植物生长调节物质与草坪草及牧草的非生物逆境应答
2019-12-19产祝龙刘梦垚
产祝龙,张 慧,刘梦垚
(华中农业大学园艺林学学院 / 教育部园艺植物生物学重点实验室 / 农业部华中都市农业重点实验室,湖北 武汉 430070)
植物在生长发育过程中,面临着多种非生物逆境,包括温度、干旱、涝害、盐渍、矿物质缺乏、离子毒害等,造成植物细胞膜系统损坏、细胞脱水、光合速率、呼吸速率下降,从而影响植物生长。在逆境条件下,植物的应激反应引起内源多种激素及代谢物合成、分解及信号传导途径的变化,激发植物下游的防御反应,导致抗逆性的产生。近年来的研究表明[1-3],植物激素、小分子物质、渗透调节物质以及气体分子在植物逆境应答中起到重要的调控作用。草坪草和牧草在庭院绿化、固土护坡、生态治理、环境保护、畜牧业发展及保持人类健康中起到了重要的作用。本文对植物生长调节物质提高草坪草和牧草抗逆性的机理进行综述,以期为草坪保护剂开发和抗逆育种提供理论基础。
1 植物激素和生长调节物质
1.1 脱落酸
1.1.1 脱落酸对植物抗性的影响
脱落酸(abscisic acid, ABA)是一种倍半萜类小分子物质,它在植物根部和顶芽中产生,ABA 的合成和分解受到多种胁迫的影响[2-3]。研究表明,ABA 和ABA 受体PYR/PYLs 等结合以后,抑制了ABA 信号传导途径负调节因子PP2C 磷酸酶的活性,激活了激酶SnRK2。SnRK2 自身磷酸化,并磷酸化ABF 类转录因子,启动下游的ABA 应答通路基因的表达,从而提高了植物的抗逆性[1-3]。
ABA 不仅在提高草坪草抗旱性有明显效果,也能提高草坪草的对低温、高温、盐等胁迫的抗性。研究发现,干旱处理增加了草地早熟禾(Poa pratensis)、匍匐剪股颖(Agrostis stolonifera)、绒毛剪股颖(A. canina)、细弱剪股颖(A. capillaris)、高羊茅(Festuca arundinacea)等多种草坪草内源ABA的含量[4-6]。外源ABA 提高了草地早熟禾和匍匐剪股颖的抗旱和抗盐性,但是草地早熟禾上的效果更明显[7]。ABA 处理以后,草地早熟禾和匍匐剪股颖在干旱条件下APX、POD、SOD 的活性增强,MDA 的积累减少,蒸腾速率降低,叶片的相对含水量和水势增加,同时诱导了三羧酸循环相关有机酸的积累。ABA 处理的草坪草在干旱处理28 d后仍然维持在较好的草坪品质[7-8]。外源ABA 处理高羊茅通过减少气孔导度,降低蒸腾速率、净光合效率和MDA,提高抗氧化酶活性和逆境蛋白积累来增强抗旱性[9-10]。ABA 处理还可以通过增加脯氨酸、可溶性糖的含量,以及抗氧化酶的活性和相关基因的表达水平来提高狗牙根和多年生黑麦草(Lolium perenne)的抗旱性[11-12]。
狗牙根(Cynodon dactylon)和垂穗披碱草(Elymus nutans)在低温驯化阶段,体内ABA 含量增加,但是在脱驯化过程中,ABA 含量降低。外源ABA 处理降低了垂穗披碱草活性氧的积累,而ABA 抑制剂fluridone 处理促进了膜脂过氧化[13-14]。外源ABA类似物AM1 处理降低了狗牙根细胞膜的损伤程度,减少了活性氧的含量,同时诱导了抗氧化酶基因和DREB1 等基因的表达,增强狗牙根幼苗对冻害的抗性[15]。在低温条件下,ABA 处理的柱花草(Stylosanthes guianensis) APX 和CAT 活性,以及还原型谷胱甘肽和抗坏血酸和含量明显增加,抗寒性增强[16]。盐胁迫条件下,抗性匍匐翦股颖品种叶片中积累更多的ABA、JA、SA、ZR,而根当中积累的IAA 和SA 含量较高[17]。但是高温条件下匍匐翦股颖对于ABA 和乙烯含量的影响不大,反而在恢复阶段ABA 和乙烯含量增加[18-19]。外源ABA处理提高了蒺藜苜蓿(Medicago truncatula)中内源NO 和脯氨酸的含量,增强了蒺藜苜蓿对渗透胁迫的抗性[20]。紫花苜蓿在ABA 处理以后,固氮能力增强,多胺含量增加,对盐胁迫的抗性增强[21]。
