浙江西天目山古树树干健康状况及其随海拔梯度变化规律
2019-12-20张凤麟张讴凯张昭臣
张凤麟, 王 昕, 张讴凯, 张昭臣∗, 张 健
(1.华东师范大学生态与环境科学学院, 浙江天童森林生态系统国家野外科学观测研究站, 上海200241; 2.上海污染控制与生态安全研究院, 上海200092)
古树是自然和城市环境中的重要组成部分,为各种动植物的生长和繁殖提供了重要生境(Lindenmayeret al,2017;Zemanováet al,2017),为人类社会提供生态系统服务(Körner,2017),同时具有较高的美学和文化价值。 由于古树树龄往往较高,自身抵抗外界干扰及进行自我修复的能力较弱,大部分古树都存在一定的健康问题。 鉴于全球范围内古树和大树所面临的气候变化和人类干扰等的严重威胁,著名保护生物学家Lindenmayer、Laurance 和著名森林生态学家Franklin 曾于2012 年在Science 杂志联合发文,呼吁大家重视这一问题的严重性,并尽快采取措施来保护濒临死亡的古树和大树(Lindenmayeret al,2012)。 基于CNKI 数据库,在1990—2018 年期间,古树健康相关的中文文献仅70 篇,且主要为近6 年发表,其研究范围主要集中在较发达国家的城市、保护区等,研究对象主要在城市区域,而古树资源丰富的山区(Liuet al,2019;Myerset al,2000;Sandelet al,2013)研究较少。 山区作为生物多样性热点区域,地形复杂,存在一定的海拔梯度。 沿海拔梯度,环境因子(温度、降水、大气成分以及土壤成分等)呈现极大的变化(Gaston,2000),古树的分布随海拔梯度也可能会呈现出有规律的分布格局。 同时,沿海拔梯度分布的生物受到不同程度的人类干扰(Sandelet al,2013),一般来说,低海拔地区受到较强程度的人类干扰,而高海拔地区干扰较少。 这些人类干扰程度的差异可能也会对古树的生长和繁殖等产生重要影响。 因此,分析古树健康状况沿海拔梯度的分布规律及其限制因素有助于古树管理和保护策略的制定与实施等。
浙江天目山国家级自然保护区植被类型多样、生物资源丰富,在保护区内从低海拔到高海拔分布着数量巨大的古树资源。 根据楼涛等(2004)调查结果显示,该区域古树种类繁多,数量丰富,常见的古树种类有银杏(Ginkgo biloba)、柳杉(Cryptomeria fortunei)、金钱松(Pseudolarix amabilis)等。 现存古树约5 511 株,隶属43 科,73 属,共计100 种,其树龄主要集中在100~300 年,500 年以上的一级古树数量较少,大多为柳杉、金钱松和银杏等古老树种(楼涛等, 2004)。 同时,天目山古树名木在科属种间的数量上存在悬殊差异,存在1 000 株以上个体的优势科为杉科(Taxodiaceae)和松科(Pinaceae);优势种为柳杉,其个体数量达2 000 余株,然而个体数小于5 株的物种有54 个。 尽管针对天目山的古树已经开展了一些研究工作,但尚未对天目山古树健康状况沿海拔梯度的变化规律及其影响因素进行系统性和定量化的研究工作。
