糖原合成激酶3β抑制剂通过p38负调节感染性炎性反应的研究*
2019-11-14卢建强梁培松王伟佳
陈 康,傅 强,卢建强,梁培松,韩 慧,董 谦,王伟佳
(广东省中山市人民医院检验医学中心,广东中山 528403)
铜绿假单胞菌是一种条件致病菌,可引起医院获得性肺炎、角膜炎等多种机会感染[1-2]。尽管抗菌药物可以杀伤细菌,却不能抑制因感染而激发的过度免疫应答。因此,如何有效抑制过度免疫应答是预防免疫病理损伤和治疗疾病的关键。糖原合成激酶3β(GSK3β)是组织器官中广泛表达的丝氨酸/苏氨酸蛋白激酶,可参与糖原合成,多种病理生理过程,如高血压、糖尿病、炎性肠病等[3-5]。此外,还可参与多种病原体感染引起的免疫应答,如病毒、真菌等[6-7]。磷酸化是调节GSK3β活性最常见的方式,其中以失活型9位丝氨酸(Ser9)的磷酸化最为常见,当Ser9的磷酸化水平降低时,提示在该疾病中GSK3β活性增高,破坏细胞的稳态,导致细胞发生氧化应激和炎性反应。丝裂原活性蛋白激酶(MAPK)是调节炎性反应通路中的重要分子,GSK3β可与MAPK家族中成员包括p38、c-Jun氨基末端激酶(JNK)、细胞外调节蛋白激酶(ERK)协同调控细胞的炎性反应。GSK3β还可参与多种疾病的调控,被用作疾病治疗的靶点。GSK3β的小分子抑制剂SB216763可抑制其活性,调节免疫应答[8],因此本研究旨在探讨感染铜绿假单胞菌时,GSK3β抑制剂调节细胞因子分泌及免疫功能的作用及机制。
1 材料与方法
1.1试剂 GSK3β抑制剂SB216763、p38抑制剂SB203580、JNK抑制剂SP600125均购自美国MCE公司;p38、磷酸化p38(P-p38)、JNK、磷酸化JNK(P-JNK)、ERK、磷酸化ERK(P-ERK)、磷酸化GSK3β失活型9位丝氨酸[P-GSK3β(Ser9)]和β-actin的抗体均购自美国CST公司;Trizol、反转录试剂盒购自美国Invitrogen公司;异丙醇和无水乙醇购自中国广州鼎国生物技术有限公司;实时荧光定量PCR(qPCR)检测用SYBR Green试剂盒购自美国Bio-Rad公司。
1.2细胞培养 小鼠巨噬细胞购自美国菌种保藏中心,细胞培养于DMEM培养基中。将4×105个/mL 小鼠巨噬细胞铺入12孔细胞培养板中,采用GSK3β抑制剂(10 μmol/L)、p38抑制剂(1 μmol/L)或JNK抑制剂(1 μmol/L)预处理细胞1 h,加入铜绿假单胞菌(MOI=1)刺激后,检测蛋白或mRNA水平的表达。
1.3免疫印迹试验(Western blot) 用冷的磷酸盐缓冲液冲洗细胞培养板中的细胞2次,加入蛋白裂解液,收集细胞裂解碎片,冰上裂解20 min。12 000 r/min离心10 min,收集蛋白裂解上清,考马斯亮蓝法进行蛋白定量。接着进行聚丙烯酰胺凝胶电泳,并电转至硝酸纤维素膜上。将膜用5%脱脂奶粉室温孵育2 h,4 ℃过夜分别孵育一抗p38(1∶1 000)、P-p38(1∶1 000)、JNK(1∶1 000)、P-JNK(1∶1 000)、ERK(1∶1 000)、P-ERK(1∶1 000)、P-GSK3β(Ser 9)(1∶1 000)、β-actin(1∶5 000),室温下孵育二抗1 h,采用电化学发光方法检测蛋白水平。
1.4qPCR RNA抽提试剂法进行RNA提取,分光光度计测RNA浓度和纯度,用于下一步实验。将提取后的RNA(1 μg)转录为cDNA,详细步骤如下:RNA 1 μg,oligo(dT)1 μL,补DEPC水至体积12 μL,混匀;65 ℃,5 min;每孔加入5×反应液4 μL,20 U/μL RNA酶抑制剂1 μL,10 mmol/L dNTP 2 μL,200 U/μL反转录酶1 μL,至总体积20 μL,混匀;42 ℃,60 min,70 ℃,5 min终止反应。反转录的cDNA保存备用。应用SYBR Green试剂盒进行扩增,详述如下:SYBR Green(2×) 10 μL,引物上下游混合液(5 μmol/L) 1.6 μL,cDNA模板2 μL,灭菌H2O补齐总体积20 μL充分混匀;按照两步法PCR扩增程序,第1步,95 ℃ 30 s,1个循环;第2步,95 ℃ 5 s,60 ℃ 30 s,40个循环PCR反应。反应结束后确认qPCR的扩增曲线和溶解曲线,得到循环阈值(Ct)值,采用2-ΔΔCT法进行相对定量。IL-10、IL-6和β-actin的引物序列见表1。