1.1.2 脱落酸途径相关基因与抗逆性的关系
研究发现,高羊茅ABA 受体PYR1 可以与拟南芥PP2C 蛋白相互作用,激活ABA 信号通路,提高拟南芥对干旱胁迫的抗性[22]。匍匐剪股颖AsEXP1基因功能研究发现,该基因受到ABA 和SA 的共同调控[23],而在匍匐剪股颖中过表达拟南芥DREB1A基因,可以提高匍匐剪股颖ABA 和IAA 的含量和对渗透胁迫的抗性[24]。过表达匍匐剪股颖AsHSP17也诱导了ABA 合成、分解以及信号传导相关基因的表达[25]。玉米黄素环氧化酶(ZEP)是ABA 前体合成中的关键酶。紫花苜蓿MsZEP 在根中的表达受到ABA 的抑制。在烟草中过表达MsZEP,提高了干旱和盐胁迫条件下ABA 的含量,增强了植物的抗旱性[26]。
1.2 褪黑素
1.2.1 褪黑素对植物抗性的影响
褪黑素(melatonin, MT)是脊椎动物体内像松果般大小的“松果腺”器官分泌的一种吲哚胺类激素,是迄今发现的最强的内源性自由基清除剂。1995 年以来,研究发现植物中也有褪黑素存在,在植物抗逆性、开花和昼夜节律变化方面起到重要的调控作用[27-28]。
匍匐剪股颖在褪黑素处理以后,叶绿素含量和叶片水分含量增加,活性氧的产生减少,JUB1和DREB2A等基因表达量上调,延缓了干旱胁迫导致的衰老[29]。在高温条件下,外源褪黑素处理增加了多年生黑麦草内源褪黑素和细胞分裂素的含量,但是降低了内源ABA 的含量[30]。外源褪黑素延缓了黑暗诱导的多年生黑麦草的衰老[31]。在狗牙根中,外源褪黑素处理通过调控基因表达、蛋白质含量以及代谢物的产生,从而提高干旱、盐以及低温胁迫的抗性[32]。干旱、盐、低温和氧化胁迫也能提高狗牙根体内褪黑素的含量。外源褪黑素处理的狗牙根,体内抗氧化酶活性增强,活性氧的含量降低,膜脂过氧化程度减轻,细胞膜的损伤程度减小,光合效率增强[33-34]。高羊茅的抗热性在褪黑素处理以后得以增强。褪黑素处理诱导了高羊茅体内FaHSFA3、FaAWPM和FaCYTC2基因的表达,但是抑制了FaF-box、FaHSFA6B和FaCYP710A基因的表达[35]。褪黑素影响了紫花苜蓿脯氨酸代谢和一氧化氮(NO)途径并增强了抗旱性[36],同时通过增加H2O2的含量而刺激侧根的发育[37]。
1.2.2 褪黑素途径相关基因与抗逆性的关系
在拟南芥中过表达紫花苜蓿褪黑素合成SNAT基因,H2O2的含量增加,侧根的数目增多,侧根发育相关的基因受到褪黑素处理的诱导表达[38]。在拟南芥中过表达紫花苜蓿褪黑素合成相关的SNAT基因,提高了内源褪黑素的含量,增强了苜蓿中与镉抗性相关的ABA transporter 以及拟南芥中PDR8基因的表达,减少了镉的吸收[38]。
1.3 多胺
1.3.1 多胺对植物抗性的影响
多胺(polyamines, PAs)是一类具有强烈生物活性的低分子量脂肪族含氮碱。高等植物中主要有5 种多胺,包括腐胺(putrescine)、尸胺(cadaverine)、亚精胺(spermidine)、 精胺(spermine)和鲱精胺(agmatine)。植物体中多胺合成途径主要有两条。一条是以鸟氨酸为前体,在鸟氨酸脱羧酶(ODC)作用下直接脱羧形成腐胺。另一条是从头合成途径,以精氨酸为前体,在一系列酶的作用下生成腐胺。腐胺形成后,通过连续加入氨丙基残基而逐步形成精胺和亚精胺[39]。
