夜蛾黑卵蜂寄生草地贪夜蛾和斜纹夜蛾卵的生物学特性
2019-11-27霍梁霄周金成宁素芳
霍梁霄 周金成 宁素芳

摘要 2019年7月,我们在贵州黔东南地区采集的草地贪夜蛾Spodoptera frugiperda卵块内发现1种卵寄生蜂,经分子鉴定确认为夜蛾黑卵蜂Telenomus remus。为验证该蜂对草地贪夜蛾卵的寄生效能,本研究以草地贪夜蛾卵为主要研究材料,以斜纹夜蛾Spodoptera litura卵为对照,统计了夜蛾黑卵蜂对这两种寄主卵的卵粒寄生率、单卵出蜂数、雄性比和个体大小等指标。结果显示,夜蛾黑卵蜂对草地贪夜蛾卵和斜纹夜蛾卵的卵粒寄生率均可达到95%以上。与斜纹夜蛾卵相比,草地贪夜蛾卵育出的子代蜂个体较大,但单卵出蜂数相对较低。结果说明夜蛾黑卵蜂是草地贪夜蛾潜在的高效生防作用物,斜纹夜蛾卵可以作为替代寄主用于繁育夜蛾黑卵蜂。本研究将为我国利用夜蛾黑卵蜂防治草地贪夜蛾提供重要参考。
关键词 夜蛾黑卵蜂; 草地贪夜蛾; 卵寄生蜂; 入侵生物; 生物防治
中图分类号: S 476.3, S 433.4 文献标识码: A DOI: 10.16688/j.zwbh.2019406
Abstract In July 2019, we found a species of egg parasitoid wasp emerged from Spodoptera frugiperda eggs collected in southeast of Guizhou province. This parasitoid species was identified by molecular method as Telenomus remus. To test the parasitic efficacy of T.remus against S.frugiperda and S.litura eggs, we investigated the parasitic rates, offspring number per host eggs, body size, and male ratio of T.remus on S.frugiperda or S.litura eggs. The results showed that the parasitic rates of eggs reached at least 95% regardless of S.frugiperda or S.litura eggs. T.remus offspring emerged from S.frugiperda eggs showed larger body size and lower offspring number per host egg than those emerged from S.litura eggs. Our results revealed that T.remus is a potentially effective biological control agent