沉默lncRNA DANCR调节尼古丁胁迫下NSCLCs的炎症微环境、增殖活性和迁移
2019-12-11兰飞,赵鹏,刘冬
兰 飞,赵 鹏,刘 冬
(空军军医大学第二附属医院肿瘤科,西安 710038;*通讯作者,E-mail:liudong19791120@126.com)
肺癌是一种严重的致死性肿瘤疾病,是癌症相关死亡的主要原因[1]。据统计每年死于肺癌的患者比乳腺癌、前列腺癌和结肠癌三者的总和还要多[2]。非小细胞肺癌(non-small cell lung cancer,NSCLC)占肺癌病例约87%。长期的临床表明,外界刺激下多种炎症反应的增加是导致包括肺癌在内的多种癌症转移的主要原因之一,并称此为肿瘤相关炎症(cancer-related inflammation,CRI)[3]。CRI在非小细胞肺癌的恶性转化中发挥着关键作用,并介导上皮间充质转化(epithelial mesenchymal transformation,EMT),EMT也是癌细胞转移的基础[4]。研究表明,NF-κB炎症信号通路参与了肺癌的转移[5]。目前,有关对CRI与NSCLC关系及其调控机制的研究还较为局限。
长链非编码RNA(lncRNA,>200 nt)是近年来发现的一类关键的肿瘤相关RNA[6]。lncRNA具高度保守性,其通过调控基因转录因子、直接结合靶蛋白或扮演内源竞争性RNA(ceRNA)与其靶microRNA(miRNA)结合、修饰基因甲基化等方式调节多种抑癌基因或促癌基因的表达和功能[6,7]。lncRNA DANCR是一种关键的致癌lncRNA,其通过直接调控癌基因或者通过扮演ceRNA角色调控miRNA对靶基因的调节作用[8]。研究表明,DANCR在NSCLC组织和细胞系中表达上调,而且具有促进NSCLC细胞增殖、迁移、侵袭等作用[9]。近年研究发现,lncRNA DANCR在肺炎病理组织中诱导炎症因子的表达,介导免疫环境的发生[10],但其在NSCLC相关炎症中的作用并不清楚。
尼古丁(nicotine),又称烟碱,是烟草中的关键成分,具有成瘾性。尼古丁已被证明与肺癌的发生具有密切相关性[11],并具有促进NSCLC细胞增殖、EMT、抑制细胞凋亡、诱导侵袭和迁移的作用[12]。更重要的是尼古丁可诱导肺癌细胞和人支气管上皮细胞炎症反应[13,14]。因此,本研究拟借助尼古丁作为炎症环境的诱导因素,探索lncRNA DANCR是否介导NSCLC的炎症微环境,并进一步探索DANCR对尼古丁环境下诱导的非小细胞肺癌细胞增殖及迁移的影响,为NSCLC等癌症的CRI发生发展机制理解以及新的NSCLC炎症微环境标记物的筛选提供实验依据。