研究表明,多胺预处理提高了匍匐剪股颖干旱胁迫下的草坪品质和抗旱性[40]。多胺降低了干旱条件下匍匐剪股颖IAA 和GA3的含量[41],调节与能量代谢、转运、抗氧化、胁迫等途径相关基因的表达[42]。狗牙根在3 种多胺处理以后,对于干旱和盐胁迫的抗性都明显提高。与电子传递和能量代谢途径相关的蛋白受多胺处理的诱导[43]。亚精胺处理通过改善光合系统和活性氧代谢提高了高羊茅对高温的耐受性[44]。外源亚精胺处理提高了内源多胺和脯氨酸的含量并增强了草地早熟禾和结缕草对盐胁迫的耐受性[45-47]。通过蛋白质组学分析,在白三叶(Trifolium repens)中鉴定了273 个受亚精胺处理发生含量变化的蛋白质[48]。
1.3.2 多胺途径相关基因与抗逆性的关系
过表达黄花苜蓿ERF 转录因子提高了与多胺合成以及代谢基因的表达,提高了脯氨酸含量,增强了抗寒性[49]。S-腺苷基甲硫氨酸合成酶(SAMS)是多胺前体合成的关键酶。紫花苜蓿SAMS1 基因受到低温、ABA、H2O2和NO 的诱导表达。过表达SAMS1 基因促进了多胺代谢途径相关基因的表达,增强了转基因紫花苜蓿的抗寒性[50]。在假俭草中过表达狗牙根多胺合成CdSAMDC 基因,提高了精胺、亚精胺以及NO 的含量,增强了低温抗性[51]。
1.4 细胞分裂素
1.4.1 细胞分裂素对植物抗性的影响
细胞分裂素(cytokinin, CK)是一种N6-异戊烯腺嘌呤。天然的细胞分裂素有玉米素(ZT)、玉米素核苷(ZR)、二氢玉米素(DHZ)、异戊烯基腺苷(iPA)等。而常见人工合成的细胞分裂素主要有激动素(KT)、6-苄基腺嘌呤(6-BA)和四氢吡喃苄基腺嘌呤(PBA)。高等植物的细胞分裂素主要是通过从头合成途径生成[52]。
研究发现,海藻提取物和腐殖酸中含有丰富的细胞分裂素成分,可以有效提高匍匐剪股颖对干旱和高温胁迫的抗性,改善草坪品质[53]。反式玉米素核苷(t-ZR)处理提高了匍匐剪股颖对干旱和高温的耐受性[54-55]。狗牙根和结缕草在低温驯化阶段,体内t-ZR 含量降低,但是在脱驯化过程中,t-ZR含量增加[14,56]。匍匐剪股颖在细胞分裂素处理以后,高温下氨基酸的积累明显高于没有处理的对照[57]。细胞分裂素处理延缓了黑暗诱导一年生黑麦草(L. multiflorum)叶片的衰老[58]。6-BA 处理多年生黑麦草,抑制了Na+的积累并保持较高的K+/Na+比,提高了的抗盐性[59]。
1.4.2 细胞分裂素途径相关基因与抗逆性的关系
腺嘌呤异戊烯基转移酶(ipt)是细胞分裂素合成的关键基因。在匍匐剪股颖中过表达SAG12 和HSP18.2 启动子驱动的ipt 基因,可以提高内源细胞分裂素的含量,促进了干旱条件下根系的发育,有效增强匍匐剪股颖对干旱胁迫和渗透胁迫的抗性[60]。匍匐剪股颖中SAG12-ipt转基因调控了107 个转录因子的表达,激活JA 和IAA 途径相关基因、但抑制了ABA 途径基因的表达[61-62]。转SAG12-ipt 基因的匍匐剪股颖在黑暗和高温条件下,内源细胞分裂素含量明显高于野生型,叶绿素的降解显著低于野生型,有效延缓了黑暗或高温诱导的衰老[63]。
1.5 水杨酸
1.5.1 水杨酸对植物抗性的影响
水杨酸 (salicylic acid, SA)是一种天然存在的酚类化合物。外源SA 处理显著提高保护酶的活性,降低MDA 和脯氨酸的积累,可以有效地缓解干旱胁迫对结缕草和匍匐剪股颖的伤害[64-65]。SA 处理还提高了沟叶结缕草(Z. matrella)和紫花苜蓿的抗寒性[66-67]以及匍匐剪股颖和草地早熟禾的抗热能力[68-69]。董慧等[70]研究表明,水杨酸能够提高多年生黑麦草的抗氧化酶活性,减轻盐胁迫造成的氧化伤害。外源SA 处理紫花苜蓿根瘤,会抑制盐胁迫诱导的多胺合成和活性氧代谢,增加ACC 的含量,提高紫花苜蓿在盐胁迫下生长速率[71]。
1.6 油菜素内酯
1.6.1 油菜素内酯对植物抗性的影响
油菜素内酯(brassinosteroids, BRs)是一类新的植物类固醇激素。2,4-表油菜素内酯(EBL) 和28-高油菜素内酯(HBL)是两种最常用的和商业化的油菜素内酯。
