不同灌溉条件下冠菌素对大豆光合特性与产量的调控效应
2019-11-13鱼海跃张钰石张明才李召虎
鱼海跃 闫 岩 张钰石 张明才 李召虎
不同灌溉条件下冠菌素对大豆光合特性与产量的调控效应
鱼海跃 闫 岩 张钰石 张明才*李召虎
植物生长调节剂教育部工程研究中心 / 中国农业大学农学院, 北京 100193
干旱是限制大豆丰产稳产的重要因素之一, 利用生物调节剂提高大豆耐旱性是生产中一种新型的生物节水管理模式。本研究在常规灌溉与无灌溉条件下, 采用生物调节剂冠菌素(COR)于大豆初花期进行叶面喷施处理, 研究COR对植株农艺性状、叶片水势和光合特征、产量及其构成因素的调控效应。试验结果表明: 在正常灌溉条件下, COR处理对大豆叶片水势、叶绿素含量、光合速率、叶绿素荧光参数、RuBP羧化酶和SPS活性等影响较小, 与对照相比其产量和生物量差异不显著。但在生长季无灌溉的雨养条件下, COR处理会显著提高大豆开花后叶片水势、叶绿素含量、光合速率和叶绿素荧光参数, 增加叶片RuBP羧化酶和SPS活性, 改善大豆产量构成因素, 最终导致籽粒产量增加。总之, 在雨养条件下, COR对大豆光合特征和产量形成具有积极的调控效应。
冠菌素; 灌溉; 大豆; 光合特性; 产量
大豆是我国主要粮食作物之一, 也是人们生活中优质蛋白和食用油脂的重要来源, 但大豆生长发育过程中需水量大, 且对干旱较为敏感[1]。我国大豆种植区域广泛, 且存在水资源不足或降雨量季节间分布不均, 导致季节性干旱和间歇式干旱时常发生, 严重制约大豆产量提高, 加剧了我国大豆对外依存度。因此, 提高大豆抗旱性和水资源利用效率, 是我国大豆可持续生产需解决的重要问题, 也是国内外农学家关注的热点问题。当前, 利用抗旱品种和生物节水充分挖掘大豆本身的抗旱潜力, 是缓解我国大豆生产中干旱和水资源不足的一个切实可行途径。利用植物生长调节物质调节和控制大豆生长发育和生理生化过程, 提高植株耐旱性, 是一种新兴的作物生物节水模式。有研究表明, 植物生长调节剂如油菜毒内酯、烯效唑、MeJA等能够显著增强大豆抗旱能力, 提高大豆单株生产力[2-4]。
冠菌素(Coronatine, COR)是丁香假单胞菌分泌的一种代谢产物, 具有与茉莉酸类化合物类似的结构和功能, 是一种低分子量高活性的生长调节物质。近年研究表明, 应用COR提高了植物体内抗氧化酶活性和ROS清除能力, 增强植物对非生物逆境如干旱、盐害、低温等的耐受性或抗性[5-9]。我们前期研究表明, 在温室盆栽干旱胁迫下, COR处理显著提高了大豆叶片水势和相对含水量, 维持膜稳定性和较高的光合性能, 增强了干旱胁迫下14C转运, 促进了干物质积累, 提高了生物量[10]。但在大田开放条件下, 未见有关COR在大豆上应用效应及其抗性调控相关研究报道, 从而限制了COR在大豆抗旱栽培中的应用。本研究将探索大田水分胁迫条件下COR对大豆产量形成的调控效应, 从光合特征、水分状况角度解析其提高大豆抗旱的生理生化机制, 为建立大豆高效生物节水栽培技术提供理论依据。
1 材料与方法
1.1 试验设计
试验于2015—2016年在河北省沧州市中国农业大学吴桥试验站(37°41′N, 116°37′E)进行。试验点属于温带季风气候, 常年降雨量562 mm, 主要分布夏季6月至8月份, 全年光照2725 h, 年平均气温12.9℃,无霜期201 d。夏大豆2个生长季天气条件如图1, 2016年各月份平均气温高于近25年平均气温和2015年, 而2015年各月份平均气温与近25年差异不大; 近25年平均降雨量为404 mm, 2015年降雨量为351 mm, 而2016年降雨量为499 mm。试验地土质为沙壤土, 含全氮1.21 g kg–1, 有效钾124.5 mg kg–1, 有效磷51.1 mg kg–1, 有机质16.2 g kg–1。