1 材料与方法
1.1 细胞株与主要试剂
人NSCLC细胞A549(中国科学院微生物所菌种保藏中心,上海);DANCR沉默质粒(siRNA-DANCR:si-DANCR;siRNA-NC:si-NC,BioVector质粒载体菌种细胞基因保藏中心,北京);肿瘤坏死因子(TNF)-α、IL-6及IL-8的检测试剂盒(Santa Cruz Biotechnology,美国);反转录试剂盒(Fermentas,德国);SYBR Green Master Mix(Life Technologies,美国);BCA蛋白检测试剂盒(Sigma,美国);兔源性的p-IκBα(ab133462)、p65-NF-κB(ab16502)、Lamin B1(ab16048)、GAPDH(ab124957)、辣根过氧化酶二抗及RPMI 1640培养基(Abcam,美国);TRIzol试剂盒(Invitrogen,美国);核蛋白抽提试剂盒(碧云天,海门);PVDF膜(Millipore,美国);Image-Pro Plus 6软件进行(Media Cybernetics,美国)。
1.2 人NSCLC细胞的培养与分组
采用常规培养方法培养A549细胞(37 ℃,5% CO2),使用含10% FBS及1%青-链霉素的RPMI 1640培养基。每2-3 d换液1次,待细胞融合率达到80%时,进行传代,并且将细胞浓度调整为1.0×106/ml用于后续实验。细胞分组如下:分别以不同浓度尼古丁(50,250,500 ng/ml)处理A549细胞8 h,根据尼古丁处理浓度分别命名为0 ng/ml组、50 ng/ml组、250 ng/ml组及500 ng/ml组[15]。此外,进一步研究中,A549细胞中的lncRNA DANCR以siRNA法沉默(见方法1.3)。siRNA转染48 h联合尼古丁处理8 h,根据尼古丁(0 ng/ml、500 ng/ml)及DANCR沉默(未处理、si-NC转染、si-DANCR转染)联合处理的不同,细胞分为6组:即0 ng/ml组、0 ng/ml+si-RNA组、0 ng/ml+si-DANCR组、500 ng/ml组、500 ng/ml+si-NC组和500 ng/ml+si-DANCR组。
1.3 si-DANCR质粒构建与细胞转染