研究表明,BR 处理增强高羊茅水分利用效率,提高叶绿素的含量和抗氧化酶的活性,抗旱性增强[9]。高羊茅中高温诱导的基因如FaCRK8、FaCML38、 FaHSFB2B以 及FaHSFA3等 都 受 到EBL 处理的诱导表达[35]。在盐胁迫下,EBL 处理促进了多年生黑麦草和高羊茅在盐胁迫下的生长势,提高了多年生黑麦草中脯氨酸、可溶性糖、可溶性蛋白等的含量,增强了抗盐性[72-73]。寇江涛和师尚礼[74]发现EBR 显著提高盐胁迫下紫花苜蓿种子萌发和幼苗的生长发育。
1.6.2 油菜素内酯途径相关基因与抗逆性的关系
在匍匐剪股颖中过表达拟南芥BR 降解有关的AtBAT1 基因,转基因植株表现出BR 缺失的表型,植株变矮,节间变短,叶片夹角变小。转基因匍匐剪股颖对干旱胁迫的耐受性增加,干旱诱导的衰老减缓[75]。
1.7 乙烯
1.7.1 乙烯对植物抗性的影响
研究表明,高温条件下匍匐剪股颖乙烯含量没有明显变化,但是在恢复阶段乙烯(ethylene)的含量明显增加[19]。AVG 处理抑制了高温诱导的匍匐剪股颖的衰老[55]。在狗牙根中,乙烯前体ACC 处理以后,常温下调控了37 个蛋白、低温下调控了60 个蛋白质含量的变化,增强了抗寒性[76]。外源乙烯处理降低了蒺藜苜蓿对低温的抗性,而乙烯合成抑制剂处理提高了蒺藜苜蓿的抗寒性[77]。
1.7.2 乙烯途径相关基因与抗逆性的关系
乙烯不敏感突变体skl 对低温的耐受性增强。低温驯化可以提高野生型但不能增强skl 突变体的抗寒性。乙烯应答响应基因如EIN3 和ERF 基因的表达量在skl 突变体中显著下调[77]。紫花苜蓿乙烯应答因子MsERF8 受到盐、PEG 和多种激素的诱导表达。MsERF8 过表达使烟草积累了大量的脯氨酸,丙二醛含量降低,增强了对盐胁迫的耐受性[78]。缺磷促进了黄花苜蓿乙烯的产生,但是这种诱导作用被乙烯合成抑制剂AVG 和CoCl2所抑制[79]。
1.8 生长素
1.8.1 生长素对植物抗性的影响
生长素(indole-3-acetic acid, IAA)是一类含有一个不饱和芳香族环和一个乙酸侧链的内源激素,包括吲哚乙酸(IAA)、4-氯-IAA、5-羟-IAA 等。人工合成的生长素类似物,如2, 4-D、萘乙酸(NAA)、吲哚丁酸(IBA)等,具有与生长素相似的生理效应。
适宜浓度的IAA 能够有效提高白三叶幼苗对干旱环境的适应能力[80]。干旱胁迫降低了匍匐剪股颖内源IAA 和iPA 的含量。外源IBA 处理增强了干旱条件下植物的光合作用,提高了叶片IAA、iPA 以及ABA 的含量,增强了抗旱性[81]。接种产IAA 的根瘤菌以后,蒺藜苜蓿在盐胁迫条件下衰老延迟,乙烯信号相关基因的表达受到抑制,固氮能力增强,对盐胁迫的抗性提高[82]。铝胁迫抑制了紫花苜蓿顶芽和根尖IAA 的浓度,减少了茎尖IAA 向根尖的运输,与生长素运输有关的AUX1 和PIN2 基因的表达上调[83]。
1.8.2 生长素途径相关基因与抗逆性的关系
目前在草坪草和牧草生长素途径相关基因功能的研究较少。通过转录组分析,发现蒺藜苜蓿在盐胁迫处理后,与生长素运输有关PIN9 和PIN10基因大量表达[84]。通过生物信息学分析,在蒺藜苜蓿中鉴定一些与生长素代谢及信号传导相关的基因,包括Aux/IAA 基因[85]、生长素转运蛋白基因[86]、生长素应答MtARF 基因[87]等。
1.9 赤霉素
1.9.1 赤霉素对植物抗性的影响
赤霉素 (gibberellin, GA)是植物体内的一种四环双萜类化合物。外施赤霉素后,紫花苜蓿幼苗叶绿素含量都有所提升,在一定程度上提升了其光合效率,增强了抗旱性[88]。外源赤霉素溶液处理也能够在不同程度上提高碱茅种子的耐盐能力[89]。GA3处理也提高了紫花苜蓿的萌发率和盐胁迫下的干重[90]。
