飞蝗磷酸乙酰氨基葡萄糖变位酶基因表达及功能
2019-11-21张政张学尧张建珍刘晓健
张政,张学尧,张建珍,刘晓健*
(1.山西大学 应用生物学研究所,山西 太原 030006;2.山西大学 生命科学学院,山西 太原 030006)
0 引言
昆虫经过变态获得飞翔和生殖的能力,从而使昆虫扩大了生存的领域,也因此增强了适应能力[1]。飞蝗是一种主要分布于亚、欧、非、澳四大洲的破坏力极强的洲际性农业害虫。目前在治理蝗虫工作中以化学农药为主,但频繁使用化学农药会产生一定程度的抗药性[2-3],导致防治效果不佳,还会污染生态环境,所以研发绿色新型农药对我国防治蝗虫工作具有重要意义。
几丁质是昆虫表皮和中肠围食膜等的主要成分,高等动物没有几丁质,因此昆虫几丁质代谢途径作为重要靶标对害虫防治极具研究价值[4]。几丁质的合成中有8种酶的参与,每一种酶都起着至关重要的作用[5-6],其中研究较多的是海藻糖酶(Trehalase)和几丁质合成酶(Chitin synthase)。本课题组已有报道表明:飞蝗有4个海藻糖酶基因(LmTreS1、LmTreS2、LmTreM和LmTreM-like)[7-8]、1个葡糖胺-6-磷酸-N-乙酰转移酶基因(glucosamine-6-phosphate-N-acetyltransferase,LmGNA)[9]、2个UDP-N-乙酰葡糖胺焦磷酸化酶基因(LmUAP1和LmUAP2)[10]和2个几丁质合成酶基因(LmCHS1和LmCHS2)[11-13],其中LmUAP1和LmCHS1基因负责体壁几丁质的合成,而LmCHS2基因负责中肠围食膜几丁质的合成,RNAi分别沉默上述基因的表达后,均导致飞蝗高死亡率,而其他基因不是飞蝗生长发育必需的。磷酸乙酰氨基葡萄糖变位酶(Phosphoacetylglucosamine mutase,PAGM)作为几丁质合成中的关键酶之一,仅在埃及伊蚊(Aedesaegypti)中有所报道,作者克隆获得一条AePAGM基因,并且发现该基因在整个龄期均有表达[14],尚未在其他昆虫中开展相关研究。
