淹水胁迫及胁迫解除对甜瓜幼苗呼吸等生理代谢的影响
2019-11-22张永平刁倩楠张文献田守波范红伟
张永平 刁倩楠 张文献 田守波 许 爽 范红伟*
(1 上海市农业科学院园艺研究所,上海市设施园艺技术重点实验室,上海 201403;2 上海市农业技术推广服务中心,上海 201103)
甜瓜(Cumumis meloL.)喜温但不耐热。我国上海及周边地区夏秋季甜瓜播种育苗及移栽在7~8月进行,期间强降雨容易造成土壤渍水,影响甜瓜夏秋季的正常种植(王俊良 等,2013)。由于不同甜瓜品种间抗逆性差异较大,对水分的耐受性不同,不同的生长发育阶段、不同水分胁迫强度及胁迫时间对甜瓜的生长发育、生理生化过程均有不同的影响,因而对甜瓜不同品种的水分逆境适应性进行研究对甜瓜的栽培种植具有重要意义。
据统计,在全球范围内10%的灌溉土地受到洪涝灾害的影响,这可能会降低20%的农作物产量(Shabala,2011)。作物在淹水逆境下,生理生化代谢会产生变化来适应不良环境。研究表明,淹水条件能显著抑制不结球白菜幼苗的有氧呼吸,加强无氧呼吸,最终抑制根系的生长(郭欣欣 等,2015)。淹水胁迫下,植物根系缺氧,无氧呼吸增强,乙醇脱氢酶(alcohol dehydrogenase,ADH)、丙酮酸脱羧酶(pyruvate decarboxylase,PDC)和乳酸脱氢酶(lactate dehydrogenase,LDH)活性提高,促进了植株根系对缺氧环境的适应性(全瑞兰和玉永雄,2015)。淹水后根系干物质积累量、根长密度、伤流速率和根系活力等显著降低,说明淹水胁迫抑制了根系的正常生长发育,从而导致植株地上部的正常生长发育受到抑制(任佰朝 等,2017)。淹水胁迫能诱导细胞产生自由基,进而导致离子渗漏和细胞死亡,高水平的超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)和 过 氧 化氢酶(catalase,CAT)活性在长时间淹水条件下在多种植物的存活中发挥重要作用(陈鹭真 等,2006)。到目前为止,国内外学者对淹水胁迫下植株生长发育、生理生化及分子调控等特征进行了深入研究,但主要集中于水稻(黄万勇 等,2017)、玉米(僧珊珊 等,2012)、辣椒(刘周斌 等,2015)、青花菜(高旭 等,2018)和不结球白菜(王华 等,2013)等作物上,对淹水胁迫下甜瓜生长和生理变化研究相对较少。