本研究选择使用PICUS-3 应力波树木断层图像诊断仪对古树的树干腐烂程度调查结果作为古树健康状况研究的切入点,对浙江省天目山国家级自然保护区内不同海拔梯度上不同树种的古树健康状况开展调查,分析不同健康状况下的古树分布情况,讨论影响古树健康状况的主要因素,以期为西天目山古树的保护提供科学依据。
1 研究区域自然地理状况
天目山国家级自然保护区(北纬30°18′30″~30°21′37″,东经119°24′11″~119°27′11″),位于浙江省西北部的西天目山(图1)。 1986 年,西天目山经国务院批准,成为全国首批20 个国家级自然保护区之一。 1996 年,被联合国教科文组织批准接纳为国际人与生物圈保护区(MAB)网络成员。 目前保护区面积为4 284 hm2,保护区内植被保存完好,生物资源丰富(天目山自然保护区管理局,1992)。 1.5 亿年前的火山活动使天目山形成了如今的丘陵地貌,西天目山主峰仙人顶,最高海拔1 506 m,最低海拔约300 m。 保护区内气候较为温和,地处中亚热带向北亚热带过渡区域,受海洋季风影响,四季分明。 年均温度8.8~14.8 ℃,最冷月平均温度为-2.6~3.4 ℃,极值最低气温-13.1~-20.2℃,最热月平均温度为19.1 ~28.1 ℃,极值最高气温29.1~38.2 ℃(商侃侃,2011)。 年降水量为1 390 ~1 870 mm,形成浙江西北部的降水中心(夏冰等,1992);无霜期209~235 d;相对湿度76%~81%。 该地区光照充足,年太阳辐射量为3 279~4 460 MJ·m-2,春秋季较短,冬夏季较长(楼涛等,2004)。 作为长江和钱塘江支流的发源地和分水岭,西天目山水文基础良好,沟谷中地表水径流量受降水控制,变化幅度较大(商侃侃,2011)。 土壤厚度约50 cm,海拔600 m 以下为红壤带,600 ~1 200 m 为黄壤带,1 200 m以上为棕黄壤带,土壤腐殖质较厚,具有较高的肥力(蒋文伟,2001)。 总体而言,西天目山优良的环境为古树的生长提供了良好的条件。
图1 浙江天目山国家级自然保护区位置图Figure 1 Location of Tianmu Mountain, Zhejiang Province
2 研究内容与方法
基于上文中各种问题,本文拟从以下两个方面展开研究:(1) 浙江西天目山古树健康状况的定量评估:根据测量所得的古树树干断层图像对所测古树树干胸高断层上的腐烂及空洞情况做出分析,综合得出古树的整体健康状况;(2) 古树健康状况的海拔梯度差异:通过对不同海拔段上的古树健康状况进行比较,分析古树健康状况沿海拔梯度的变化规律;并通过不同海拔段上的气候数据与古树健康状况之间的相关性分析验证海拔梯度与古树健康状况之间的联系,探究海拔变化导致的环境变化对古树健康状况的影响。
2.1 海拔梯度划分