1.9.2 赤霉素途径相关基因与抗逆性的关系
结缕草GA 合成关键酶ZjGA20 氧化酶基因在叶片和茎中表达量较高。低温、高温和渗透胁迫抑制了ZjGA20 的表达。外源GA3处理增强了POD的活性,减少了丙二醛的产生,提高了结缕草的抗寒性[91]。在百喜草(Paspalum notatum)中过表达拟南芥GA 降解有关的AtGA2ox1基因,活性GA1的含量降低,茎杆缩短,开花延迟,但是分枝增多,修剪后恢复性优于对照[92]。
1.10 独脚金内酯
1.1 0.1 独脚金内酯对植物抗性的影响
独脚金内酯(strigolactones, SLs)是一种新型的植物激素,是从植物根际分泌物中被分离出来作为刺激根寄生植物独脚金种子发芽的物质。外源独脚金内酯处理抑制了百脉根(Lotus corniculatus)中干旱诱导的ABA 的产生,下调了ABA 合成相关基因NCED2 的表达[93]。独脚金内酯类似物GR24处理促进了对照和高温条件下高羊茅根系细胞的数量,上调了细胞循环相关的基因,下调了生长素转运基因,促进了根系的生长[94]。
1.1 0.2 独脚金内酯途径相关基因与抗逆性的关系
百脉根中沉默独脚金内酯合成相关的CCD7 基因,引起气孔导度的增加,ABA 处理诱导的气孔关闭减缓,抗旱性减弱[93]。干旱条件下高羊茅内源独脚金内酯含量增加,独脚金内酯合成相关的FaD17 和信号传导相关的FaD14 以及FaMAX1 基因的表达上调。独脚金内酯含量的增加可能是干旱胁迫对侧芽生长抑制的主要因素[95]。
1.11 茉莉酸酯对植物抗性的影响
茉莉酸酯(jasmonate acids, JAs)包括茉莉酸(JA)和茉莉酸甲酯(MeJAs),是存在于所有高等植物体内的内源调节物质。外施JAs 都能够有效提高植物的抗逆性,但目前在草坪草、牧草上的相关研究甚少。蒺藜苜蓿悬浮细胞在MeJA 处理以后,与三萜合成有关的香树脂醇合酶基因上调了50 倍以上,同时在鉴定到的30 多个三萜皂甙类物质中的大部分在MeJA处理后含量增加[96]。
2 渗透调节物质
2.1 可溶性糖
在越冬阶段,盐草(Distichlis spicata)的果糖、葡萄糖、棉子糖和水苏糖含量有着明显的季节性变化趋势,在隆冬季节含量最高[97]。低温诱导了紫花苜蓿蔗糖、棉子糖和水苏糖的积累,降低其了果糖、葡萄糖和淀粉的含量[98]。草地早熟禾和结缕草的抗寒性分别与果聚糖和蔗糖,以及总还原糖和果糖含量相关[99-100]。李丽群等[101]指出外施蔗糖处理可以明显改善高羊茅植株的生长,提高其耐阴性。连俊方等[102]研究表明外源海藻糖可缓解盐胁迫对多年生黑麦草生长发育的抑制作用。在紫花苜蓿中过表达海藻糖合成相关的TPS1 和TPS2基因,增加了内源海藻糖的含量,增强了紫花苜蓿对干旱、低温、盐和高温胁迫的耐受性[103]。壳聚糖能够提高渗透胁迫下沿阶草和草地早熟禾脯氨酸含量,降低渗透势,增强抗旱和抗盐能力[104-105]。
2.2 甜菜碱
甜菜碱(glycine betaine, GB)是一种水溶性生物碱,广泛存在于植物中。研究表明,外源甜菜碱处理能够增加内源性甜菜碱的含量,提高抗氧化酶的活性,增强匍匐剪股颖和高羊茅的抗旱性[106-107]。根施适宜浓度的甜菜碱可以有效减轻盐胁迫对黑麦草幼苗的伤害[108]。外源甜菜碱能够有效缓解低温胁迫下结缕草的草坪质量,增强结缕草对低温胁迫的抵抗性[109]。甜菜碱处理可同时提高草地早熟禾和匍匐剪股颖的抗旱和抗盐性[7]。
2.3 脯氨酸