所以本试验在前期工作基础上选择耐涝性稍强和耐涝性较弱的2个甜瓜品种为材料,通过对幼苗期甜瓜根系生长、呼吸酶代谢和抗氧化酶活性等指标的测定,比较分析不同甜瓜品种对不同程度淹水胁迫的响应差异及其生理原因,以期为进一步培育耐淹的甜瓜品种及其抗逆育种提供理论参考。
1 材料与方法
1.1 供试材料与试验设计
参试的2个甜瓜品种为耐涝性稍强的脆禧和耐涝性较弱的世纪蜜,2个品种均由上海市农业科学院园艺研究所选育,试验在上海市农业科学院园艺研究所培养室进行。2017年2月5日将饱满、整齐一致的种子浸种催芽,出芽后播于装有西甜瓜专用育苗基质的塑料营养钵中,置于昼(28±1)℃/夜(22±1)℃培养室中进行幼苗培养,光照12 h,光照强度为400 μmol·m-2·s-1左右。
2017年2月25日,当幼苗长到三叶一心时,选取生长势基本一致的幼苗,采用“双套盆法”进行淹水处理:半淹水处理(T1)为水面淹没植株根部基质,超过基质表面0~1 cm;全淹水处理(T2)为水面刚淹过植株生长点;对照(CK)进行常规水分管理。注意每天及时补充散失的水分,确保处理所需的水面高度。分别在淹水1、3 d 及解除胁迫恢复后1、3、5 d 时取样,测定根系生长指标和根系活力,然后取幼苗生长点下第2片完全展开叶片测定呼吸酶和抗氧化酶活性指标,每处理50株,3次重复。
1.2 测定指标与方法
1.2.1 根系生长指标的测定 于淹水处理1、3 d 及解除胁迫后1、3、5 d 时,先用自来水将根部基质冲掉,再用蒸馏水冲洗3次,用干净的滤纸吸干根部表面的水分,测定根系鲜质量,之后在105 ℃的烘箱中杀青30 min,然后80 ℃条件下烘干至恒重,测定根系干质量。
1.2.2 根系活力的测定 采用李合生(2000)的TTC 法测定根系活力。
1.2.3 呼吸酶活性的测定 按照Mustroph 和Albrecht(2010)的方法提取酶液,参考Waters 等(1991)方法测定ADH 活性,依据郭欣欣等(2015)的方法测定LDH 活性、苹果酸脱氢酶(malate dehydrogenase,MDH)和琥珀酸脱氢酶(succinate dehydrogenase,SDH)活性。