根据天目山现有的古树资源调查情况(楼涛等,2004),以及西天目山的植被分布情况(夏爱梅,2004),本研究对天目山的海拔进行分段,分别为300 ~400、400 ~500、500 ~600、600 ~700、700~800、800~900、900~1 000、1 000~1 100 和1 100~1 200 m。 在不同的海拔梯度上,视古树分布情况选择不同物种的挂牌古树,尽可能保证每个海拔段上选取的古树均匀分布在该海拔段内。
2.2 古树健康检测方法
为检测西天目山古树的健康状况,本研究选用德国ARUGS 公司制造的PICUS-3 应力波树木断层图像诊断仪来进行数据采集。 该诊断仪用于测量每株古树树干距离地面130 cm 处的树干断层腐烂情况,测量时在树干朝北方向的树皮上钉入第一颗钉子,测量树干周长,按一定的长度间隔(≥20 cm)选择测量点位置,逆时针依次钉入其余钉子,挂号牌。 选取3 颗钉子作为基准点,分别测量记录每个钉子到基准点间的直线距离,输入到PICUS Q74 分析软件中,软件自动生成可视化的树干的测量层的外形几何图。 记录树种、海拔位置等相关信息。 将PICUS-3 应力波树木断层图像诊断仪主机利用绑带固定在树干上6 ~7 号钉子中间的位置上,将传感器分别连接在对应号牌的钉子上,将电子锤和电脑连接至PICUS 主机,准备工作完成。测量时摘下相应钉子上的传感器,将电子锤上的应力波接收器放置在该钉子上,敲击接收器,其余钉子上的传感器接收到应力波,将数据传输至主机。 沿逆时针方向依次敲击所有钉子,PICUS 主机计算出树干断层的层析图像,传至电脑端软件,保存数据。 测量结束后摘下钉在树皮上的钉子,完成本次测量工作。
2.3 数据处理
对于收集到的树木断层图像数据,使用PICUS Q74 软件获得直观的图像数据,再利用R语言v.3.5.1(http:∥www.R-project.org/)对源文件进行数据提取,获得不同颜色在图像中的面积占比数据。
由于获得是百分比数据,我们选择β回归分析(Ospinaet al,2012)进行数据分析。β回归模型常被用于模拟因变量属于β分布的分析。β分布可以方便地根据因变量的均值和精度参数进行参数化,能够较为直观的表现出不同因素之间的相关性,并拟合线性结果。 本研究所获得数据为树木断层诊断图像中不同颜色所占总面积的比值,分布在(0,1)区间内,属于β分布,可以使用β回归模型分析断层中的腐烂水平,判断古树健康水平。 在R 语言中利用betareg包(Cribari-Netoet al,2010)完成。
各采样点的气候数据从ClimateAP 软件中提取,ClimateAP 是由加拿大英属哥伦比亚大学的Tongli Wang 和阿尔伯塔大学的Andreas Hamann 等采用插值技术和高程修正的方法制作的局域尺度上高精度的气候数据集(Wanget al,2006)。 通过输入每株被测量古树的地理位置信息获得其对应地理位置上1981—2010 年间气候数据(张凤麟等,2018),考虑到各因子之间的共线性(multilinearity)可能对数据分析的影响,我们对变量进行了筛选,最终选取了年均温、年降水量、热湿比和无霜期这4 个环境因子用于本研究。