脯氨酸(proline)是一种兼容性小分子,在高等植物中广泛存在,有助于稳定细胞结构,清除自由基以及在胁迫条件下调节细胞氧化还原状态。结缕草内源脯氨酸的含量与抗冻性正相关[100]。渗透胁迫导致脯氨酸在蒺藜苜蓿中的积累。MtP5CS2在对照条件下表达量极低,但是盐胁迫处理后在根中的表达量急剧增加[110]。在蒺藜苜蓿中过表达乌头叶豇豆(Vigna aconitifolia) P5CS 基因,增加了蒺藜苜蓿内源脯氨酸的含量,增强了盐胁迫条件下的固氮能力[111]。紫花苜蓿脯氨酸脱氢酶(MsDH)基因的表达受到盐胁迫的显著抑制,造成内源脯氨酸的积累,增强了抗盐性[112]。
2.4 无机离子类物质
2.4.1 钙
钙(calcium, Ca2+)是植物必需的营养元素,同时是胞内生理生化反应的第二信使。研究表明,外源钙提高了紫花苜蓿的抗旱性[113]。在盐胁迫下,外源钙处理减少了高羊茅对Na+的吸收,提高植物耐盐性,同时促进了高羊茅在盐胁迫下的萌发率[114-115]。高温条件下外源Ca2+处理提高了草地早熟禾和高羊茅抗氧化酶的活性,减少了丙二醛的产生,提高了抗热性[116]。Ca2+信号也参与了ABA 途径。研究发现Ca2+信号抑制剂LaCl3拮抗ABA 提高柱花草抗寒性的作用[117]。在干旱胁迫下,外源钙离子处理提高了紫花苜蓿和结缕草叶片相对含水量、叶绿素含量、POD 活性,降低了细胞膜透性,提高保护酶的活性,进而增强紫花苜蓿和结缕草的 抗旱性[113,118-119]。
2.4.2 硅
硅(silicon, Si)是一种重要的植物成分或代谢物,主要以氧化物和硅酸盐的形式存在,对草坪草的生长有促进作用[120]。董文科等[121]指出在干旱、高温及双重胁迫下,外源硅酸盐能够显著缓解各种胁迫对黑麦草种子萌发的影响,清除氧自由基,减轻胁迫的伤害。硅还提高了海滨雀稗体内脯氨酸和糖的含量、增强了抗氧化酶活性,降低了丙二醛的含量,增强抗寒性[122]。添加外源硅提高了狗牙根、高羊茅和多年生黑麦草对高盐胁迫的抗性[123],同时增强了草地早熟禾对盐和干旱胁迫的耐受性[124-125]。外源硅可以明显缓解铝胁迫下鸭茅植株的生长,并降低植株对铝离子的吸收[126]。
3 气体分子
3.1 硫化氢
研究表明,硫化氢(hydrogen sulfide, H2S)在低浓度时能够起到一种气态信号分子的作用[127-128]。王燕琴[129]研究表明,硫化氢能够缓解盐胁迫导致的种子萌发和幼苗生长受抑制的现象。经硫化氢的外源供体NaHS 处理的狗牙根材料,胁迫过程中活性氧积累少,抗氧化酶活性高,渗透调节物质积累多,细胞膜损伤小,对盐、干旱和低温胁迫的抗性增强[130]。NaHS 处理还能够通过调节酶促和非酶促抗氧化剂的活性,减轻镉胁迫对狗牙根生长的抑制和细胞损伤[131]。在紫花苜蓿中,NaHS 促进了NO 的产生,增强了抗氧化酶活性,提高了K+/Na+比,促进了紫花苜蓿在盐胁迫下的萌发率[132]。
3.2 一氧化氮
一氧化氮(nitric oxide, NO)是一种兼具水溶性和脂溶性特点的自由基气体信号分子。研究表明,盐胁迫下,外源NO 对高羊茅种子的萌发和对幼苗有保护作用,能够明显促进紫花苜蓿幼苗根系中NO 的积累,提高抗氧化系统的活性,降低自由基含量和膜质过氧化作用,缓解盐胁迫和干旱对紫花苜蓿和狗牙根的伤害[132-135]。在渗透和干旱胁迫下,外源NO 可显著减轻胁迫对多年生黑麦草、匍匐剪股颖和高羊茅生长的抑制,增强其抗渗透胁迫和干旱胁迫的能力[136-137]。NO 供体SNP处理调控了多胺和脯氨酸代谢相关酶基因的表达,增加了蒺藜苜蓿腐胺和脯氨酸的含量[138]。外源NO 处理抑制了蒺藜苜蓿IAA 氧化酶的活性和IAA 的降解,促进了多年生黑麦草在镉胁迫中对多种元素的吸收,提高了蒺藜苜蓿和多年生黑麦草对镉的耐受性[139-140]。
3.3 一氧化碳
一氧化碳 (carbon monoxide, CO)作为信号分子参与植物体内重要的生理代谢过程。在对紫花苜蓿的研究中发现,外源CO 处理能够缓解镉胁迫对苜蓿根的氧化伤害,并提高SOD、POD、GST 等酶的活性,调节植物体内ROS 代谢,缓解等胁迫下对根系的损伤[141]。