against S.frugiperda, and S.litura eggs are the potential alternative host for the mass rearing of T.remus. This study will supply a primary reference for the application of T.remus against S.frugiperda.
Key words Telenomus remus; Spodoptera frugiperda; egg parasitoids; invasive species; biological control
草地貪夜蛾Spodoptera frugiperda是一种原产于美洲地区的多食性和迁飞性害虫[1]。该虫于2019年1月被报道首次入侵我国云南,随后凭借我国夏季季风迅速北上迁飞扩散[2-3]。截至8月9日,草地贪夜蛾已入侵我国22个省份[4],并跨海迁飞至韩国的济州岛和全罗南道[5]以及日本的冲绳县、鹿儿岛、宫崎县、川崎县和熊本县等地区[6]。草地贪夜蛾在我国及周边国家的入侵性创造了空前的记录,目前已成为东亚地区的头号危险性入侵害虫。
一般而言,当外来生物初次被发现时,使用化学防治等应急防控手段短时间内迅速扑灭入侵生物种群被认为是有效的。然而,草地贪夜蛾具有极强的迁飞扩散能力,其入侵速度和为害面积远超人们的预料,仅依靠应急防控措施已难以在短时间内完全扑杀入侵种群[7-8]。随着草地贪夜蛾在我国的定殖,讨论和建立针对草地贪夜蛾的长效防控策略将成为未来草地贪夜蛾防控工作的重要议题。
天敌等生态抑制因子的缺失是外来物种在入侵地暴发成灾的重要原因之一。筛选和利用专食性天敌可以重新恢复针对外来入侵物种的生态平衡,因而是国内外防控入侵害虫的“模式方法”[8]。草地贪夜蛾具有远距离迁飞的特性,迁飞成虫产下的大量卵块是其在田间为害的源头。因此,寻找草地贪夜蛾卵期的专食性天敌对草地贪夜蛾的长期防控尤为重要。
在草地贪夜蛾的原产地美洲,夜蛾黑卵蜂Telenomus remus、短管赤眼蜂Trichogramma pretiosum和甲腹茧蜂Chelonus spp.是当地草地贪夜蛾卵期的主要天敌[7]。其中,夜蛾黑卵蜂(图1)在我国亦有分布,其在田间主要寄生与草地贪夜蛾同属灰翅夜蛾属的斜纹夜蛾S.litura和甜菜夜蛾S.exigua等鳞翅目害虫的卵[9-11]。我国分布的夜蛾黑卵蜂种群可能是一种针对草地贪夜蛾卵期的潜在生防作用物。近期,我们从贵州田间采集的草地贪夜蛾卵块内成功育出了一种卵寄生蜂,经分子鉴定确认为夜蛾黑卵蜂。为验证夜蛾黑卵蜂对草地贪夜蛾的寄生效能,田间采回的夜蛾黑卵蜂雌蜂被分别供以草地贪夜蛾卵和斜纹夜蛾卵进行寄生,调查并统计了夜蛾黑卵蜂对这两种寄主卵的寄生率、单卵出蜂数、子代蜂个体大小(以后足胫节长度表示)和雄性比。研究结果将为该蜂的大规模繁育和田间应用提供基础性数据。
1 材料与方法
1.1 供试寄主