1.2.4 抗氧化酶活性的测定 SOD 和CAT 活性采用Dhindsa 等(1981)的方法测定,POD 活性采用张永平等(2017)的方法测定。
1.2.5 数据分析 每个指标测定重复3次,取平均值。数据采用Excel 软件绘图,用SPSS 统计软件对平均数用Duncan’s 新复极差法进行多重比较。
2 结果与分析
2.1 淹水胁迫及胁迫解除对甜瓜幼苗根系生长的影响
从表1可以看出,甜瓜幼苗淹水1 d 时,与对照相比,耐涝性稍强的脆禧根系鲜质量无明显变化,而耐涝性较弱的世纪蜜全淹水处理(T2)比对照显著降低;当淹水3 d 时,半淹水(T1)和全淹水(T2)处理使脆禧根系鲜质量分别显著降低了7.95%和17.64%,世纪蜜分别显著下降了14.68%和26.38%。在恢复1~5 d 后,脆禧T1处理的根系鲜质量与对照之间的差异不显著,T2处理显著低于对照,世纪蜜不同处理间差异显著。
此外,随着淹水程度的增加,甜瓜幼苗根系干质量逐渐下降(表1)。其中,淹水1 d 和3 d 时,2个品种T1和T2处理的根系干质量均与对照存在显著差异(除脆禧T1处理1 d 外),T2处理下降程度明显大于T1处理,且世纪蜜下降程度大于脆禧。在胁迫恢复1 d 时,脆禧不同处理之间差异显著,胁迫解除3 d 和5 d,T1处理的根系干质量分别恢复至对照的96.06%和97.17%,与对照相比差异不显著,但T2处理仍显著小于对照;世纪蜜胁迫解除后,T1、T2处理的根系干质量显著小于对照。
2.2 淹水胁迫及胁迫解除对甜瓜幼苗根系活力的影响
由图1可见,淹水1 d 时,脆禧幼苗T1和T2处理的根系活力与对照相比差异均不显著,随着淹水胁迫时间的延长根系活力急速下降,胁迫3 d 时脆禧T1和T2处理的根系活力分别比对照显著降低了17.14%和40.64%。淹水1 d 的世纪蜜T1处理的幼苗根系活力与对照无显著差异,T2处理则比对照显著降低,胁迫3 d,T1和T2处理的根系活力分别比对照显著降低了25.90%和58.34%,耐涝性较弱的世纪蜜降低幅度大于耐涝性稍强的脆禧。胁迫恢复后,随着时间的延长,2个品种的根系活力逐渐上升,T1处理在胁迫解除5 d 时与对照接近,T2处理下脆禧和世纪蜜与对照相比分别降低13.63%和21.82%,脆禧降低幅度小于世纪蜜,说明T2处理下脆禧根系活力恢复比世纪蜜快。