3 结果与分析
3.1 古树健康状况的种间差异
本次调查古树共计137 株,通过对所有古树树干断层图像的分析(图2),在胸高断面上未出现腐烂或空洞情况的古树仅3 株,占总量的2.1%;出现了一定程度的腐烂但树干中尚未出现明显空洞的古树占总量的35.8%;树干中已经出现明显空洞的古树占总量的62.1%,其中空洞占树干面积5%以上的有32 株,占总量的23.4%。

图2 不同古树的PICUS 诊断画像Figure 2 PICUS Sonic Tomograms of different ancient trees
通过对所有古树树干断层图像中有明显腐烂和空洞情况的面积占比进行分析(图3),结果显示,所测量的古树树干断层腐烂及空洞面积所占比例的均值为18.89%,各个物种的情况相较于均值而言有一定程度的差异。 调查结果中树干腐烂及空洞面积占比较高的树种有金钱松、天目木姜子(Litsea auriculata)、天目铁木(Ostrya rehderiana)和柳杉,其树干内部腐烂及空洞面积占比高于均值,分别为:金钱松38.89%、天目木姜子29.29%、天目铁木20.68%和柳杉19.34%,其中本次所调查测量的6 株金钱松其树干中均出现明显空洞。 树干腐烂及空洞面积占比较低的树种有枫香、浙江楠(Phoebe chekiangensis)、黄山松(Pinus taiwanensis)、麻栎(Quercus acutissima)、银杏、榧树(Torreya grandis)和水杉(Metasequoia glyptostroboides),其树干内部腐烂及空洞面积占比均低于平均值,分别为:枫香13.50%、浙江楠14.35%、黄山松10.54%、麻栎6.05%、银杏5.64%、榧树0.96%以及水杉0。
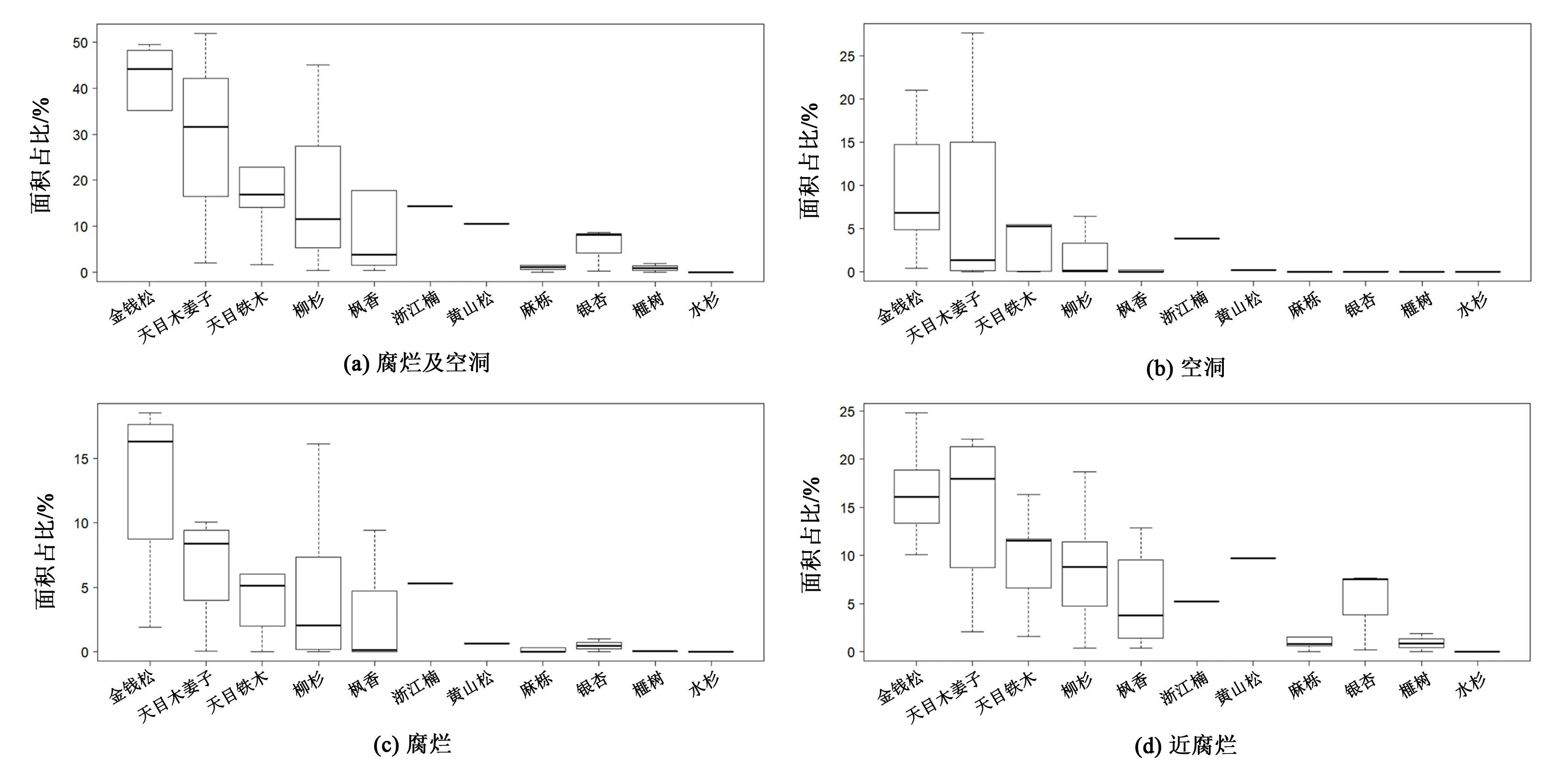
图3 古树树干腐烂及空洞面积占比种间差异Figure 3 The ratio of the tree trunk decay and hollow area of ancient trees among different species
3.2 各古树物种的健康状况在海拔梯度上的差异
通过对不同海拔下的各树木断层图像进行分析发现,小于400 m 海拔范围内的古树树干腐烂水平的均值为10.67%,400~500 m 海拔范围内古树树干腐烂水平的均值为8.75%,500 ~600 m 海拔范围内古树树干腐烂水平的均值为11.28%,600 ~700 m 海拔范围内古树树干腐烂水平的均值为16.54%,700~800 m 海拔范围内古树树干腐烂水平的均值为18.71%,800 ~900 m海拔范围内古树树干腐烂水平的均值为14.71%,900~1 000 m 海拔范围内古树树干腐烂水平的均值为19.10%,1 000~1 100 m 海拔范围内古树树干腐烂水平的均值为31.47%,1 100 ~1 200 m 海拔范围内古树树干腐烂水平的均值为30.06%。