斜纹夜蛾室内种群由南京农业大学农作物生物灾害综合治理教育部重点实验室提供。草地贪夜蛾室内种群最初采集自贵州省黔东南州凯里市丹寨县附近的玉米田。这两种夜蛾幼虫在室内均以同一种人工饲料饲养至化蛹。人工饲料的配制以Bowling于1967年[12]提出的人工饲料配方为基础,加入微量复合维生素、韦氏盐和甲醛加以改进。待成虫羽化后分别置于圆柱形养虫笼(直径20 cm,高35 cm)内,笼壁覆牛皮纸作为两种夜蛾成虫的产卵介质。以无菌脱脂棉蘸取10%蜂蜜水每12 h饲喂成虫1次。每日将牛皮纸上所产的草地贪夜蛾或斜纹夜蛾卵块剪下,作为供试寄主卵块。斜纹夜蛾和草地贪夜蛾的繁育条件均为(26±1)℃,RH 60%~80%,光周期L∥D=16 h∥8 h。
1.2 夜蛾黑卵蜂的采集和分子鉴定
本试验中的夜蛾黑卵蜂种群由采自贵州省黔东南州凯里市丹寨县附近的玉米田内的4块草地贪夜蛾卵块中育出,共羽化出约300头卵寄生蜂。从每一卵块羽化的寄生蜂中分别随机选取1头蜂单头提取DNA作为模板,利用线粒体COⅠ基因的通用引物对DNA模板进行PCR扩增。扩增产物测序后在NCBI GenBank中对所得序列进行比对,检测其与数据库中已有寄生蜂序列的相似性,鉴定各卵块育出的寄生蜂是否均为夜蛾黑卵蜂。羽化后的夜蛾黑卵蜂成蜂以10%蜂蜜水每日饲喂。试验统一选取2日龄已交配的雌蜂作为供试雌蜂。夜蛾黑卵蜂的繁育条件为(25±1)℃,RH 70%,光周期L∥D=16 h∥8 h。
1.3 寄生试验
将24 h内产出的斜纹夜蛾或草地贪夜蛾卵块剪下后单个卵块分装于玻璃指形管内,然后每卵块引入5头夜蛾黑卵蜂雌蜂,寄生24 h后移出。每种寄主卵进行6次重复。为统计卵块上的总卵粒数,寄生后的卵块在体视显微镜下用镊子小心剥除卵表的鳞毛以方便对卵粒计数(图1)。之后每12 h观察1次卵块,用小毛笔轻轻刷掉卵块上孵化的斜纹夜蛾或草地贪夜蛾幼虫,以防止幼虫取食卵粒。记录孵出幼虫的卵粒数。卵块继续培养至卵粒变黑,表明卵粒被成功寄生,记录寄生卵粒数。待子代蜂羽化后,通过观察子代蜂个体触角末端是否膨大来区分子代个体的性别(触角末端膨大的为雌性;图1),记录卵块上的羽化蜂数和雄蜂比例。死亡后的雌蜂转移至体视显微镜下测量其后足胫节长度,以其表示个体大小。
1.4 数据处理
采用t检验分析夜蛾黑卵蜂寄生两种寄主卵块的卵粒寄生率、单卵出蜂数、个体大小和雄性比的差异。寄生率和雄性比等比例数据预先进行反正弦转换,以满足数据分析的正态性假定。本试验所得数据均使用R统计软件进行分析[13]。
2 结果与分析
2.1 夜蛾黑卵蜂的分子鉴定
本试验所检测的所有卵寄生蜂个体的线粒体COⅠ序列均完全一致。在NCBI GenBank中比对后,发现该序列与夜蛾黑卵蜂COⅠ序列相似度达到99%以上,其中与南非(序列编号MH681663.1)、肯尼亚(MK533758.1)和巴基斯坦(KY835081.1)的夜蛾黑卵蜂COⅠ序列相似度均为99.63%,与美国佛罗里达州(KM485690.1)的夜蛾黑卵蜂序列相似度为99.56%。通过形态观察,我们发现该蜂的形态特征与唐雅丽等对夜蛾黑卵蜂的形态描述基本一致[10]。