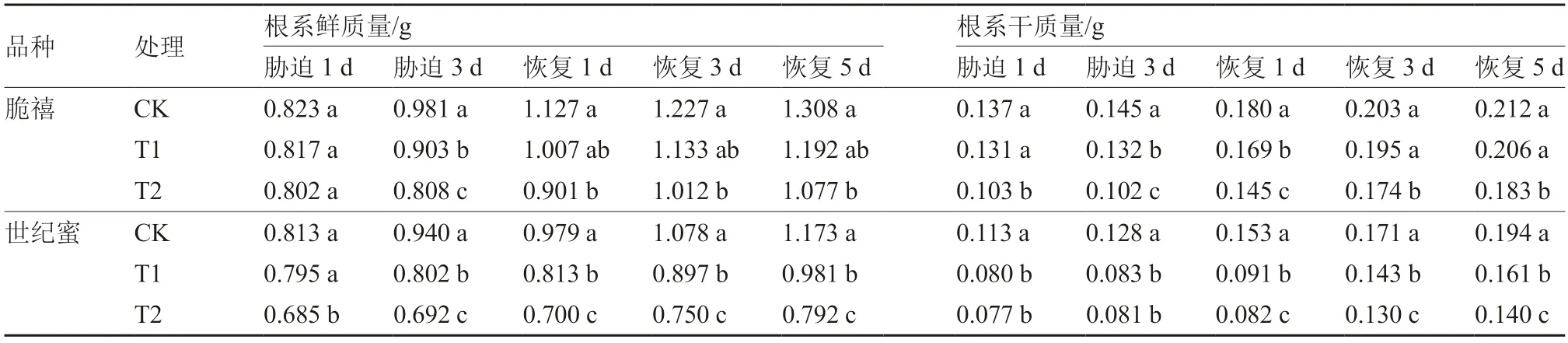
表1 淹水胁迫及胁迫解除对甜瓜幼苗根系生长的影响
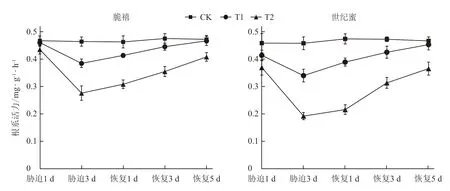
图1 淹水胁迫及胁迫解除对甜瓜幼苗根系活力的影响
2.3 淹水胁迫及胁迫解除对甜瓜幼苗根系有氧呼吸酶活性的影响
由图2-A 和2-B 可以看出,淹水处理的2个甜瓜幼苗根系的MDH 活性显著低于对照,淹水胁迫3 d 时达到最低,脆禧T1和T2处理分别比对照显著降低了27.31%和34.66%,世纪蜜分别比对照显著降低了35.88%和48.74%,耐涝性较弱的世纪蜜降低幅度大于耐涝性稍强的脆禧。胁迫解除后,随着时间的延长,2个品种的MDH 活性逐渐上升,在胁迫解除5 d 时脆禧T1处理与对照接近,但其他处理仍未恢复到对照水平。SDH 活性与MDH 活性的变化趋势基本相同(图2-C 和2-D),且当淹水胁迫3 d 时达到最低值,脆禧T1和T2处理的SDH活性分别比对照显著降低了17.10%和41.53%,世纪蜜分别比对照显著降低了23.22%和59.24%,耐涝性较弱的世纪蜜降低幅度大于耐涝性稍强的脆禧。
2.4 淹水胁迫及胁迫解除对甜瓜幼苗根系无氧呼吸酶活性的影响
在LDH 活性方面(图3-A 和3-B),在淹水1 d 后,T1和T2处理的2个甜瓜品种均显著高于对照;与对照相比,脆禧分别升高了35.39%和77.78%,世纪蜜分别升高了29.72%和80.72%;在淹水3 d 后,2个品种T1、T2处理的LDH 活性均下降。胁迫解除后,各处理的LDH 活性仍旧不断降低,随着恢复时间的延长,在胁迫解除5 d 后,各处理的LDH 活性接近对照,且差异不显著。从图3-C 和3-D 可以看出,淹水1 d 后,2个甜瓜品种的T1和T2处理的ADH 活性显著高于对照,随着胁迫时间的延长,ADH 活性逐渐增加。胁迫3 d 时,ADH 活性达到最高,与对照相比,脆禧T1、T2处理的ADH 活性分别升高了51.58%和67.37%,世纪蜜分别升高了39.76%和48.19%,脆禧ADH 活性增加的幅度大于世纪蜜,T2处理的ADH 活性增加的幅度大于T1处理;胁迫解除后,各处理的ADH 活性逐渐下降,随着恢复时间的延长,在胁迫解除5 d 时,各处理与对照相比差异均不显著。