本文以飞蝗为研究对象,通过搜索飞蝗转录组数据库及PCR技术,得到PAGM基因全长cDNA序列,将其命名为LmPAGM;通过Real-time PCR技术分析了LmPAGM基因在5龄期的时空表达模式;利用RNAi技术研究LmPAGM的生物学功能,为基于RNAi的飞蝗防治奠定理论依据。
1 材料与方法
1.1 供试昆虫
飞蝗虫卵购自河北沧州蝗虫养殖公司,在光周期为14 L:10 D,湿度为60%,温度为30±2℃的条件下,在人工气候箱进行孵化。待卵孵化后,将飞蝗若虫转移至干净的纱笼中,以新鲜小麦幼苗和麦麸进行饲喂。
1.2 供试药品

1.3 实验方法
1.3.1 cDNA全长序列及基因组序列获得
通过搜索飞蝗转录组数据库,得到PAGM基因全长cDNA序列,将其命名为LmPAGM。为了进一步验证该序列,利用Primer 5.0设计特异性全长引物,由南京金斯瑞生物科技有限公司合成引物(表1)。利用五龄飞蝗整虫cDNA为模板进行PCR扩增,扩增产物纯化后连接至T载体,细菌培养、质粒DNA的提取和PCR鉴定后,将产物送至南京金斯瑞生物科技有限公司进行测序,测序正确的LmPAGM使用ExPaSy 网站上的相关软件计算分子量及等电点。在飞蝗基因组数据库[15]中输入LmPAGM基因的cDNA序列进行Blast比对,分析该基因的内含子与外显子序列。
1.3.2 不同组织部位及发育日龄飞蝗磷酸乙酰氨基葡萄糖变位酶基因的表达
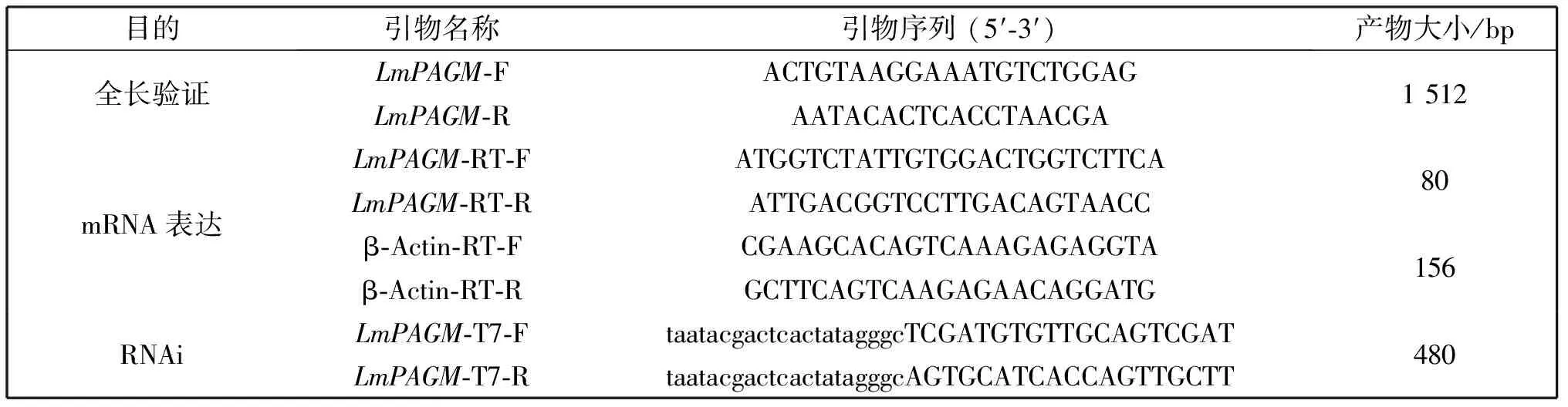
表1 LmPAGM研究中使用的引物
1.3.3 RNAi分析飞蝗磷酸乙酰氨基葡萄糖变位酶基因的功能
通过E-RNAi 网站设计含有T7序列(taatacgactcactataggg)的LmPAGMRNAi引物,利用五龄飞蝗整虫cDNA为模板,2×Taq PCR MasterMix进行PCR扩增,扩增产物进行凝胶电泳确定其为单一目的条带,之后通过胶回收试剂盒FastPure Gel DNA Extraction Mini Kit进行目的条带回收,回收产物作为合成dsRNA的模板。运用试剂盒 HiScribeTMT7 Quick进行dsRNA的合成。