水流对草鱼幼鱼趋光行为的影响
2019-11-26林晨宇许家炜施家月白艳勤尹入成石小涛
张 宁 林晨宇 许家炜 施家月 罗 佳 白艳勤 尹入成 石小涛
(1. 三峡大学三峡库区生态环境教育部工程研究中心, 宜昌 443002; 2. 河海大学水利水电学院, 南京 210098;3. 中国电建集团华东勘测设计研究院有限公司, 杭州 310014)
光诱驱鱼技术被认为是潜在的鱼类无伤定向驱导技术之一[1—3], 在协助鱼类过坝等实践中被证明有较好的应用前景。具体而言, 其可被应用于鱼道进口、过鱼旁路系统及下行通道等位置以吸引或诱导鱼群[4,5]。而在栖息地修复中, 该技术可用于营造更适于鱼类生存的光环境, 提高鱼类三场利用率和栖息地适宜度[6—8]。因此, 充分了解目标鱼类对光环境的行为响应显得极为重要。
如同其他环境因子, 光照对鱼类的影响体现在诸多方面, 如摄食、生长、繁殖、躲避敌害等[9—12]。例如, 鲻(Mugil cephalus)幼鱼的摄食率在102lx时最高[13], 真鲷(Pagrosomus major)在50—500 lx的光照强度范围内的摄食强度最高[14]。巴丁鱼(Pangasias sutchi)幼鱼生长的最佳光照强度为0.1 lx[15], 而使用人造光改变斑马鱼(Danio rerio)所处的光环境则会抑制其的繁殖行为[16]。此外, 芬兰学者Ana Carolina Luchiari 等[17]还发现, 梭鲈(Lucioperca lucioperca)在低光强情况下, 具有更强的摄食和躲避敌害能力。而在有关诱驱鱼技术和光环境营造的报道当中, 由于鱼类的趋光性行为在工程中具有更实用的价值, 故其受到了更多的关注。鱼类趋光性是指鱼对光刺激产生的定向运动的特性。其中, 光趋性行为根据鱼类的偏好与回避程度又可分为正趋光性和负趋光性[18—21]。大西洋鲟(Acipenser sturio)和中华鲟(Acipenser sinensis)幼鱼是已知典型的正趋光性鱼类[22,23]。相比之下, 兰圆鲹(Decapterus maruadsi)正趋光性较弱[24], 而研究发现孔雀鱼(Poecilia reticulata)幼苗的趋光率会随着光源照度的增加呈先升后降的趋势, 表明其具有最适趋性照度[25]。一些鱼类, 如中吻鲟(Acipenser medirostris),在整个生活史中都无明显趋性, 既不趋光, 也不避光[23]。最后, 鱼类光趋性还与生长阶段有关, 如史氏鲟(Acipenser schrenckii)在光照强度为250—1500 lx时, 其趋光性随鱼体生长先增加后减少[26,27]。
除光环境外, 水流被认为是天然河流中影响鱼类行为的主导因素, 它通过刺激鱼类侧线, 使其产生相应的反应行为及游泳模式[2,28]。张硕等[29]对黑鲪(Sebastodes fuscescens)幼鱼的趋流性进行探究,初步描绘了该鱼种对水流的偏好区间和耐受能力。而进一步研究表明, 大多数的鱼类都具有趋流性, 且适当流速范围的水流就会对鱼类产生一定的诱集效果[19]。在实际过鱼工程诱鱼水流设计当中,感应流速作为鱼能够辨别水流方向的最小流速, 往往被设定为诱鱼水流的最低阈值[19,30—32]。在天然河流环境中, 水流和光照对鱼类的作用同时存在并相互影响。国外有研究表明, 适当光照可增加鲑(Salmo salar)幼鱼应对加速流时的回避行为[33]。亦有研究指出, 相比于大型无脊椎动物, 低光照环境下多数鱼类在鱼道水流中的上溯积极性显著下降[34]。在国内, 邓雅敏等[35]对光环境与流速对鱼类行为影响这方面进行了探究, 但仅限于在鱼礁场景中应用。罗佳等[1]研究显示在特定的流速条件下, 闪光可以对光倒刺鲃(Spinibarbus hollandi)产生吸引作用。当前, 如何定量鱼类在两种因素共存条件下的响应行为并探究其内在机理已成为研究重点, 而相关工作在国内外均乏善可陈。
为对该问题展开深入探讨, 本研究选择草鱼(Ctenpharyngodon idellus)作为实验对象。草鱼作为我国四大家鱼之一, 具有典型的江湖洄游特性。由于基本生活于水体中下层, 导致其杆状视觉细胞逐渐退化, 表现出一定的负趋光性[36], 同时对水流流态也较为敏感[37]。基于这一物种特性, 本研究采用室内行为学实验方法, 分析感应流速对草鱼在特定光强照度下的趋避行为, 为相关工程应用提供基础数据参考。
1 材料与方法
1.1 实验材料
实验用鱼由湖北枝江某养殖场提供, 共计200尾, 平均体长(12.26±1.01) cm, 平均体重(36.73±7.03) g, 暂养于三峡大学生态水利学实验室。暂养缸规格均为直径2.0 m、高0.5 m、水深0.3 m的圆形蓝色玻璃纤维水缸。暂养用水为曝气后的自来水, 每2天更换池水的1/2—1/3。暂养条件采用自然光周期(0.002—1500 lx), 水温设置18℃, 保持水中溶氧含量> 7.0 mg/L, pH为7.1—7.3。暂养7d, 期间每天投喂2次(7:30和18:30), 待其可正常进食游动并禁食48h后才可用于实验。随机挑选健康且体长均等的草鱼作为实验对象, 为防止对环境产生适应性,实验用鱼均不重复使用。待实验结束后, 将实验鱼放入另一暂养缸中进行养殖。
1.2 实验装置
实验采用自制的大型开放式自循环鱼类游泳水槽(图 1), 尺寸为250 cm×90 cm×60 cm(长×宽×高), 试验区长为200 cm。水槽主体由砖混结构搭建, 实验用水采用可调流量循环水。光源由1根220 V,45 W的可调LED灯管提供, 布置于实验区域前方上端。监控系统由显示器、录像机并连接1个SNUA3160GP(大华)红外摄像仪组成。摄像仪置于实验区上方3 m处, 通过录像视频对实验区域进行观察和记录实验数据。
1.3 实验方法
实验在光源照度为300 lx的环境下进行, 根据草鱼感应流速0.12 m/s[32], 设有3种不同的流速工况(0、0.1和0.2 m/s), 并设置有黑暗静水的对照组。每次实验同时放入5尾实验鱼, 每组工况需进行10次重复。实验用水均为小型水库循环水(不需再做曝气处理), 实验前检查水槽中的水温是否与暂养水温一致并控制水深为0.1 m, 而后在暂养水缸中随机捞取5尾幼鱼放入水槽实验区内进行适应。黑暗适应30min后, 按工况要求调节流速, 同时在2min内均匀提高光源照度至实验处理值, 并开启红外摄像仪, 正式开始实验, 每组实验时长为60min。为避免自然光的影响, 实验时间为当日20:00至次日06:00。在实验结束后, 对所获取的视频数据进行处理分析。每隔10s统计一次实验鱼在实验区内的分布情况和头部朝向, 并记录整个区域内的光照度。
所记录的数据用于描述草鱼幼鱼在不同工况下对光环境的选择, 趋流性及总体偏好情况。
(1)以光强期望值F为指标表示草鱼在不同工况下对光照度的平均选择。