具有稳定转染效率的si-DANCR及其si-NC质粒用于沉默A549细胞中的lncRNA DANCR。主要过程是对人源DANCR的cDNA使用RT-PCR进行扩增,并且将产物插入siRNA质粒,进行转染效率优化筛选。将浓度为1.0×106/ml的A549细胞转入24孔板中培养,使用20 μg si-NC质粒或者20 μg si-DANCR分别对A549细胞进行转染,未处理组为对照。转染进行48 h后,使用实时荧光定量PCR(RT-qPCR)法检测DANCR水平,以验证沉默效果。
1.4 ELISA检测细胞炎性因子分泌
收集各处理组的细胞,将细胞悬液离心(3 500g,15 min)。将上清液收集用于炎性因子分析。根据生产商的说明,炎症因子TNF-α、IL-6、IL-8使用ELISA试剂盒法进行检测。以上炎性因子的ELISA检测灵敏度分别是:TNF-α为4.32 pg/ml;IL-6为1.6 pg/ml;IL-8为<25 pg/ml。
1.5 RT-qPCR检测lncRNA DANCR及IKKβ水平
使用TRIzol试剂盒,按照使用说明书抽提细胞总RNA。使用反转录试剂盒将5.0 μg RNA作为模板合成cDNA,随后使用SYBR Green Master Mix进行基因表达的实时分析。扩增过程包括变性(95 ℃,5 min,1个循环)、延伸(95 ℃,5 s,40个循环)与退火(60 ℃,1 min,40个循环)。使用2-ΔΔCt法计算相对mRNA的表达量,每组数据均以倍数变化进行展示。GAPDH用于实时定量结果的标准化处理。DANCR引物序列Forward:5′-CGT CTC TTA CGT CTG CGG AA-3′,Reverse:5′-TGG CTT GTG CCT GTA GTT GT-3′;IKKβ引物序列Forward:5′-TAG CAT GAA TGC CTC TCG ACT-3′,Reverse:5′-TTC AGC CAC CAG TTC TTC ACT-3′;GAPDH引物序列Forward:5′-TGC ACC ACC AAC TGC TTA GC-3′,Reverse:5′-GGC ATG GAC TGT GGT CAT GAG-3′。
1.6 Western blot检测NF-κB炎症信号通路相关蛋白水平