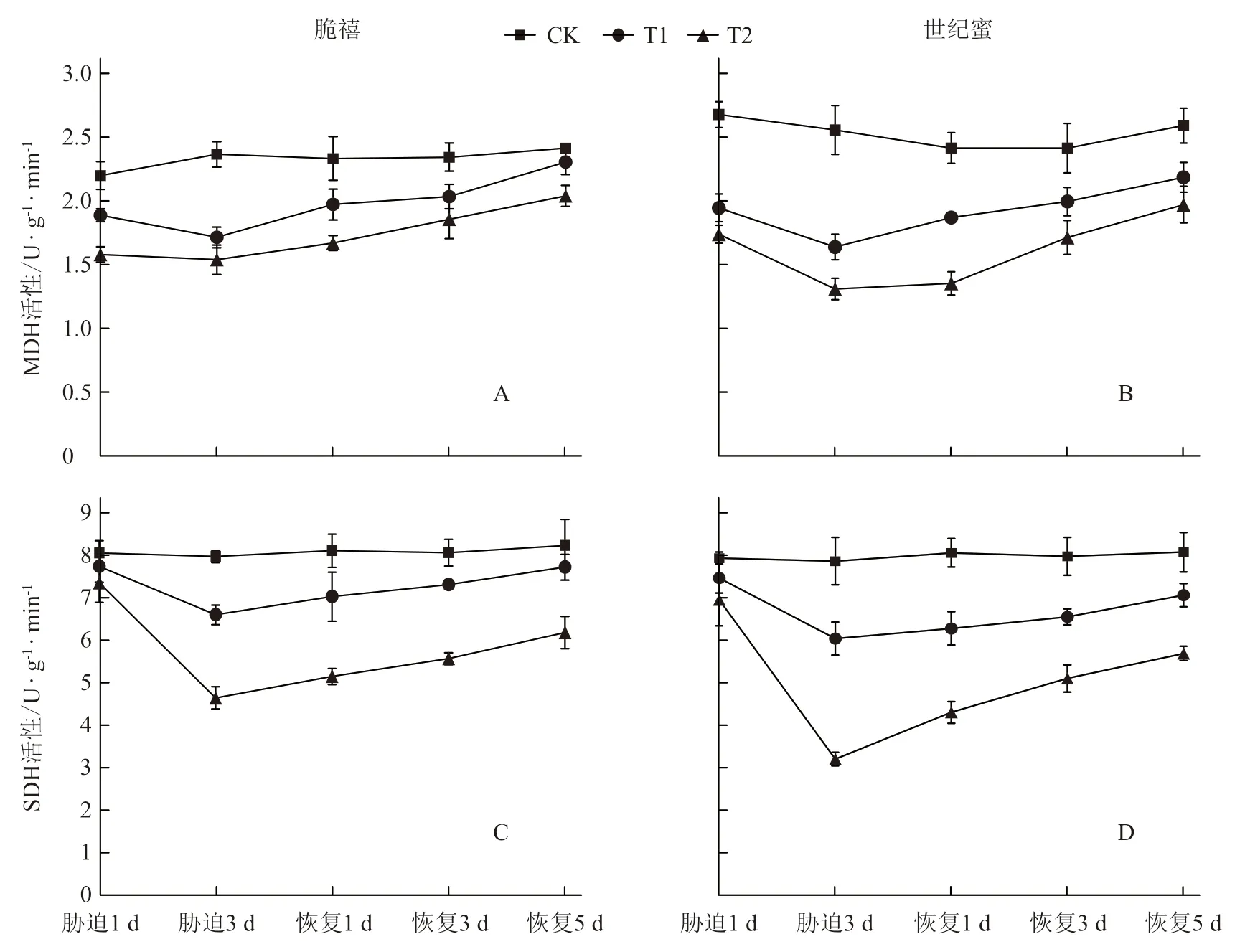
图2 淹水胁迫及胁迫解除对甜瓜幼苗根系有氧呼吸酶活性的影响
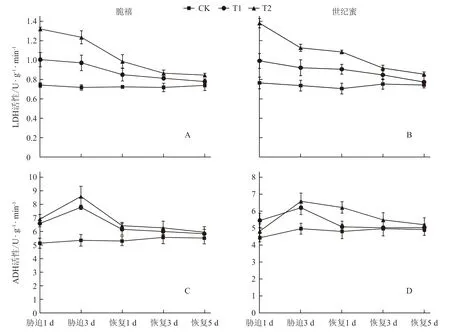
图3 淹水胁迫及胁迫解除对甜瓜幼苗根系无氧呼吸酶活性的影响
2.5 淹水胁迫及胁迫解除对甜瓜幼苗叶片抗氧化酶活性的影响
淹水胁迫下,2个甜瓜品种的SOD 活性均发生了变化(图4-A 和4-B)。淹水胁迫1 d 后脆禧T1处理的SOD 活性与对照相比有所下降,但差异不显著,T2处理则显著高于对照。胁迫3 d 时,T1和T2处理分别比对照升高了7.86%和37.36%;与对照相比,淹水处理1 d 后世纪蜜的SOD 活性增加,但只有T2处理显著高于对照,胁迫3 d 时,T1和T2处理分别比对照显著升高了15.78%和43.30%,耐涝性较弱的世纪蜜升高幅度大于耐涝性稍强的脆禧。胁迫解除后,随着时间的延长,2个品种的SOD 活性逐渐降低,T1处理在胁迫解除5 d 时与对照接近,T2处理仍未恢复到对照水平。
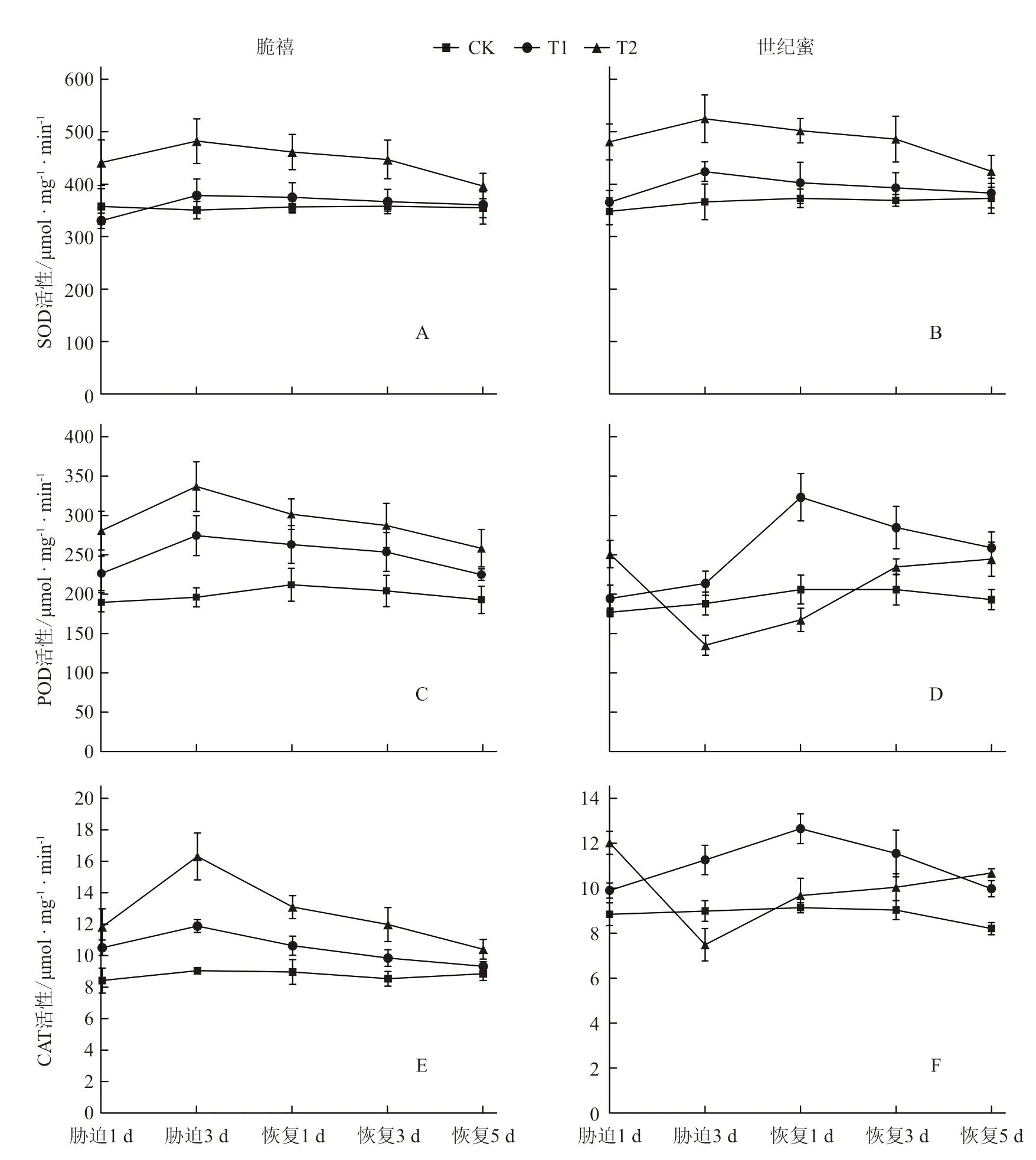
图4 淹水胁迫及胁迫解除对甜瓜幼苗叶片抗氧化酶活性的影响
在POD 活性方面(图4-C 和4-D),淹水1 d时,2个甜瓜品种T1和T2处理的POD 活性均显著高于对照(世纪蜜T1处理除外);随着胁迫时间的延长,脆禧POD 活性逐渐增加,在淹水3 d 时,T1和T2处理分别比对照显著增加了40.16%和72.13%;胁迫解除后,随着恢复时间的延长,POD活性逐渐下降,在胁迫解除5 d 时,仍未恢复到对照水平。而世纪蜜随着胁迫时间的延长,T1处理的POD 活性逐渐上升,T2处理急剧下降,胁迫3 d 时,与对照相比显著降低了28.21%;胁迫解除1 d 时,T1处理的POD 活性继续上升,随着恢复时间的延长,开始下降,T2处理则从胁迫解除后逐渐上升,在胁迫解除5 d 时,T1、T2处理的POD活性均显著高于对照。
CAT 活性变化趋势与POD 活性基本一致(图4-E 和4-F)。