表1 引物序列

2 结 果
2.1GSK3β参与到铜绿假单胞菌感染中 实验结果显示,在铜绿假单胞菌感染小鼠巨噬细胞6 h后,P-GSK3β(Ser9)水平增高,然后随感染时间的增加,水平下调。这些结果提示,GSK3β参与到铜绿假单胞菌感染中。见图1。
2.2GSK3β抑制剂调节细胞因子的表达 实验结果显示,在铜绿假单胞菌感染巨噬细胞后,GSK3β抑制剂SB216763可促进细胞因子IL-10水平的表达,同时抑制细胞因子IL-6水平的表达。结果提示抑制GSK3β可在铜绿假单胞菌感染中发挥免疫负调的作用。见图2。
2.3GSK3β抑制剂调节MAPK家族成员的磷酸化水平 实验结果显示,在铜绿假单胞菌感染后15 min、30 min和2 h,GSK3β抑制剂可上调p38和JNK的磷酸化水平,而对ERK的磷酸化水平无调节作用。实验结果同时发现,细菌感染前后,GSK3β抑制剂对p38、JNK和ERK的蛋白水平无调节作用。见图3。

注:A表示P-GSK3β(Ser9)蛋白表达水平随感染时间的变化情况;B表示P-GSK3β(Ser9)蛋白表达定量随感染时间的变化情况;***P<0.001
图1 GSK3β参与到铜绿假单胞菌感染中

注:A表示IL-10基因水平表达情况;B表示IL-6基因水平表达情况;*P<0.05;***P<0.001
图2 GSK3β抑制剂通过p38调节细胞因子的表达
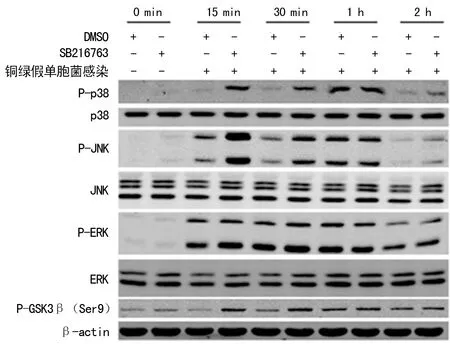
图3 GSK3β抑制剂调节MAPK的磷酸化水平
2.4GSK3β抑制剂通过p38调节细胞因子的表达 实验结果显示,GSK3β抑制剂可上调细胞因子IL-10的表达,在应用GSK3β抑制剂的同时,应用p38抑制剂可抑制细胞因子IL-10的表达,应用JNK的抑制剂不可抑制IL-10的表达;GSK3β抑制剂可下调细胞因子IL-6的表达,在应用GSK3β抑制剂的同时,应用p38抑制剂可上调细胞因子IL-10的表达,而同时应用JNK的抑制剂不可上调IL-10的表达。这些结果提示,GSK3β抑制剂通过p38 MAPK信号通路调节细胞因子的表达。见图2。
3 讨 论
本研究发现,在感染铜绿假单胞菌时,GSK3β活性增高;GSK3β抑制剂可通过p38调节细胞因子的分泌,负调节免疫反应。
GSK3β参与到多种疾病的致病过程和病原体感染中[3-7],因此被用作疾病治疗的靶点。研究报道,GSK3β的抑制剂可发挥免疫调控的作用[8]。MARTIN等[9]报道,抑制GSK3β可抑制多种配体激活的炎性反应,抑制促炎性细胞因子的表达,同时促进抑炎性细胞因子IL-10的表达。在败血症休克中,抑制GSK3β可上调抑炎性因子IL-10的表达[10]。然而,SHEN等[11]报道,GSK3β抑制剂SB216763可在心肌细胞中促进脂多糖诱导肿瘤坏死因子的表达。本课题组研究发现,GSK3β抑制剂SB216763可促进抑炎性细胞因子IL-10的表达,同时抑制促炎性细胞因子IL-6的表达,在铜绿假单胞菌感染中负调节免疫应答。
GSK3β可参与多个信号通路的调控,可通过抑制NF-κB信号通路或上调环磷腺苷效应元件结合蛋白抑制免疫应答[12]。NOH等[10]报道,GSK3β抑制剂可通过抑制ERK的活性上调抑炎性因子IL-10的表达。本课题组研究发现,GSK3β抑制剂可上调p38和JNK的磷酸化水平,并通过p38 MAPK信号通路调节细胞因子IL-10和IL-6的表达,发挥免疫负调节作用。
4 结 论
在感染铜绿假单胞菌后,GSK3β的活性增高;应用GSK3β的抑制剂SB216763可调节细胞因子的表达,负调节免疫应答;进一步实验发现,GSK3β抑制剂通过p38 MAPK信号通路调节细胞因子的表达。这些研究提示在铜绿假单胞菌感染中GSK3β可作为免疫调控的靶点。