收集各处理组的细胞进行裂解,按照供应商提供的方法抽提总蛋白和核蛋白(核蛋白抽提试剂盒)。使用12%的聚丙烯酰氨凝胶电泳SDS-PAGE分离总蛋白中不同大小的蛋白质,随后使用PVDF膜进行湿法转膜。使用3%的脱脂牛奶对转膜完成的PVDF膜进行室温封闭1.5 h。用PBS洗涤3×5 min,使用一抗包括兔源性的p-IκBα(1∶400,ab133462)以及GAPDH(1∶2 000,ab124957)4 ℃孵育过夜,洗去一抗,并且在室温使用辣根过氧化酶二抗(1∶18 000)孵育1.5-2 h,洗涤抗体液。随后使用ECL化学发光试剂盒在化学发光检测系统中进行显色。对于核蛋白的检测,电泳和转膜过程均与总蛋白一致,一抗使用兔源性的p65-NF-κB(1∶500,ab16502)以及Lamin B1(1∶5 000,ab16048),4 ℃孵育过夜。洗去一抗,并且在室温使用辣根过氧化酶二抗(1∶18 000)孵育1.5-2 h,洗涤抗体液。随后使用ECL化学发光试剂盒在化学发光检测系统中进行显色。Western blot定量分析采用Image-Pro Plus 6软件进行。
1.7 MTT实验细胞增殖活性
将各组待检测细胞的密度调整为5×103/ml,并接种于96孔板,加入MTT(终浓度为5 mg/ml),于37 ℃孵育4 h,弃上清液后加入DMSO。使用酶标仪检测6,12,24,48 h的490 nm处的吸光度值(OD)。计算细胞成活率。细胞存活率=(处理组OD值-空白对照组OD值)/(对照组OD值-空白对照组OD值)×100%。
1.8 细胞划痕实验检测细胞迁移能力
收集各处理组细胞至48孔板。使用RPMI 1640培养基常规培养细胞至融合率达到80%,换成无血清的RPMI 1640培养基培养24 h。使用无菌200 μl枪头尖端行培养板内侧底部划痕,吸除培养基,换加新鲜无血清培养基。用配有数码相机的Olympus IX70显微镜进行拍照,拍照时间为划痕后0 h及24 h。利用Image J 1.5软件计算划痕两侧的距离,测量划痕宽度。
1.9 统计学分析
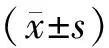
2 结果
2.1 不同浓度尼古丁诱导NSCLC细胞炎症微环境发生
与0 ng/ml尼古丁组相比,TNF-α、IL-6、IL-8水平在50,250,500 ng/ml尼古丁刺激下均显著增加,差异有统计学意义(P<0.05,见图1)。此外,TNF-α、IL-6、IL-8的水平增加均呈现尼古丁剂量依赖性(见图1)。以上结果证实,尼古丁可以剂量依赖性增加A549细胞的炎症因子分泌,表明尼古丁能够诱导NSCLC细胞炎症微环境的发生。
2.2 尼古丁刺激激活NSCLC细胞NF-κB炎症信号通路