式中:x为实验鱼至光源距离;I(x)为光衰减函数;D(x)为实验组鱼沿实验区长度方向的分布概率函数。

图1 装置示意图Fig. 1 Top view of experimental apparatus
(2)趋流性主要考察实验鱼头部朝向分布率H,该指标表示草鱼幼鱼头部运动方向与水流方向夹角(180°时为“顶流行为”, 0°时为“顺流而下”)在各角度范围内的分布概率。

式中:i-j为特定工况下实验鱼鱼头与水流方向夹角所处的角度区间;ni-j表示所成夹角落在i-j区间内实验鱼的分布尾数,N表示实验鱼在特定工况下的总尾数。
(3)为排除鱼类在实验水槽中自由游泳本底值的影响, 采用环境偏好指数P为指标表示实验组草鱼对实验区不同位置的真实偏好情况。

式中:C(x)为对照组实验鱼沿实验区度方向的分布概率函数;P>0即表示喜好,P=0时表示没有偏好,P<0则表示回避,P的绝对值大小表示对该区域的偏好强烈程度。
1.4 数据处理
本实验所有的数据都使用Origin Pro 2017软件进行分析和处理, 采用回归分析拟合多项式方程,对比在同一照度下水流流速逐渐增大时鱼群的分布及行为变化规律, 得到相关数据结果。
2 结果
2.1 头部朝向分布
根据公式(2)可得, 各工况下草鱼头部朝向分布率如图 2所示。其中, 当处于静水工况时, 无论是否有光, 草鱼幼鱼在各方向上的朝向分布都较为均匀,所有分布率均小于10% (图 2a和图 2b)。在有流速的工况下, 实验鱼头部朝向趋近于180°, 即呈现出顶流的游泳姿态。当角度范围在160°—200°时, 分布率均超过10%。此外, 在0.1和0.2 m/s两种流速工况下, 鱼类头部朝向亦存在明显差别。0.1 m/s的流速小于草鱼的感应值, 实验鱼除对逆水流方向选择较多外, 其朝向在其他方向上也有一定分布(图 2c)。反之, 0.2 m/s的流速大于草鱼感应值, 完全激发了鱼类的趋流性, 对游泳方向有很好的约束作用, 在180°—200°内, 分布率接近50% (图 2d)。