3 讨论与结论
淹水胁迫抑制植物的幼苗生长,降低植物的干物质积累,造成植物个体生理差异。本试验结果表明,淹水胁迫下,2个甜瓜品种的根系生长均受到抑制,根系的鲜质量和干质量均低于对照,且耐涝性稍强的脆禧比耐涝性较弱的世纪蜜降低幅度小。在淹水胁迫解除5 d 后,T1处理的脆禧根系鲜质量和干质量与对照均没有显著差异,这与郭欣欣等(2015)在不结球白菜上的研究结果一致。
根系活力作为植物根系适应性的一个生理遗传指标,能够反映根系对营养的吸收及合成能力的强弱。研究发现,根系是吸收养分和水分的重要器官,因此许多胁迫主要是通过根系对植物造成伤害,根系受到伤害后,根系活力降低,吸收能力减弱(王俊良,2010)。周广生和朱旭彤(2002)的研究结果表明,淹水条件下小麦的初生根和次生根根系活力与不淹水的对照相比显著下降。本试验中,2个甜瓜品种根系活力随着淹水胁迫时间的延长,均有不同程度的降低,其中世纪蜜的降低幅度大于脆禧,这与刘华山等(2001)对芝麻的研究结果有一定的相似性。在淹水胁迫解除后,脆禧的根系活力恢复比世纪蜜幅度大,说明其根系的耐淹水性较强。
作为根系代谢的中心,呼吸作用对养分的吸收、根系更新和植株生长发育具有重要作用。水分胁迫会造成根系缺氧,三羧酸循环和电子传递受阻,细胞能荷水平明显下降,根系功能减弱,从而导致植物生长发育迟缓,为了减轻低氧胁迫造成的伤害,植物启动无氧呼吸代谢,将NADH 氧化成NAD+,产生ATP(Sachs et al.,1996)。本试验结果表明,淹水胁迫3 d 后,LDH 和ADH 无氧呼吸酶活性显著高于对照,MDH 和SDH 有氧呼吸酶活性显著低于对照,在胁迫解除后,T1、T2处理下的无氧呼吸和有氧呼吸酶活性与对照之间差异均明显减小,胁迫解除5 d 时,根系无氧呼吸酶活性已接近对照水平,有氧呼吸MDH 活性脆禧T1处理恢复到对照水平。LDH 是乳酸发酵途径的关键酶,而乳酸积累引起细胞质酸化是造成低氧胁迫下细胞伤害的主要原因之一(Chen &Qualls,2003)。淹水胁迫1 d 时,与对照相比,耐涝性较弱的世纪蜜T2处理的LDH 活性的增幅最大,表明其细胞质酸化较重,进而导致呼吸强度下降得更快,使其对淹水更敏感。