选择500 ng/ml尼古丁剂量作为后续研究浓度。进一步结果表明,在500 ng/ml尼古丁刺激下,A549细胞中IKKβ mRNA水平和p-IκBα蛋白水平及细胞核内p65-NF-κB蛋白水平均显著增加,差异均具有统计学意义(P<0.05,见图2)。结果证实,在尼古丁刺激下,NSCLC细胞中的NF-κB炎症信号通路被激活。
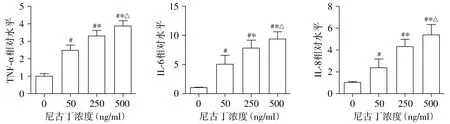
与0 ng/ml组相比,#P<0.05;与50 ng/ml组相比,*P<0.05;与250 ng/ml组相比,△P<0.05
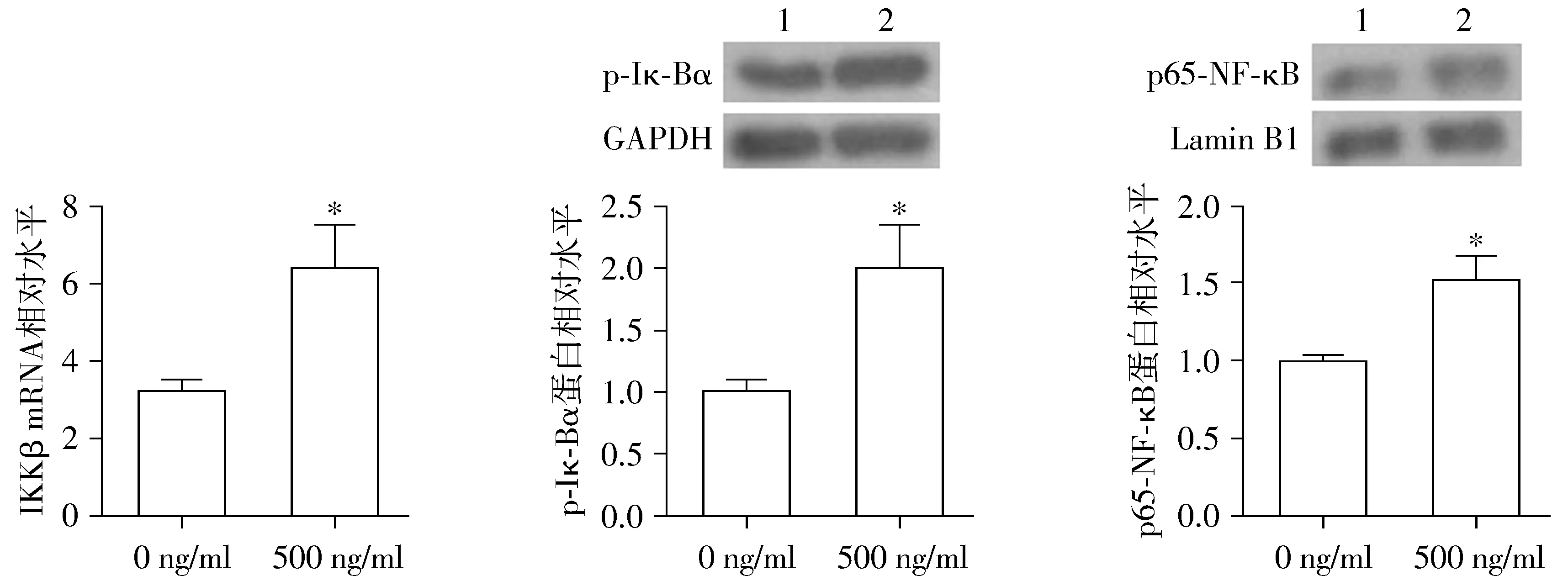
1.0 ng/ml尼古丁;2.500 ng/ml尼古丁;与0 ng/ml组相比,*P<0.05
2.3 沉默lncRNA DANCR能够抑制尼古丁诱导的NSCLC炎症反应
使用siRNA沉默DANCR,以研究DNACR的功能。同时,探究了单独沉默DANCR或尼古丁联合沉默DANCR作用下细胞中炎症反应的变化。结果显示,分别与0 ng/ml组和0 ng/ml+si-NC组相比,0 ng/ml+si-DANCR组中TNF-α、IL-6、IL-8及细胞核内p65-NF-κB水平均显著下降,差异有统计学意义(P<0.05,见图3);而0 ng/ml组与0 ng/ml+si-NC组相比,以上分子的水平无统计学差异(P>0.05,见图3)。此外,与0 ng/ml尼古丁组相比,由500 ng/ml尼古丁诱导的炎症因子和核内p65-NF-κB的水平增加均可被si-DANCR显著抑制,差异有统计学意义(P<0.05,见图3);而500 ng/ml组与500 ng/ml+si-NC组相比,以上分子的水平无统计学差异(P>0.05,见图3)。结果表明,沉默DANCR能够抑制由尼古丁刺激而诱发的NSCLC炎症反应。
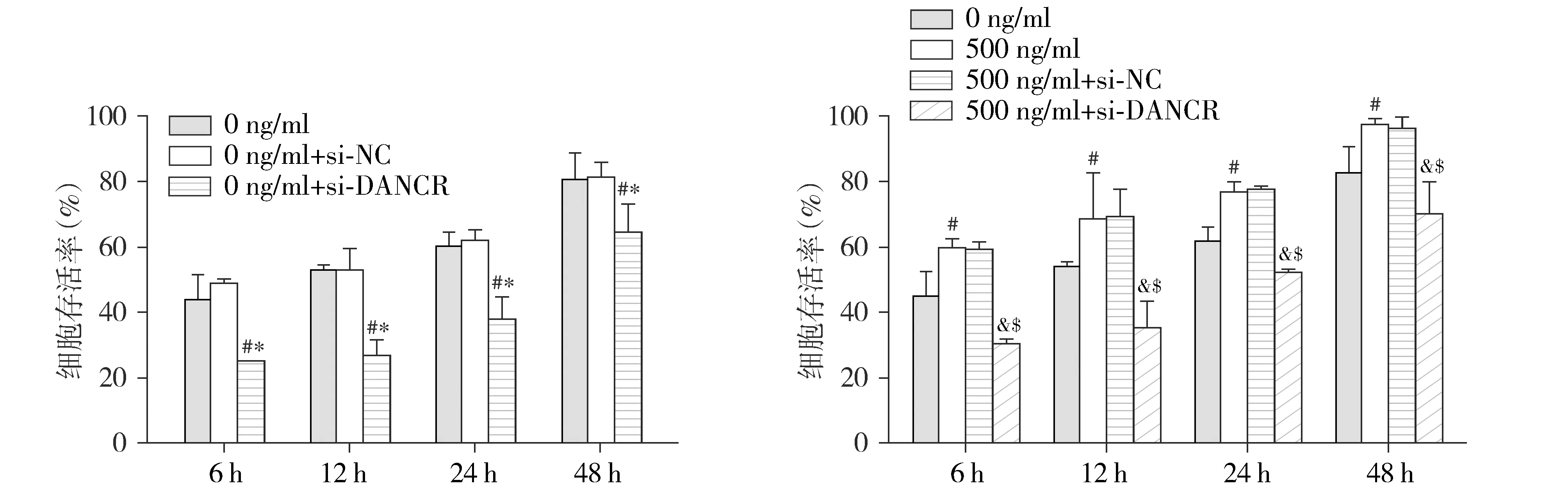
2.4 沉默lncRNA DANCR对尼古丁诱导的NSCLC细胞增殖活性的影响
进一步探究单独沉默DANCR或尼古丁联合沉默DANCR作用下细胞增殖活性的变化,结果显示,与0 ng/ml组或0 ng/ml+si-NC组相比,0 ng/ml+si-DANCR组细胞的增殖活性在各时间点(6,12,24,48 h)均显著降低,差异有统计学意义(P<0.05,见图4A);而0 ng/ml组和0 ng/ml+si-NC组细胞的增殖活性无显著性差异(P>0.05,见图4A)。此外,与0 ng/ml尼古丁组相比,由500 ng/ml尼古丁诱导的NSCLC细胞增殖活性增加在各时间点(6,12,24,48 h)均可被si-DANCR显著抑制,差异有统计学意义(P<0.05,见图4B)。本部分结果表明,沉默DANCR对尼古丁诱导的NSCLC增殖活性具有抑制作用。