图2 不同工况下草鱼头部朝向分布率Fig. 2 Directional heading distribution pattern of experimental fish under different configurations
2.2 光强期望值
当光源照度为300 lx时, 实验区域内的光强衰减情况如图 3所示。
实验鱼各工况下沿长度方向的分布概率如图 4所示。从图中可明显看出, 当在实验区引入光源时,静水工况下, 实验鱼在实验区尾端的分布即明显高于其他位置, 可拟合公式:
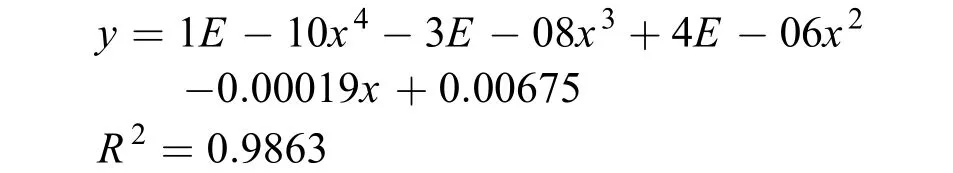
根据公式(1)计算可得此时鱼群的光强期望值为52.45 lx(图 4a)。当照度不变, 流速增大至0.1 m/s时, 草鱼幼鱼在水槽中间部位的分布明显减少, 而在尾部的分布则进一步增加, 对该流速表现出回避行为, 分布曲线拟合公式为:

计算得鱼群的光强期望值为34.62 lx (图 4b),比静水工况降低了17.83 lx。当流速继续增大到0.2 m/s时, 实验鱼在尾部的分布开始减少, 重新开始向前端移动, 从而在水槽前中部的分布增多, 分布曲线拟合公式为:

计算得鱼群的光强期望值重新回升到37.86 lx(图 4c)。

图3 光照度衰减图Fig. 3 Optic attenuation of experiment light source

图4 不同工况下草鱼沿水槽概率分布情况Fig. 4 Potential distribution of experimental fish along test channel under different configurations
2.3 环境偏好指数
根据公式(3)计算得到的草鱼幼鱼在不同流速及不同光照条件下对水槽环境的偏好情况如图 5所示。其中, 图 5a表示黑暗静水工况(对照组)下实验鱼的分布情况, 是偏好指数计算的本底值。在加入光源后, 实验鱼在水槽中的偏好呈现“两头高, 中间低”的分布趋势, 且两端对称, 其中当x<42.75或x>158.41时, 偏好指数为正, 当42.75<x<158.41时偏好指数为负。这表明, 相对于黑暗环境, 草鱼幼鱼并未表现出对某一光强范围的相对偏好也未因负趋性而聚集于实验区尾部, 相反地趋向于向两边“逃离”实验区(图 5b)。在流速为0.1 m/s的情况下,实验鱼总体偏好形式与静水工况下类似, 同时后部与中部鱼的数量差距进一步增大, 其中当x<28.47或x>148.45时, 偏好指数为正, 当28.47<x<148.45时偏好指数为负。这表明, 实验鱼依然保留了静水工况下的行为特性, 而小流速的引入虽在一定程度促使鱼体更多朝向逆水流方向, 但更多地引起鱼类“逆流后退”行为, 未能有效激发其上溯(图 5c)。随着流速继续增大至0.2 m/s, 尽管实验鱼分布大体形式与上述两工况保持一致, 但前后端与中部区域的偏好差距明显缩小, 其中当x<25.90或x>147.04时,偏好指数为正, 当25.90<x<147.04时偏好指数为负,同时流速0.1 m/s时存在的对实验区尾部的聚集水平也基本恢复至静水工况水平。这说明, 趋流性此时已在鱼类行为导向中发挥重要作用, 不但增加了鱼类对实验区中前部的偏好, 还降低了其在水槽两端聚集的情况, 减弱了原有光环境的影响(图 5d)。