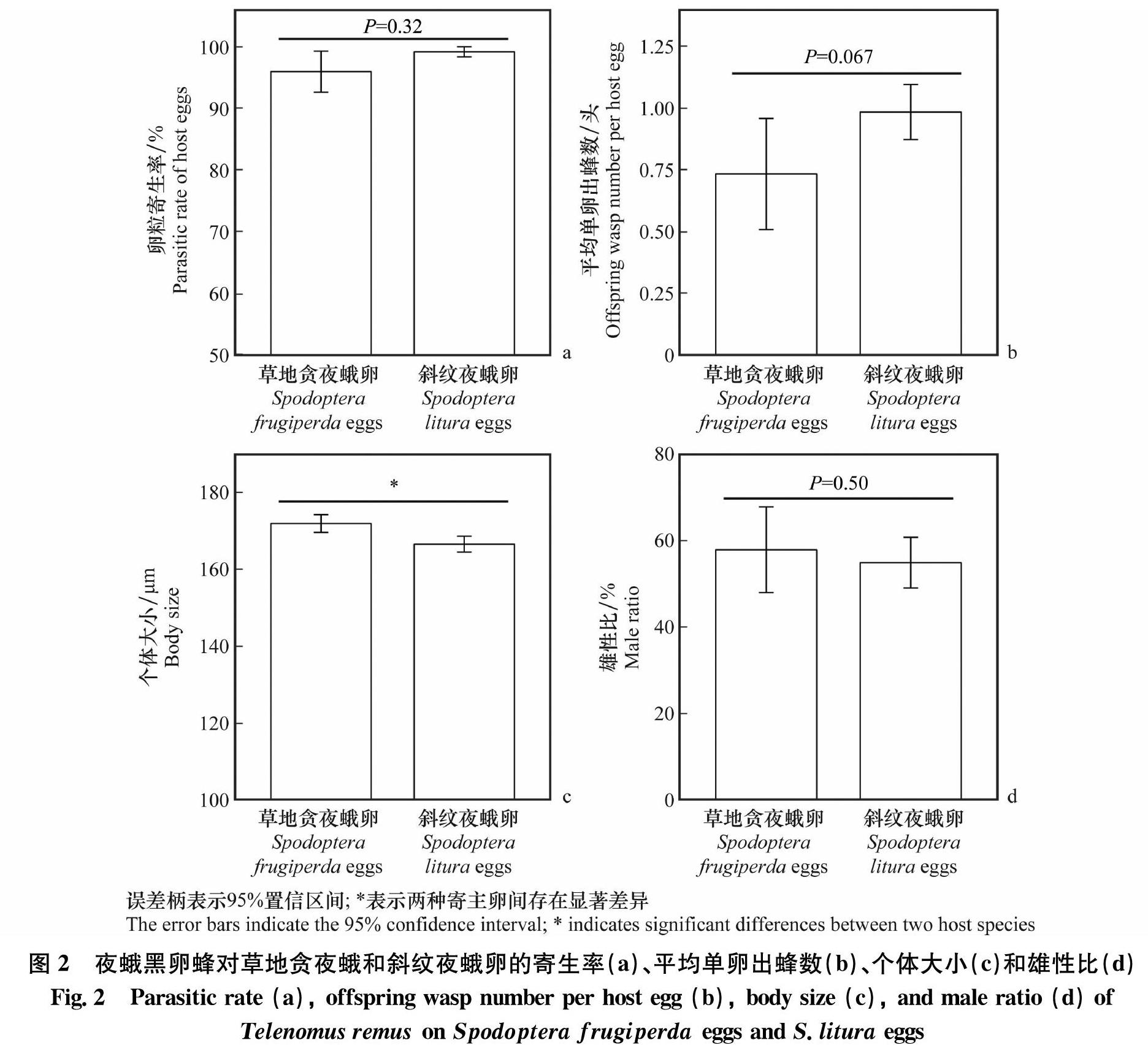
2.2 夜蛾黑卵蜂寄生草地貪夜蛾卵和斜纹夜蛾卵的生物学特性 夜蛾黑卵蜂对草地贪夜蛾卵块和斜纹夜蛾卵块的卵粒寄生率无显著差异(t=5.77,P=0.32),且均在95%以上,分别为97.22%±1.98%和99.26%±0.55%(图2a)。尽管草地贪夜蛾卵块和斜纹夜蛾卵块的单卵出蜂数差异未达到显著水平(t=2.14,P=0.067),但草地贪夜蛾卵块的平均单卵出蜂数(0.73头±0.12头)要低于斜纹夜蛾卵块(0.99头±0.057头)(图2b)。草地贪夜蛾卵育出的子代蜂个体大小(172.01 μm±1.28 μm)显著大于(t=3.20,P<0.05)斜纹夜蛾育出的子代蜂(166.62 μm±1.15 μm)(图2c)。不同寄主卵育出的黑卵蜂子代雄性比之间无显著差异(t=9.51,P=0.50)。草地贪夜蛾卵和斜纹夜蛾卵育出的黑卵蜂子代雄性比分别为57.89%±5.07%和54.91%±3.00%(图2d)。
3 讨论
研究结果表明,夜蛾黑卵蜂对草地贪夜蛾卵和斜纹夜蛾卵的卵粒寄生率均可达到95%以上,这与国外相关研究结果相似[14]。国外学者发现,单头夜蛾黑卵蜂雌蜂在2 h内对1日龄和2日龄的草地贪夜蛾卵的卵粒寄生率可达到90%以上,对3日龄草地贪夜蛾卵的卵粒寄生率也能维持在60%左右[14]。在巴西,有报道称田间人工释放的夜蛾黑卵蜂对草地贪夜蛾卵的寄生率可达70%[15]。此外,一些研究发现夜蛾黑卵蜂偏好较高的湿度环境,同时对高温也具有一定的耐受能力[16-17]。当相对湿度在80%时,夜蛾黑卵蜂雌蜂的寿命和寄生能力显著高于相对湿度40%条件下的雌蜂[16]。另有研究发现,在34℃的恒定高温条件下,夜蛾黑卵蜂依然可以完成整个世代的生长发育[17]。我国大部分区域属亚热带或温带季风气候,持续性的高温多雨天气是我国农作物生长季节的主要气候特征,我们推测夜蛾黑卵蜂可能更适应这种高温高湿的田间环境。鉴于夜蛾黑卵蜂的诸多优点,拉美国家的生物防治工作者早已开展了夜蛾黑卵蜂的规模化饲养和田间应用的相关研究,已取得了许多可借鉴的经验[14-17]。在草地贪夜蛾已经入侵我国的形势下,关于夜蛾黑卵蜂的工厂化繁育和田间应用技术的研究将被提上日程。
由于草地贪夜蛾高龄幼虫具有自相残杀习性,室内饲养时需单头分装饲养,费时费力,繁殖成本较高。因此,要想实现夜蛾黑卵蜂的大规模繁育,就需要寻找繁育成本相对较低且容易获得的替代寄主。国外同行利用米蛾Corcyra cephalonica卵繁育夜蛾黑卵蜂并获得了成功[18-20],但我们前期未发表的研究和其他国内同行近期的研究均表明,目前国内以米蛾卵繁育夜蛾黑卵蜂的尝试均不成功[21]。我们推测这可能是由于野外自然寄主育出的夜蛾黑卵蜂在室内无法识别并寄生米蛾卵。国内曾有学者利用甜菜夜蛾S.exigua卵和斜纹夜蛾卵来繁育夜蛾黑卵蜂[10-11]。我们的研究也发现,以斜纹夜蛾卵繁育夜蛾黑卵蜂是可行的。由于斜纹夜蛾的人工饲养方法已相对成熟,以斜纹夜蛾卵作为中间寄主规模化繁育夜蛾黑卵蜂值得进一步探索。