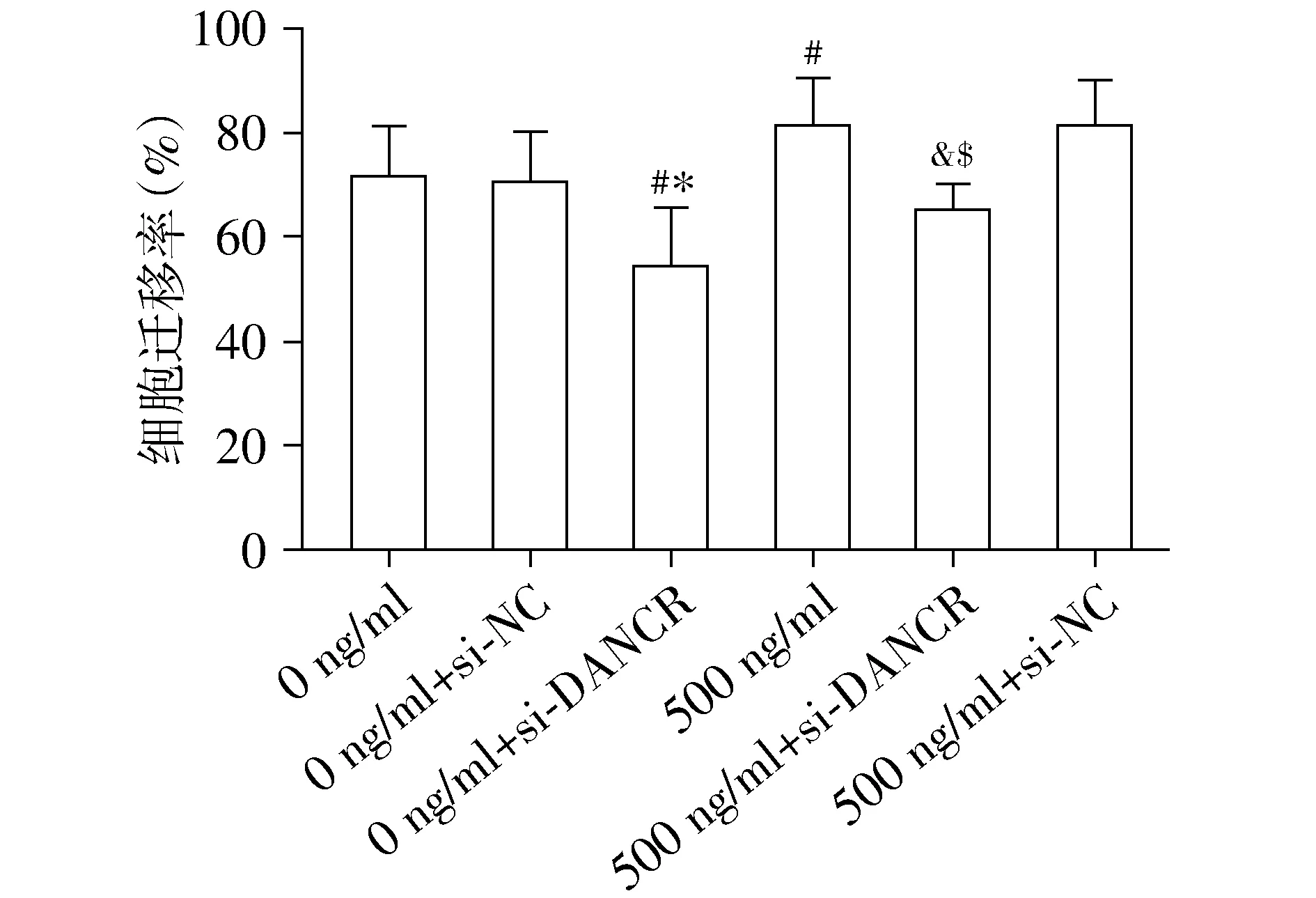
2.5 沉默lncRNA DANCR对尼古丁诱导的NSCLC细胞迁移能力的影响
进一步探索了尼古丁联合DANCR慢病毒沉默对NSCLC细胞迁移的影响,结果见图5。分别与0 ng/ml组和0 ng/ml组+si-NC组相比,0 ng/ml+si-DANCR组细胞的迁移能力显著下降,差异有统计学意义(P<0.05);而0 ng/ml组和0 ng/ml+si-NC组之间,细胞的迁移能力差异无统计学意义(P>0.05)。同时,与0 ng/ml尼古丁组相比,由500 ng/ml尼古丁引起的NSCLC细胞迁移能力增加被si-DANCR显著抑制,差异有统计学意义(P<0.05);而500 ng/ml组和500 ng/ml+si-NC组相比,差异无统计学意义(P>0.05,见图5)。该部分结果表明,沉默DANCR对尼古丁诱导的NSCLC迁移能力增加具有抑制作用。
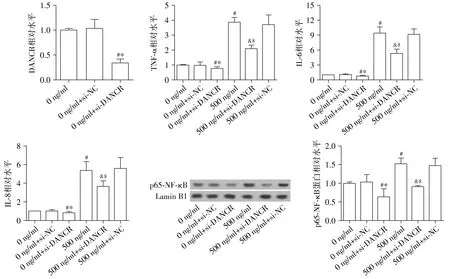
与0 ng/ml组相比,#P<0.05;与0 ng/ml+si-NC组相比,*P<0.05;与500 ng/ml组相比,&P<0.05;与500 ng/ml+si-NC组相比,$P<0.05
F处理因素=64.01,P<0.000 1;F时间=76.95,P<0.000 1 F处理因素=59.99,P<0.000 1;F时间=85.04,P<0.000 1
与0 ng/ml组相比,#P<0.05;与0 ng/ml+si-NC组相比,*P<0.05;与500 ng/ml组相比,&P<0.05;与500 ng/ml+si-NC组相比,$P<0.05
3 讨论
CRI在NSCLC的恶性转化中发挥着关键作用,可介导EMT并促进癌症侵袭[16]。本研究选用人NSCLC细胞系A549,以尼古丁诱导其细胞炎症反应,模拟细胞炎症微环境。首先分别使用浓度为50,250,500 ng/ml的尼古丁刺激细胞,结果显示促炎症因子TNF-α、IL-6、IL-8分泌水平呈现剂量依赖性增加,说明尼古丁诱导了NSCLC细胞的炎症反应。研究认为NF-κB炎症信号通路参与肺癌的转移[5],以往研究中已经明确NF-κB炎症通路的激活是通过IKKβ蛋白将胞质IκBα磷酸化从而导致p65与p50二甲基组成移位到细胞核与特异性κB序列结合,诱导转录相关蛋白[17]。本研究中,NSCLC在尼古丁刺激下,其IKKβ mRNA表达升高,提示细胞NF-κB信号通路被增强。进一步研究证实,尼古丁导致NSCLC细胞内p-IκBα水平和核内p65-NF-κB水平均上调,说明尼古丁激活了NF-κB炎症信号通路。以上研究表明,尼古丁刺激可模拟炎症微环境发生。