4 其他小分子物质
4.1 γ-氨基丁酸
γ-氨基丁酸(γ-aminobutyric acid, GABA)是一种非蛋白氨基酸。研究表明,外源GABA 的施用能够有效缓解干旱引起的白三叶叶片损伤,提高相关保护酶活性,并导致内源谷氨酸和GABA 含量增加,从而增强白三叶的耐旱性[142]。GABA 处理匍匐剪股颖,在干旱条件下降低了电导率,提高了叶片相对水分含量,诱导了多种氨基酸和有机酸的积累,提高了抗旱性[65]。
4.2 5-氨基乙酰丙酸
5-氨 基 乙 酰 丙 酸(5-aminolevulinic acid, ALA)是四氢吡咯生物合成的前体。干旱胁迫下,ALA 处理能明显提高高羊茅的草坪质量、叶片相对含水量、叶绿素含量、可溶性糖含量,增强了抗旱能力[143]。低浓度的ALA 可以缓解盐胁迫对匍匐剪股颖的伤害,而高浓度则会加重盐胁迫的伤害作用,同时ALA 还可以降低盐胁迫下植株对Na+的过量吸收从而缓解盐胁迫的伤害作用[144]。林艳艳等[145]也指出外源施加低浓度ALA 能够促进草地早熟禾种子在逆境条件下的萌发、促进幼苗生长和可溶性蛋白的积累。低温条件下ALA 对垂穗披碱草的种子萌发起到保护作用,降低种子在低温胁迫下的氧化损伤[146]。
4.3 抗坏血酸
抗 坏 血 酸 (ascorbic acid, AsA)又 叫 做 维 生 素C,是一种水溶性抗氧化剂分子,能够清除植物各项代谢过程中产生的自由基。研究表明,抗坏血酸能够提高高羊茅在盐胁迫下的根冠比和含水量,增强膜的稳定性从而提高耐盐性[147]。AsA 处理提高了高羊茅非酶促抗氧化物质的含量、抑制ROS 的产生,促进水分胁迫条件下高羊茅根的生长[148]。
4.4 谷胱甘肽
谷胱甘肽(glutathione, GSH)是一种小的细胞内硫醇分子,被认为是一种强的非酶促抗氧化剂。谷胱甘肽能够清除细胞内产生的自由基,从而减少膜质过氧化对细胞膜造成的损伤。在草坪草中,非生物胁迫提高了多种草坪草谷胱甘肽的含量,但是外源谷胱甘肽处理的研究少[11,131]。
4.5 柠檬酸
柠檬酸 (citric acid)是三羧酸循环中间产物中一种重要的有机酸,调控植物生长和胁迫应答。在高羊茅抗高温研究中,发现柠檬酸途径受高温胁迫的诱导[149]。外源柠檬酸[150]处理可以减缓高温对高羊茅生长的抑制作用,提高了光合效率,降低了细胞膜透性。
5 结论与展望
5.1 结论
综上所述,多种植物激素和小分子物质提高了草坪草和牧草对非生物逆境的抗性。与模式植物中的研究结果类似,不同激素及小分子物质之间存在着协同或拮抗的关系。干旱和低温处理提高了草坪草中ABA 和脯氨酸的含量,但是降低了细胞分裂素的含量[4-5,17]。外源褪黑素处理提高了ABA的含量[13]。外源ABA 处理提高了蒺藜苜蓿中内源NO 和脯氨酸的含量[20]。紫花苜蓿在ABA 处理以后多胺含量增加[21]。在高温条件下,外源褪黑素处理增加了内源褪黑素和细胞分裂素的含量,但是降低了内源ABA 的含量[30]。多胺预处理匍匐剪股颖后,GA1、GA4、GA20和ABA 的含量也有所提高[40]。但是也有证据表明多胺处理匍匐剪股颖材料降低了干旱条件下IAA 和GA3的含量[41]。在蒺藜苜蓿中腐胺的含量在NO 供体处理以后增加[138]。假俭草中过表达与多胺合成相关的狗牙根CdSAMDC基因增加了NO 的含量[51]。外源EBL 处理提高了多年生黑麦草内源ABA、ZR、JA 和SA 的含量[73]。在匍匐剪股颖上的研究发现,t-ZR 处理减少了乙烯的产生[55]。外源IBA 处理提高了匍匐剪股颖叶片IAA、iPA 以及ABA 的含量[81]。另外,不同的外源激素和小分子物质处理对草坪草的影响也有差异。ABA、SA 和GABA 诱导处理都提高了匍匐剪股颖的抗旱性,但是诱导了不同物质的积累[65]。外源SA、ABA、钙、H2O2以及乙烯分别处理以后,发现对于高温胁迫下匍匐剪股颖抗氧化酶活性的影响具有一定的差异性[68]。这些说明多种植物激素和小分子物质协同或拮抗地影响草坪草和牧草对外界逆境条件的应答。