我们的研究还发现,与斜纹夜蛾卵相比,草地贪夜蛾卵育出的子代蜂个体较大,但单卵出蜂数和寄生率相对较低。我们推测夜蛾黑卵蜂寄生斜纹夜蛾卵时可能存在较高概率的“过寄生”现象,即雌蜂可能在一个寄主卵内产下多个子代。虽然过寄生行为有可能会增加单卵出蜂的数量,但在寄主营养资源有限的条件下,同一卵内发育的多个子代可能会受到营养胁迫,进而导致子代个体偏小[22]。另一方面,夜蛾黑卵蜂幼虫具有发达的上颚。当多个子代在同一寄主卵内发育时,孵化的幼虫在卵内相遇时会利用其上颚互相撕咬[23]。这种子代幼虫个体间的相互争斗有可能导致幼虫“受伤”,进而对其生长发育产生不利影响。
与其他类似研究[24]不同的是,本试验的夜蛾黑卵蜂子代雄性比偏雄。一般而言,夜蛾黑卵蜂等聚寄生性寄生蜂的子代性别比例以雌性偏多,这与其在自然界中的交配模式有关。小型膜翅目昆虫扩散能力相对较弱,其通常在1个寄主或1个卵块内产下一窝子代。这些昆虫的雄性子代和雌性子代在羽化后会在同一窝内进行近亲交配[25]。为了最大限度地扩张其种群数量并避免子代雄性交配时发生竞争,母代雌蜂通常会在1窝子代中仅分配少量的雄性。然而,当多个雌蜂同时存在于同一卵块时,根据哈密尔顿的局部配偶竞争原理(local mate competition,LMC),雌蜂又会根据后代的数量适当增加雄性后代的比例,这种性别分配行为普遍存在于黑卵蜂等聚集性寄生蜂中[25-26]。在本试验中,由于我们在同一个卵块上接入了5头雌蜂对其寄生,较多的母代雌蜂数量可能是后代雄性比例较高的原因。国外同行的类似研究也发现,夜蛾黑卵蜂的子代性别分配行为严格遵循哈密尔顿性别分配模型,当“建群雌蜂”较多时,其子代雄性比例明显升高[26-27]。在该蜂的工厂化繁育过程中,增加具有寄生功能的子代雌蜂比例对于提高其田间生物防治效能是有益的。因此,在该蜂的繁育过程中应注意保持较低的蜂卵比,避免后代雄性比例过高。
我们的研究及国外其他研究均证明,夜蛾黑卵蜂很可能是防治草地贪夜蛾的有力武器。由于草地贪夜蛾可通过远距离扩散摆脱当地天敌的自然控制作用,仅依靠小规模释放天敌,通过其在田间的缓慢定殖来实现对目标害虫的防控可能收效不大。对于迁飞性害虫而言,应当坚持统防统治的原则,考虑采取增强型生物防治策略。当迁飞成虫于田间大量产卵时,可通过淹没式释放措施,在田间释放大量卵寄生蜂以达到预期的防治效果。考虑到草地贪夜蛾已经在各地为害,目前急需建立夜蛾黑卵蜂大规模繁育技术方法,以实现该蜂在田间的大规模应用。
参考文献
[1] TODD E L, POOLE R W. Keys and illustrations for the armyworm moths of the noctuid genus Spodoptera Guenée from the Western Hemisphere [J]. Annals of the Entomological Society of America, 1980, 73(6): 722-738.
[2] 吴秋琳, 姜玉英, 吴孔明. 草地贪夜蛾缅甸虫源迁入中国的路径分析[J]. 植物保護, 2019, 45(2): 1-6.