耐涝性稍强的脆禧根系中ADH 活性增幅高于耐涝性较弱品种世纪蜜,由于乳酸发酵产生的乳酸可酸化细胞质,而乙醇发酵产生的乙醇为中性分子,能扩散到外部环境中,对植物伤害较小(李璟 等,2007)。因此,淹水胁迫下,脆禧根系中的ADH 活性升幅较高和LDH 活性升幅较小可能是其适应不利环境的能力强于世纪蜜的一个重要原因,这与康云艳等(2008)在低氧胁迫下黄瓜的研究相一致。
为了抵御淹水胁迫下活性氧的毒害作用,植物体内的抗氧化酶类(SOD、POD、CAT 等)能够清除自由基,缓解淹水胁迫对植物造成的伤害(孙小艳 等,2018)。研究表明,SOD、POD和CAT活性高,能够保护植物在长时间逆境胁迫下免受活性氧的伤害(Limón-Pacheco &Gonsebatt,2009)。作为植物细胞中最主要的抗氧化酶,SOD 显示出极强的调节作用,增强了植物的抗逆性。本试验结果表明,脆禧在淹水胁迫1 d 时,T1处理的SOD 活性低于对照,这可能是因为SOD 易受底物活性氧浓度的诱导,但淹水初期活性氧的积累不足以诱导SOD活性发生显著变化,随着淹水胁迫时间的延长,甜瓜幼苗对淹水胁迫环境的适应,SOD 活性开始逐渐上升,胁迫解除后,SOD 活性逐渐下降,这与何嵩涛等(2000)的研究结果相似。但是H2O2也会对植物产生毒害作用,而POD 可以催化H2O2分解为H2O 和O2,是细胞内H2O2的重要清除剂,所以保护酶系统的协调一致对降低活性氧对植物的伤害具有非常重要的意义。淹水胁迫初期(1 d),2个甜瓜品种POD 和CAT 活性都显著高于对照(世纪蜜T1处理除外),随着淹水胁迫时间的延长,耐涝性较弱的世纪蜜T2处理的POD 和CAT 活性逐渐下降,而脆禧活性持续增加,这种酶活性的变化可能是因为淹水时植物体内活性氧含量的增加不断激活保护酶体系的表达,以此帮助植物适应环境,POD、CAT 活性与SOD 活性变化不同步的情况与张晓平等(2006)的研究结果相似。所以在淹水胁迫下,不同植物的抗氧化酶活性的变化差异相对较大,这也反映出不同植物抗氧化防御机制存在较大差异。
综上所述,淹水胁迫使甜瓜幼苗有氧呼吸受到阻碍,同时促进了无氧呼吸代谢,根系呼吸代谢随着淹水时间加长和程度加深,受抑制的程度更加严重,最终导致根系生长受到抑制,且耐涝性较弱的世纪蜜受抑制程度大于耐涝性稍强的脆禧。