植物逆境应答是一个复杂的网络信号。外界的环境因素对草坪草和牧草的影响也是多方面的。在不利的生长环境下,草坪草体内脱落酸、赤霉素、褪黑素、多胺、水杨酸、独角金内酯的含量明显提高,而细胞分裂素和生长素的含量显著降低。不同的植物激素之间存在协同或拮抗作用。内源植物激素和小分子物质含量的变化,调控了下游相关基因的表达,包括合成/分解代谢、信号传导途径、活性氧代谢、胁迫应答途径、能量代谢、蛋白质代谢等。草坪草从而在生理水平上做出相应的调整。脯氨酸、可溶性糖、抗氧化酶活性、光合作用、渗透调节物质的含量提高。相应的,活性氧含量、丙二醛含量和电导率降低,从而达到了延缓衰老,提高抗逆性和改善草坪品质的效果(图1)。当然,草坪草和牧草中不同激素之间的相互作用关系很复杂,有些研究结果甚至是相反的,需要开展更多更深入的研究。
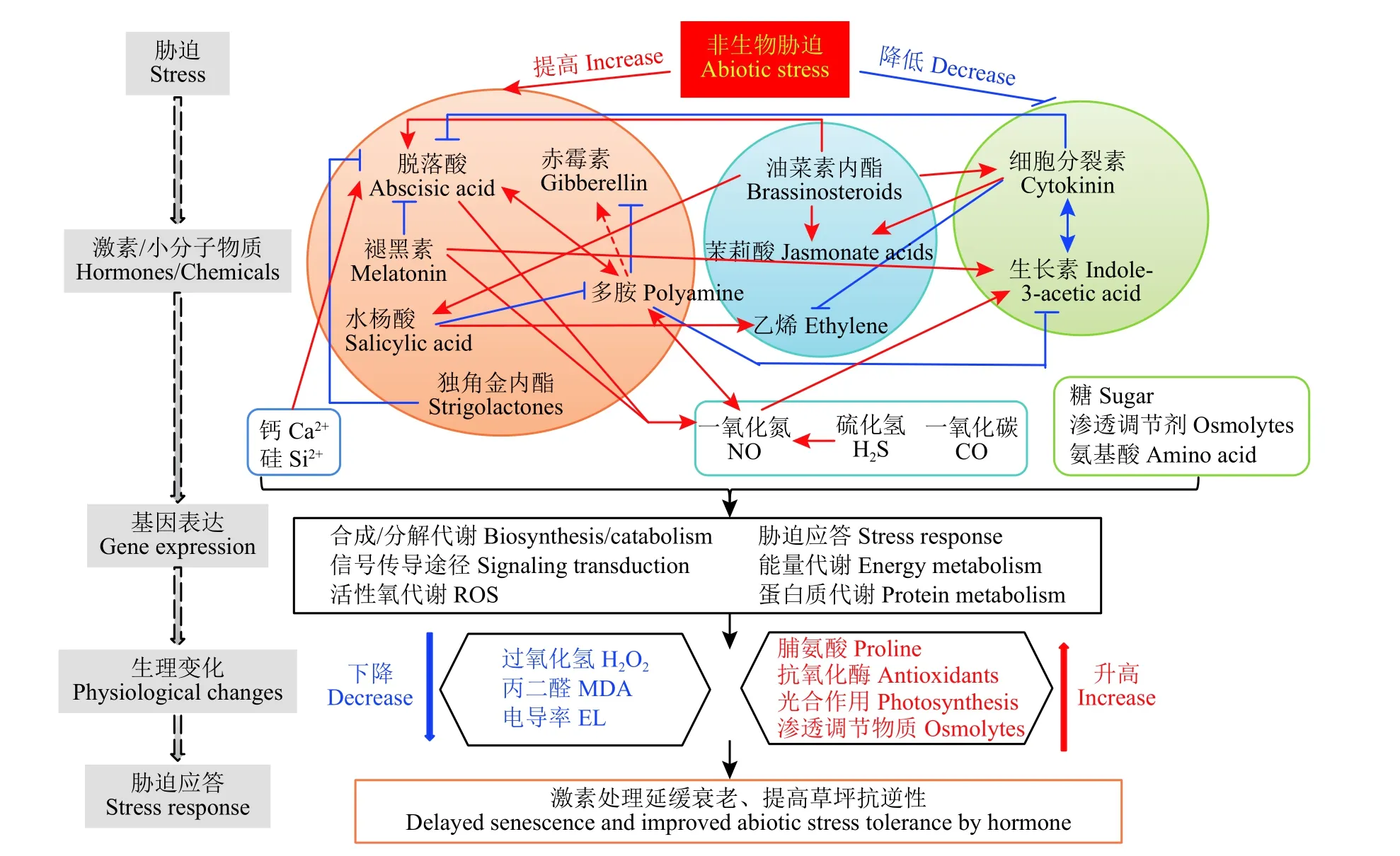
图 1 植物激素和小分子物质在草坪草和牧草逆境应答中的作用。Figure 1 Roles of plant hormones and small molecule compounds during the abiotic stress responses of turfgrass and forage
5.2 展望
近些年来在外源物质处理诱导草坪草和牧草抗逆性方面的研究取得了可喜进展。但是目前的研究更多地是集中在外源物质处理以后对草坪草和牧草抗逆性表型的观察比较和生理指标的测定,对于相关基因功能和调控网络方面的研究涉足较少。草坪草和牧草具有一些模式植物不具备的性状,例如冷季型草坪草对低温的抗性、暖季型草坪草对高温的耐受性、苜蓿与根瘤菌的共生固氮等。这种特异性表明草坪草和牧草相关基因的功能可能与模式植物的同源基因存在差异。
今后的研究建议聚焦到以下领域:1)继续对一些新型激素和小分子物质的效果进行筛选,如独脚金内酯和褪黑素等。谷胱甘肽和柠檬酸等在植物生长和胁迫应答中起到重要作用,目前在草坪草和牧草上应用和研究较少。2)开展多种激素和小分子物质复配效果的研究。不同的外源物质对草坪草和牧草抗逆性的诱导具有一定的特异性。探讨不同外源物质之间的协同作用关系,筛选复配配方,从而更有效提高草坪草和牧草的抗逆性。同时加强田间的试验,开发出生产上能真正应用的产品。3)开展草坪草和牧草激素合成分解及信号传导途径相关基因功能的研究;在模式植物中,多种植物激素的合成分解途径和信号传导途径得以解析(图2)。如ABA 合成的NCED,信号传导PYR/PYL-PP2C-SnRK-ABF;IAA 合成的YUC,信号传导TIR1/AFB2-AUX/IAA-ARF;ET 合成的ACS 和ACO,信号传导ETR-EIN2-EIN3/EILs-ERF1/EREBPs 等(图2)。目前在草坪草和牧草中,高羊茅的ABA 受体基因[22]、苜蓿褪黑素合成SNAT基因[37-38]、苜蓿多胺合成SAMS1 基因[50]、狗牙根多胺合成SAMDC 基因、匍匐翦股颖细胞分裂素合成IPT 基因[24,60]、结缕草赤霉素合成关键酶GA20ox基因[91]的功能得以部分阐述。但是多数草坪草及牧草中激素合成途径及信号传导途径基因的功能仍然是未知的,大量的工作仍然需要持续开展,从而真正揭示草坪草和牧草逆境应答的机理,构建抗逆调控网络。

图 2 植物激素和小分子物质的生物合成及信号传导途径。Figure 2 Biosynthesis and signaling transduction pathways of plant hormones and small molecule compounds.