[3] 姜玉英, 刘杰, 朱晓明. 草地贪夜蛾侵入我国的发生动态和未来趋势分析[J]. 中国植保导刊, 2019, 39(2): 33-35.
[4] 宁夏农技推广总站. 宁夏发现草地贪夜蛾成虫[EB/OL]. 植保情报, 2019年第15期[2019-08-02]. http:∥nynct.nx.gov.cn/xwzx/zwdt/201908/t20190802_1643775.html.
[5] Food and Agriculture Organization of the United Nations. Report of first detection of fall Armyworm (FAW) in Republic of Korea[EB/OL].[2019-06-21]. https:∥www.ippc.int/fr/countries/republicofkorea/pestreports/2019/06/reportoffirstdetectionoffallarmywormfawinrepublicofkorea/.
[6] 高知県病害虫防除所.令和元年8月号[EB/OL].[2019-08-01]. http:∥www.nogyo.tosa.pref.kochi.lg.jp/info/dtl.php?ID=8415.
[7] 唐璞, 王知知, 吴琼, 等. 草地贪夜蛾的天敌资源及其生物防治中的应用[J]. 应用昆虫学报, 2019, 56(3): 370-381.
[8] 万方浩, 侯有明, 蒋明星. 入侵生物学[M]. 北京:科学出版社, 2015.
[9] 李志刚, 吕欣, 押玉柯, 等. 粤港两地田间发现夜蛾黑卵蜂与螟黄赤眼蜂寄生草地贪夜蛾[J/OL]. 环境昆虫学报,2019,41(4):760-765.
[10] 唐雅丽, 陈科伟, 许再福. 夜蛾黑卵蜂(Telenomus remus Nixon)个体发育研究[J]. 长江蔬菜, 2010(18): 1-3.
[11] 杨莹,韩勇,方祝红,等.寄主卵龄和接触时间对夜蛾黑卵蜂寄生能力的影响[J].应用昆虫学报,2012,49(6):1490-1495.
[12] BOWLING C C. Rearing of two lepidopterous pests of rice on a common artificial diet [J]. Annals of the Entomological Society of America, 1967, 60(6): 1215-1216.
[13] R Core Team. R: A language and environment for statistical computing.R foundation for statistical computing, vienna, austria [EB/OL].[2019-03-11]. https:∥www.rproject.org/.