根据以上结果,使用beta 回归对古树树干断层腐烂及空洞面积占比数据分析,结果显示,古树树干断层上的腐烂及空洞面积占比随海拔的不断上升有一定的上升趋势(P<0.0001) (图4)。

图4 古树树干腐烂及空洞面积占比与海拔间关系Figure 4 Relationship between ratio of tree trunk decay and hollow area and altitude
3.3 气候对各古树物种的健康状况的影响
温度、降水、热湿比和无霜期都是具有明显海拔梯度效应的环境因子,随海拔变化而存在一定程度的变化,结合气候数据(Wanget al,2012),使用beta 回归方法,对古树树干腐烂及空洞面积占比的变化趋势分析,结果如下:(1)天目山范围内,随着海拔的不断升高,温度逐渐降低,古树树干的腐烂及空洞面积占比下降[图5(a)];(2)降水量随海拔的升高而升高,古树树干的腐烂及空洞面积占比随降水量升高而增大[图5(b)];(3)热湿比随海拔梯度的升高而降低,古树树干腐烂面积随热湿比的升高而减小[图5(c)];(4)无霜期随海拔的升高而减短,海拔较高处无霜期相对较短,古树树干的腐烂及空洞面积占比相对较高[图5(d)]。 这些分析结果在一定程度上都验证了古树树干腐烂及空洞面积占比随海拔的上升不断增大的结论。
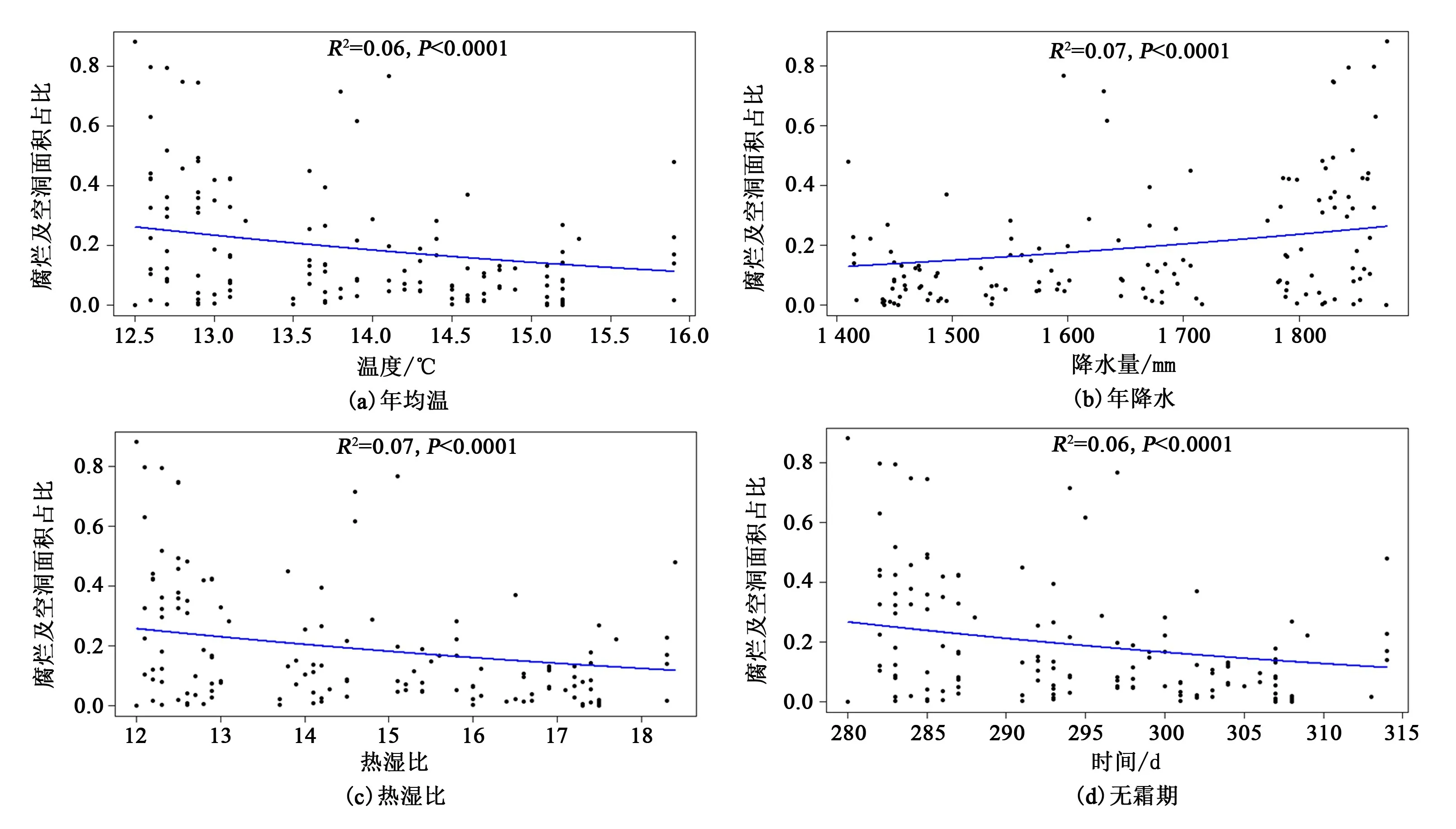
图5 古树树干腐烂及空洞面积占比与环境因子间关系Figure 5 Relationship between ratio of tree trunk decay and hollow area and environment factors
4 讨论
通过对天目山不同物种、不同海拔古树树干腐烂及空洞面积占比的分析,结果发现:不同物种间古树树干腐烂及空洞面积占比存在差异,同种个体间古树树干腐烂及空洞面积占比也存在差异;随海拔上升,古树树干腐烂及空洞面积占比存在上升趋势,即海拔与古树树干腐烂及空洞面积占比间存在联系。
(1) 相关研究表明,真菌入侵是造成树干内部空洞形成的部分原因(Arhipova,2012),物种间木材密度的不同影响了木材腐朽真菌侵蚀树干的能力(张丽燕等,2016),导致了树木间形成腐烂空洞情况的差异,而木材密度与古树树干内部空洞的面积存在负相关关系(Jacobsenet al,2007)。 在本研究中,腐烂及空洞面积占比较高的金钱松,其气干密度为0.4 ~0.5 g·cm3(成俊卿,1992),柳杉,其气干密度约为0.33 g·cm3(易咏梅等,2003)。 而腐烂及空洞面积占比较低的麻栎,其气干密度约为0.8 g·cm3(成俊卿,1992),榧树气干密度约为0.56 g·cm3(成俊卿,1992)。 不同物种其木材密度存在一定差异,且符合密度与腐烂及空洞面积的反比关系。因此,不同物种木材密度的差异对古树树干腐烂及空洞面积占比情况造成影响,进而导致古树树干腐烂及空洞面积占比的种间差异。