图5 不同工况下草鱼对不同光区域的偏好指数Fig. 5 Preference index of experimental fish at different light regimes under given configurations
3 讨论
3.1 草幼鱼对光强照度的选择
光作为普遍存在环境因子, 对水生生物的影响呈多样化趋势。不同种类或相同种类不同龄期水生生物的趋光敏感性有所不同[25,38,39]。草鱼幼鱼被普遍认可为负趋光鱼种, 而天然水域中的大量水生生物, 特别是底栖性鱼类, 也表现出对低照度环境的偏爱。如瓦氏黄颡鱼(Pelteobagrus vachelli)的偏好光照度为0—10 lx; 日本鳗鲡(Anguilla japonica)早期幼苗更偏好于25 lx的弱光; 相类似的, 铠平鲉(Sebastes hubbsi)感受强光的视锥细胞数量较少, 更适合在昏暗的条件下活动[40—42]。李大鹏等[43]的研究也表明, 史氏鲟稚鱼存在避强光的行为, 这也与它本身生活在浑浊的水体环境中有关。在本实验中, 从表面来看, 实验对象对光照度偏好均不低。尤其在静水工况下, 草鱼幼鱼的光强期望值达到了52.45 lx, 但通过观察图 5b可知, 在排除水槽背景影响后, 草鱼实际上偏好聚集于实验区域的首尾两侧, 而在中部游动时间较少, 属于典型的“逃逸”倾向行为(将鱼类相比对照组往水槽边界聚集的行为定义为“逃逸”倾向)。之前计算所得的光强期望值是该现象的平均值, 而事实上在该期望值的鱼类分布较少。在其他两种流速工况下的光强期望值也与静水工况类似。在过往的研究中, 已有学者证明, 一定的光照强度会形成光压力, 激发鱼类的应激反应, 具体行为则表现为对实验装置边壁的趋触性[44,45]。由此, 实验设置的300 lx光源照度已超过草鱼幼鱼的喜好光强值[36], 其表现出的“逃逸”倾向行为在很大程度上是由负趋性所引起的应激反应。故此, 可判定草鱼在实验设置光照处理下依然呈现极大的负趋性。
3.2 趋流性及其对趋光性的影响
趋流性作为鱼类在河流中行为的主要表现形式, 已有大量报道。国外早期研究指出, 一定流速的水流对鱼类会有诱导作用, 但其诱导的效果并非随流速的增大而增大[46]。近年来, 国内相关研究成果也颇多[2,47]。就感应流速而言, 李志敏等[48]和蔡露等[49]均将感应流速作为考察鱼类启动趋流行为的关键指标。本研究统计了鱼类在略低于和略高于感应值流速下的头部朝向分布, 定量判断了鱼类的趋流性。值得注意的是, 当水流流速未达到感应值时, 其微小的水流不但不能有效触发草鱼的趋流性, 反而将其驱离, 成为鱼类前进中的阻碍因素, 致使其光强期望值从52.45 lx降至34.62 lx。这一结果在Lin等[50]的野外集鱼船模型研究中得以充分体现。而在超过感应流速的水流中, 鱼类趋流性受到完全激发, 并成为其主导行为。此时草鱼幼鱼的光强期望值从34.62 lx回升至37.86 lx。
在天然河流环境中, 水流和光环境会对鱼类行为的影响并非是线性。许家炜等的研究发现鱼类的趋光性并没有随着水流流速的增大而增大[51]。邓雅敏等[35]的研究也证明石鲷(Oplegnathus fasciatus)在高流速下的平均聚集率会更大, 且其表现出一定的负趋光性。在本研究中, 随着流速的增大,鱼类的分布逐步受趋流性影响。特别的, 当流速为0.2 m/s时, 分布于实验区两端的鱼类明显减少, 且在各位置呈现均匀分布的趋势, 这表明草鱼幼鱼在实验光环境下的负趋性逐步被趋流性所取代。而不少研究, 也强调了在自然环境, 特别是高流速环境中, 水流是引导鱼类上溯的主导因子[52,53]。
3.3 两种趋性的工程应用
在已有实践中, 可在过鱼或栖息地修复工程中合理利用灯光进行定向驱导, 以达到保护鱼类和促进繁殖的目的。例如, 根据鱼类对一定光照的偏好,调整河流环境中的照度可人为引导鱼类在该照度的水层内趋光集群[25,54]。在本研究中, 根据草鱼的负趋光性, 显然可通过营造低照度光场对指定区域草鱼进行驱离。同时, 根据本研究可知, 在不同流速下草鱼行为的主导环境因素不同。因此, 应根据工程实际情况, 对鱼类行为进行合理预判。在水体中, 随水深的不同, 流速从底层到表层变化范围较大, 且上层水体的流速较下层水体的流速大[2,55]。只有高于感应流速的水流才具有诱鱼效果。微水流不但无法诱鱼, 还会产生负面作用。因此, 在过鱼设施诱鱼水流设计中, 应充分考虑周围流场情况,合理营造感应流场, 从而提高过鱼效率。
总体而言, 在一定的流速和光照刺激下, 草鱼的行为活动均会受到一定的影响, 而这种影响可能呈现此消彼长, 相互制约的趋势。因此, 在后续的研究中, 应继续增加水流工况和光照度梯度, 以更好的得出水流流速与鱼类趋光性之间的关系, 为相关研究和工程应用提出参考及合理的建议。