[14] PEAFLOR M F G V, SARMENTO M M M, BEZERRA DA SILVA C S, et al. Effect of host egg age on preference, development and arrestment of Telenomus remus (Hymenoptera: Scelionidae) [J].European Journal of Entomology,2013,109(1): 15-20.
[15] QUEIROZ A P, DE FREITAS BUENO A, POMARIFERNANDES A, et al. Influence of host preference, mating, and release density on the parasitism of Telenomus remus (Nixon) (Hymenoptera, Platygastridae)[J]. Revista Brasileira de Entomologia, 2017, 61(1): 86-90.
[16] POMARIFERNANDES A, QUEIROZ A P, DE FREITAS BUENO A, et al. The importance of relative humidity for Telenomus remus (Hymenoptera: Platygastridae) parasitism and development on Corcyra cephalonica (Lepidoptera: Pyralidae) and Spodoptera frugiperda (Lepidoptera: Noctuidae) eggs[J]. Annals of the Entomological Society of America, 2015, 108(1): 11-17.
[17] POMARI A F, DE FREITAS BUENO A, DE FREITAS BUENO R C O, et al. Biological characteristics and thermal requirements of the biological control agent Telenomus remus (Hymenoptera: Platygastridae) reared on eggs of different species of the Genus Spodoptera (Lepidoptera: Noctuidae)[J]. Annals of the Entomological Society of America, 2012, 105(1): 73-81.
[18] POMARIFERNANDES A, DE FREITAS BUENO A, DE BORTOLI S A. Size and flight ability of Telenomus remus parasitoids reared on eggs of the factitious host Corcyra cephalonica [J]. Revista Brasileira de Entomologia, 2016, 60(2): 177-181.
[19] QUEIROZ A P, DE FREITAS BUENO A, POMARIFERNANDES A, et al. Quality control of Telenomus remus (Hymenoptera: Platygastridae) reared on the factitious host Corcyra cephalonica (Lepidoptera: Pyralidae) for successive generations [J].Bulletin of Entomological Research,2017,107(6): 791-798.
[20] QUEIROZ A P, DE FREITAS BUENO A, POMARIFERNANDES A, et al. Low temperature storage of Telenomus remus (Nixon) (Hymenoptera: Platygastridae) and its factitious host Corcyra cephalonica (Stainton) (Lepidoptera: Pyralidae)[J]. Neotropical Entomology, 2016, 46(2): 1-11.
[21] 戴鵬, 孙佳伟, 陈永明, 等. 赞比亚发现三种防治草地贪夜蛾的卵寄生蜂简报[J/OL]. 吉林农业大学学报:1-6[2019-08-12].https:∥doi.org/10.13327/j.jjlau.2019.5310.
[22] GANDON S, RIVERO A, VARALDI J. Superparasitism evolution: Adaptation or manipulation?[J]. The American Naturalist, 2006, 167(1): 1-22.
[23] DAN G. The developmental biology of Telenomus remus Nixon (Hym. Scelionidae)[J]. Bulletin of Entomological Research, 1972, 61(3): 385-388.
[24] SCHWARTZ A, GERLING D. Adult biology of Telenomus remus[Hymenoptera: Scelionidae], under laboratory conditions [J]. Entomophaga, 1974, 19(4): 483-492.
[25] WERREN J H. Sex ratio adaptations to local mate competition in a parasitic wasp [J]. Science, 1980, 208(4448): 1157-1159.
[26] WELZEN C R L V, WAAGE J K. Adaptive responses to local mate competition by the parasitoid Telenomus remus[J]. Behavioral Ecology and Sociobiology, 1987, 21(6): 359-365.
[27] CARNEIRO T R, FERNANDES O A, CRUZ I. Influence of females intraspecific competition and lack of host on Telenomus remus Nixon (Hymenoptera, Scelionidae) parasitism on Spodoptera frugiperda (J. E. Smith) (Lepidoptera, Noctuidae) eggs[J]. Revista Brasileira de Entomologia, 2009, 53(3): 482-486.
(责任编辑: 杨明丽)