近年来,大量临床和基础医学研究聚焦于lncRNA。在NSCLC相关研究中,lncRNA在细胞增殖、凋亡、转移、血管生成、EMT、肿瘤干细胞等肿瘤发展过程中均发挥着关键的作用[18,19]。lncRNA具有高度保守性,能够通过调控基因转录因子、结合靶基因,或扮演内源竞争性RNA(ceRNA),修饰基因甲基化等方式扮演抑癌或者促癌的作用[20]。lncRNA DANCR是新发现的一个促癌基因,在NSCLC中已被研究。研究发现lncRNA DANCR在NSCLC组织和细胞系中表达上调,而且具有促进NSCLC细胞增殖、迁移、侵袭等作用[9]。本研究中,沉默DANCR可导致A549细胞的增殖、迁移能力被抑制,与以往研究结果相符。
已有研究证明尼古丁与肺癌的发生具有密切相关性[11],并具有促进NSCLC细胞增殖、EMT,抑制细胞凋亡,诱导侵袭和迁移的功能[21,22]。本研究同样证实了尼古丁对NSCLC具有明显的促增殖和迁移能力。更重要的是,本研究中,尼古丁诱导的细胞增殖活性可以被沉默DANCR条件下抑制。同时,沉默DANCR显著抑制因尼古丁诱导的NSCLC细胞迁移。DANCR对于尼古丁诱导的NSCLC细胞增殖和迁移能力的抑制作用可能与NSCLC的CRI密切相关。已有研究指出,lncRNA DANCR在肺炎病理组织中诱导炎症因子的表达,介导免疫环境的发生[10]。本研究结果中,沉默DANCR不仅抑制正常NSCLC细胞中的TNF-α、IL-6、IL-8分泌水平和NF-κB炎症信号,而且抑制尼古丁诱导下TNF-α、IL-6、IL-8分泌水平和NF-κB炎症信号。表明lncRNA DANCR通过调控NF-κB炎症信号介导尼古丁诱导的炎症反应,参与调节尼古丁诱导的NSCLC增殖与迁移。
综上所述,lncRNA DANCR作为一关键促癌基因,可通过调节NF-κB炎症通路介导NSCLC细胞的炎症微环境,调控NSCLC细胞增殖和迁移。本研究后续将深入探索lncRNA DANCR对于NSCLC细胞增殖和迁移影响的下游靶基因,进一步明确DANCR介导炎症微环境调控NSCLC细胞恶性发展的分子机制。总之,lncRNA DANCR是关键的炎症微环境介导NSCLC发展机制的潜在研究靶点。
