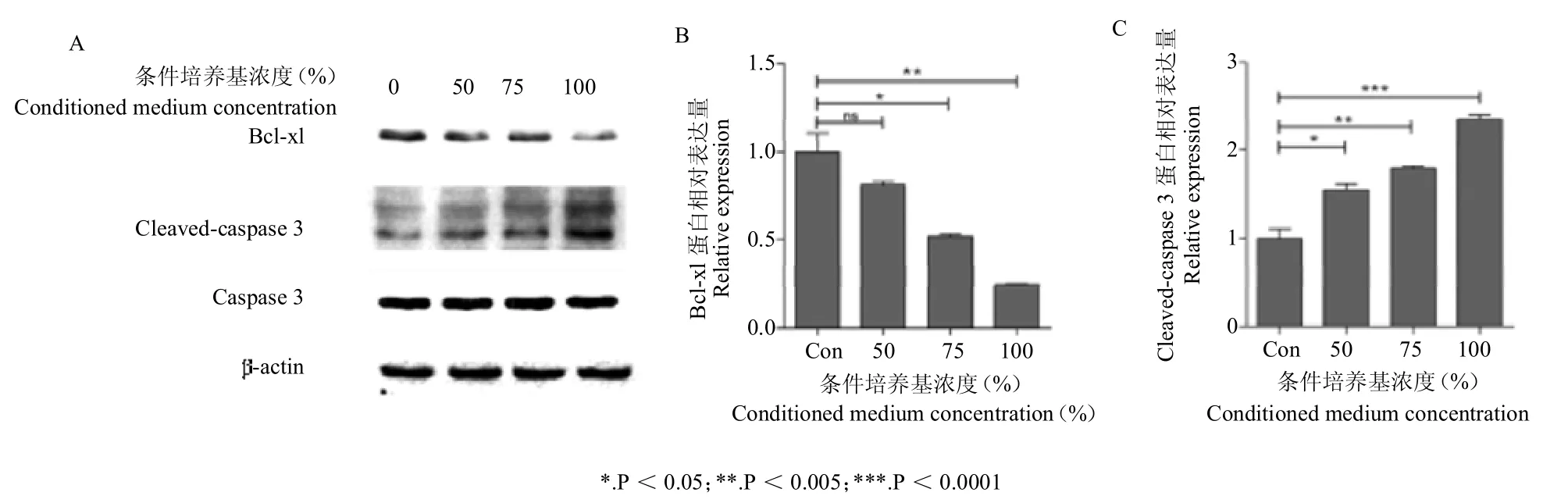
图4 Western blot 检测 Bcl-xl、cleaved-caspase 3、caspase 3 蛋白表达水平Figure 4 Western blot was used to detect the expression of Bcl-xl,cleaved-caspase 3 and caspase 3 in 4T1 cells
3 讨论
干细胞在体外可诱导分化为特定的细胞和组织。近年来,随着对其研究和应用的不断深入,间充质干细胞在肿瘤治疗中的潜在价值也逐渐为人们所认识。有研究发现,人脐带间充质干细胞可以抑制乳腺癌干细胞增殖,且抑制作用呈现出剂量依赖性,其原因可能是人脐带间充质干细胞具有抑制细胞周期、诱导肿瘤细胞凋亡的作用,并能够抑制PI3K 和Akt 蛋白激酶的活性,从而显著抑制乳腺癌干细胞的增殖[10]。然而,人脐带间充质干细胞来源有限,在很大程度上限制了其在疾病治疗中的应用,相比之下,鹿茸组织来源的间充质干细胞来源广泛,其旁分泌作用更应被考虑应用于癌症的治疗。本研究分离获得了鹿茸组织来源的间充质细胞,经流式细胞仪检测,细胞不表达白细胞表面特异性标志CD31,但可以表达CD44、CD90、CD105干细胞标志蛋白。结果表明,实验过程中所获得的细胞为鹿茸间充质干细胞(图1~2)。
目前,人们对于干细胞治疗疾病的作用机制尚未完全阐明,存在两种较为认可的机制:分化和旁分泌机制[11]。随着研究的深入,人们发现旁分泌机制起重要的调控作用,即间充质干细胞可能是通过分泌生物活性因子,对肿瘤的细胞信号通路产生一定的影响。干细胞的培养基中富含旁分泌的生物活性因子,通过条件培养基作用于癌细胞,可阐明旁分泌的作用效果。本研究中,鹿茸间充质干细胞条件培养基处理4T1 小鼠乳腺癌细胞24h后,MTT检测结果显示,条件培养基对4T1细胞的活性具有抑制作用,并随着条件培养基的浓度增加抑制作用逐渐增强(图3)。该结果证明了小鼠乳腺癌4T1 细胞的抑制依赖于旁分泌的活性因子。
在多细胞生物中,细胞凋亡是发展和维持组织稳态的一个重要的进化保守机制,但在癌细胞中其机制会受到干扰,比如原癌基因的激活、抑癌基因的失活等,导致其凋亡受到显著抑制,从而凋亡率极低[12]。细胞凋亡能导致细胞发生某些变化,如活化Caspases、线粒体去极化和DNA 断裂等[13]。Bcl-2 基因家族成员编码具有促进或抑制细胞凋亡功能的蛋白,如抗凋亡蛋白(Bcl-xl)能够通过多种刺激抑制细胞凋亡的发生。Caspases是一个成员较多的蛋白酶家族,目前发现,人体内的Caspases蛋白家族成员至少有11 种,其中Caspase 3 在细胞凋亡过程中发挥凋亡执行因子的作用。Caspase 3 在正常情况下以酶原的形式存在,当细胞发生凋亡时,Caspase 3 被活化成为Cleaved-caspase 3,发挥促进细胞凋亡的作用,并标志着细胞凋亡进入不可逆转的阶段[14-15]。在本研究中,随着条件培养基浓度升高,4T1 细胞中Cleaved-caspase 3 表达水平呈上升趋势,而Bcl-xl 蛋白表达水平呈下降趋势。这一实验结果表明,鹿茸间充质干细胞条件培养基抑制乳腺癌4T1 细胞的增殖,其机制与细胞凋亡通路有关(图4)。
综上所述,在体外实验中,鹿茸间充质干细胞条件培养基可抑制乳腺癌细胞4T1 的增殖,其机制与Cleaved-caspase 3 表达水平升高和Bcl-xl 表达水平下降有关。本研究为鹿茸间充质干细胞旁分泌效应对肿瘤的作用机制研究提供了一定的实验依据,为进一步阐明间充质干细胞旁分泌效应在肿瘤治疗中的作用提供了基础。但由于肿瘤的表现和机制以及鹿茸间充质干细胞条件培养基对肿瘤的作用十分复杂,其具体机制仍需进一步研究。