合成后的dsRNA浓度统一稀释为2 μg/μL。选取N5D2飞蝗若虫进行dsRNA的注射,用微量注射器将LmPAGMdsRNA从飞蝗第2 和3 腹节连接处注入体腔,注射剂量为10 μg/头,注射相同体积水的N5D2飞蝗若虫作为对照,15头N5D2飞蝗若虫组成1个生物学重复,共计3个生物学重复45头,注射完毕后,将所有试虫放置于人工气候箱中饲养。CK与处理组各30头,在N5D4取CK与处理组各9头生物学重复进行体壁RNA的提取。反转录为cDNA后采用Real-time PCR方法对该基因的沉默效果进行检测,操作同上。其余试虫用来观察表型。
2 结果与分析
2.1 飞蝗PAGM基因cDNA的序列分析
对飞蝗转录组数据库进行了全面搜索,得到1条PAGM基因cDNA序列。利用NCBI的BLASTX分析,结果表明这段序列是PAGM的全长序列,命名为LmPAGM。并利用PCR对其进行了全长验证,获得的LmPAGM全长1 512 bp,其中开放阅读框1 491 bp,编码497个氨基酸,5′非翻译区和3′-非翻译区的长度分别为99 bp和258 bp,GenBank号:MK568624利用ExPASy Proteomics website,预测LmPAGM蛋白的分子量为54.2 kDa,等电点为5.69。LmPAGM的基因组全长超过60 kb,含有17个外显子,最小的外显子大小为308 bp,最大的外显子大小为71 bp,如图1A。BLAST分析表明,LmPAGM的氨基酸序列与丽蝇蛹集金小蜂(Nasoniavitripennis)的氨基酸一致度最高为56%,如图1B。

Fig.1 Genomic structure of LmPAGM and alignment with other PAGM of insects图1 飞蝗LmPAGM基因结构图及其他昆虫PAGM氨基酸比对Lm (MK568624): Locusta migratoria 飞蝗; Aa (XP-001661733.1): Aedes aegypti 埃及伊蚊;Ag (EAA04228.3): Anopheles gambiae 