(2) 影响树木健康状况环境因子主要包括温度、降水、土壤性质、人为干扰强度等(刘瑜等,2013),这些因素在不同海拔间都存在一定差异。 本研究中,温度较低时,古树树干的腐烂及空洞面积占比较高。 该研究结果与海拔及古树树干腐烂面积占比间关系一致,低海拔地区温度较高,古树树干腐烂及空洞面积占比相对较低,高海拔地区温度较低,古树树干腐烂及空洞面积占比相对较高。 同样,在本研究结果中,降水量较高时,古树树干的腐烂及空洞面积占比较高。 该结果与海拔及古树树干腐烂面积占比间关系一致,低海拔地区降水量较低,古树树干腐烂及空洞面积占比较低,高海拔地区降水量较高,树干腐烂及空洞面积占比较高。 另外,热湿比在一定程度上代表环境的干旱程度,热湿比越大说明环境湿度越小(Wanget al,2006)。 以往研究表明,干旱环境下物种木材密度倾向于增大以维持内部水分平衡,减少水分丧失(Martinez-Cabreraet al,2009)。 同时其与树干空洞间存在紧密联系,空洞面积倾向于随着木材密度的增大而减小(Jacobsenet al,2007)。 本研究结果显示,热湿比较高时古树树干腐烂及空洞面积占比较低,符合以往的研究结论。
(3) 数据分析结果显示,在海拔1 000 m 以下,古树的腐烂及空洞面积均值为13.80%,古树树干腐烂及空洞面积占比相对较低;在海拔1 000 m 以上,古树的腐烂及空洞面积均值为30.82%,树干腐烂及空洞面积占比相对较高,且两者间存在较大差异。 结合马原(2007)对酸雨与天目山柳杉种群间关系的研究和郑世伟等(2015)对天目山酸雨降水范围主要在高海拔地区的研究,推测古树腐烂及空洞面积占比与酸雨之间存在一定联系,但在本研究中未进行进一步研究。
5 展望
本研究对浙江西天目山的11 个树种的137 株古树的腐烂及空洞面积占比进行了较为系统的调查和研究,并强调了古树健康与海拔、气候等的相关性,为古树保护和管理提供了科学依据。 但本研究尚存在一些不足之处,有望在接下来的工作中继续深入。
首先,由于研究区域和研究方法的限制,在古树的物种和数量的选择上存在一定限制。 例如,我们所用的测量设备PICUS 只能对胸径40 cm 以上的树木进行测量,这导致一些物种很难在各个海拔梯度上找到满足最低测量要求的树木。 在后续的古树健康研究工作中,应进一步完善对不同物种、不同海拔范围内古树健康状况的研究。
其次,本研究分析所使用的气候数据是基于全球粗分辨率的气候数据而获取的,这也是目前普遍采用的、可以公开获得的数据。 但由于这些数据空间和时间分辨率的限制,很难准确地描述各海拔段的古树所在微生境的气候条件。 在今后的调查检测中应加强对每株古树的立地环境和气候条件的同步调查,进而深入分析土壤、水文、地形、气候等对古树健康状况的综合影响。
最后,需要指出的是,正如生态学里的很多其它研究内容(如群落结构、物种多样性、林下幼苗更新)一样,古树健康状况在时间尺度上存在着极大的变化,这也给古树的保护和管理提出了挑战。 因此,需要对古树健康进行长时间的监测,以期掌握各物种健康状况的变化规律,为全球变化背景下的古树保护提供理论依据和实践基础。