冈比亚按蚊; Tc (XP-973346.2): Tribolium castaneum 赤拟谷盗;Nv (XP-016837506.1): Nasonia vitripennis 丽蝇蛹集金小蜂;Dm (NP-648588.1): Drosophila melanogaster黑腹果蝇
2.2 飞蝗PAGM基因在不同组织部位的表达
根据已经得到的飞蝗PAGM基因核苷酸序列,设计特异性的表达引物,利用 Real-time PCR对该基因在不同组织部位的表达进行了分析。结果显示:LmPAGM在所有组织中(体壁、前肠、中肠、胃盲囊、后肠、脂肪体、马氏管和翅)都有表达,各组织之间的表达量没有显著差异,见图2。
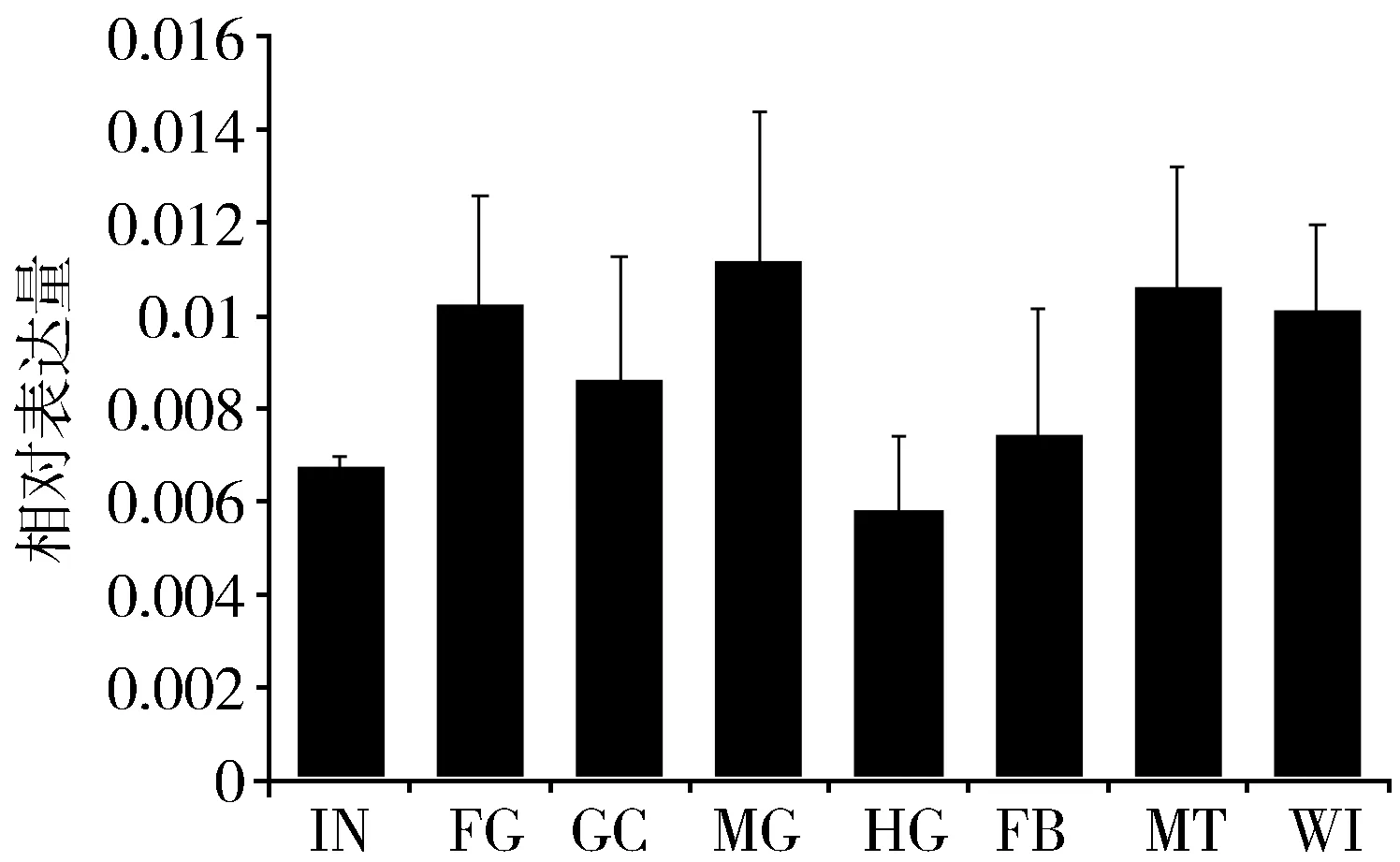
Fig.2 Expression of LmPAGM in differenttissues of the 5th instar nymphs of L. migratoria图2 五龄飞蝗若虫不同组织部位LmPAGM基因的表达注:IN (Integument):体壁;FG (Foregut):前肠;GC (Gastric caeca):胃盲囊; MG (Midgut):中肠;HG (Hindgut):后肠;FB (Fat body):脂肪体;MT (Malpighian tubules):马氏管; WI (Wing):翅。
2.3 五龄期不同天数飞蝗PAGM基因的表达分析
运用Real-time PCR对PAGM基因5龄期不同天数的表达进行了分析。实验结果显示:LmPAGM在每1天都有表达,其中第8天的表达量最高(图3)。
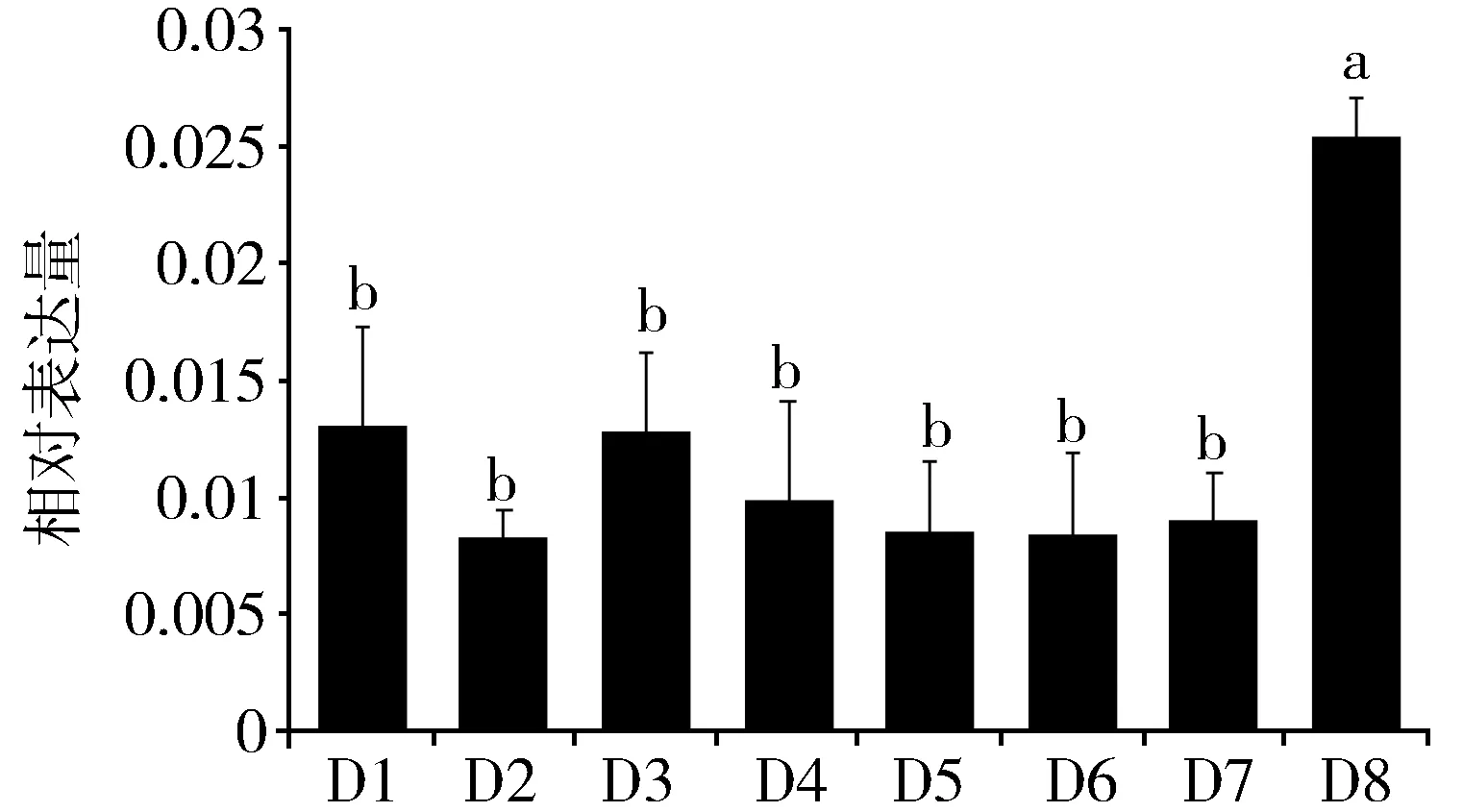
Fig.3 Expression of LmPAGM in different daysof the 5th instar nymphs of L. migratoria图3 飞蝗五龄期不同天数LmPAGM基因的表达注:柱形图上不同的字母代表基因表达差异显著(P<0.05,Tukey’s HSD;n=3.)
2.4 飞蝗PAGM基因的生物学功能分析
将dsLmPAGM注射至N5D2飞蝗若虫,48 h后收集整虫样品提取RNA,利用Real-time PCR技术检测LmPAGM基因的沉默效率,结果表明与对照组相比,注射dsLmPAGM组中LmPAGM的表达被显著沉默,沉默效率达70.3%(图4A)。30头对照在第8天成功羽化为成虫,注射dsLmPAGM的实验组在第8天有25头成功羽化为成虫,11头试虫出现死亡,注射dsLmPAGM组飞蝗致死率约为30%(图4B)。
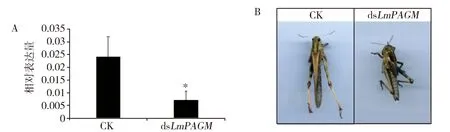
A:LmPAGM沉默效率检测;B:LmPAGM沉默后的表型图Fig.4 Effects of dsLmPAGM injected into the 5th instar nymphs on theexpression of LmPAGM and the development of L. migratoria图4 注射LmPAGM基因的dsRNA至5龄期若虫后对LmPAGM基因表达量及发育的影响
3 讨论
昆虫几丁质合成通路从血淋巴中的海藻糖被转运进入细胞质开始,由8种酶依次催化生成葡萄糖、葡萄糖-6-磷酸、果糖-6-磷酸、D-葡萄糖胺-6-磷酸、N-乙酰葡萄糖胺-6-磷酸、N-乙酰葡萄糖胺-1-磷酸、UDP-N-乙酰葡萄糖胺(UDP-GlcNAc),最后UDP-GlcNAc在几丁质合成酶的作用下合成几丁质[16]。其中,PAGM催化N-乙酰葡萄糖胺-6-磷酸生成N-乙酰葡萄糖胺-1-磷酸。我们从不完全变态昆虫飞蝗中首次获得了磷酸乙酰氨基葡萄糖变位酶基因的cDNA序列,命名为LmPAGM,该基因编码497个氨基酸,和埃及伊蚊,冈比亚按蚊,赤拟谷盗,丽蝇蛹集金小蜂,黑腹果蝇PAGM氨基酸一致度为52%~56%。
本研究采用Real-time PCR技术对LmPAGM基因在五龄若虫期的时空表达进行了分析,发现该基因除了在体壁、前肠、中肠、后肠和翅这些含有几丁质的部位表达外,在所检测的其他组织中(胃盲囊、脂肪体和马氏管)均有表达,且没有显著差异。从果糖-6-磷酸到UDP-GlcNAc也被称氨基己糖途径,已有研究报道UDP-GlcNAc是包括几丁质在内的许多重要物质合成的前体,例如细菌合成肽聚糖、革兰氏阴性菌合成脂多糖,真菌合成细胞壁的甘露糖蛋白等,以及真核生物蛋白质及脂质的N-连接的糖基化过程也与UDP-GlcNAc有关[17]。PAGM基因除存在于昆虫外,还存在于原核生物和包括人类[18]等哺乳动物在内的许多真核生物中,因此推测飞蝗LmPAGM基因在没有几丁质的胃盲囊、脂肪体和马氏管中的表达,可能是由于其参与了蛋白质及脂质的N-连接的糖基化过程。LmPAGM在第8天的体壁表达量最高,这与飞蝗几丁质合成关键基因LmUAP1[19]和LmCHS1[19]的变化规律相似,即蜕皮前表达较蜕皮中的表达显著上升。对五龄期飞蝗若虫的体壁进行切片和H&E染色,表明蜕皮前是新表皮开始形成时期,在这段发育期旧的内表皮逐渐被消化吸收,新的并没有硬化的表皮大量形成并剧烈褶皱,因此与几丁质合成相关基因的高表达与新表皮形成需要大量几丁质合成有关[19]。通过RNAi实验发现,与对照组相比,注射dsLmPAGM组LmPAGM的表达可以显著沉默,但是仅有约30%的飞蝗会出现死亡表型,本课题组已有研究表明几丁质合成关键基因LmUAP1[19]和LmCHS1[19]负责新表皮几丁质的形成,沉默上述基因的表达后新表皮几丁质含量较对照组显著下降,因此推测LmPAGM基因可能在几丁质合成过程中具有类似的功能。RNAi具有特异性及高效性等特点,在害虫的防治中具有潜在的应用价值。筛选和鉴定对害虫生长发育至关重要的基因是应用RNAi技术开发新型杀虫剂的关键环节[20],本文的实验结果为进一步探讨PAGM基因在几丁质合成途径中功能提供基础数据,并为基于RNAi的飞蝗防治提供了理论依据。