供试品种为中黄13, 种植密度为22.5万株 hm–2。按裂区试验设计, 主区为水分处理, 设常规灌溉(分枝期、花荚期和鼓粒期浇3次水, 每次灌溉750 m3hm–2)和全生育期无灌溉(雨养条件)两处理, 各处理均足墒播种。裂区设调节剂COR和水分处理2个因素, 其中COR浓度为0.1 μmol L–1(依据我们前期田间试验筛选出的适宜浓度[11]), 在初花期进行叶面喷施, 以清水处理为对照; 小区面积4 m × 5 m, 裂区按随机排列, 4次重复。行距为30 cm, 株距为10 cm。播种期分别为2015年6月15日和2016年6月18日, 收获期为2015年10月6日和2016年10月8日。整个生育期, 供给一次性基施肥料, 复合肥(N∶P2O5∶K2O为15∶15∶15)为300 kg hm–2。其他农艺措施参照当地作法。
1.2 取样及测定方法
大豆初花期处理时标记同天开花且长势一致的植株, 在花后5 d、15 d和25 d选取植株10株, 进行光合速率、荧光参数和叶片水势的测定, 另一部分以液氮速冻, −20℃保存, 用于酶活性测定。取样与测定的叶片为植株倒三叶[12]。
1.2.1 叶绿素含量与光合速率测定 采用SPAD-502叶绿素测定仪测定叶片相对叶绿素含量, 测定时避开叶脉位置; 采用便携式光合系统测定仪LI-6400 (LI-COR Lincoln, USA)测定大豆标记叶片的净光合速率(n), 每个处理测定时重复10次。
1.2.2 叶绿素荧光参数测定 利用PAM-2100便携式叶绿素荧光仪测定叶绿素荧光参数。将叶片暗适应20~30 min, 先照射弱红光(强度为0.06 μmol m–2s–1), 测初始荧光o。而后再饱和脉冲光(强度大于6000 μmol m–2s–1), 测最大荧光m。当光合作用为稳态后, 获得荧光参数s。随后照饱和脉冲光, 测定最大荧光(m¢)。记录仪器输出PSII最大光化学量子产量(v/m)等参数值。
1.2.3 叶片水势测定 利用3115型压力势水势仪测定。选取叶片, 从叶柄处切断, 将叶片迅速装入夹样器的中央孔中, 切口露出垫圈3~5 mm, 旋紧螺旋环套。以每秒30~50 kPa的速度加压。左手持放大镜从侧面仔细观察样品切口的变化, 当切口出现水膜时, 迅速关闭调压阀, 记录压力表读数, 并计算叶片水势。
1.2.4 叶片1,5-二磷酸核酮糖羧化酶(RuBPcase)和蔗糖磷酸合成酶(SPS)活性测定 参照顾万荣等[13]研究方法测定RuBPcase活性; 参照Huber[14]研究方法测定SPS活性。
1.2.5 植株性状、产量及其构成因素测定 成熟期小区实收测产, 3个重复。测产前, 每个小区选取2行, 随机挑选20株调查株高、单株分枝数、单株节数、单株生物量及产量构成因素单株荚数、单株粒数和百粒重。
1.3 数据分析
用SPSS 17.0 进行统计分析。
2 结果与分析
2.1 不同田间灌溉条件下COR对大豆产量与植株农艺性状的调控
在正常田间灌溉条件下, COR对大豆单株结荚数、单株粒数和百粒重影响较小, 产量在处理与对照间差异不显著(表1), 且在2个生长季内COR处理表现出类似的效应。但2016年生长季产量构成因素如单株荚数、粒数和粒重以及产量显著高于2015年度, 这主要是2015年度夏大豆生长季降雨量显著低于2016年度造成的(图1)。在无灌溉条件下, 2个生长季COR处理均显著提高了单株荚数、粒数和百粒重, 增加了产量。

表1 不同田间灌溉条件下COR对大豆产量及其构成因素的影响
同一列中同一年份间不同小写字母表示在0.05水平差异显著。
Means within a column and same year followed by different letters are significantly different at< 0.05.
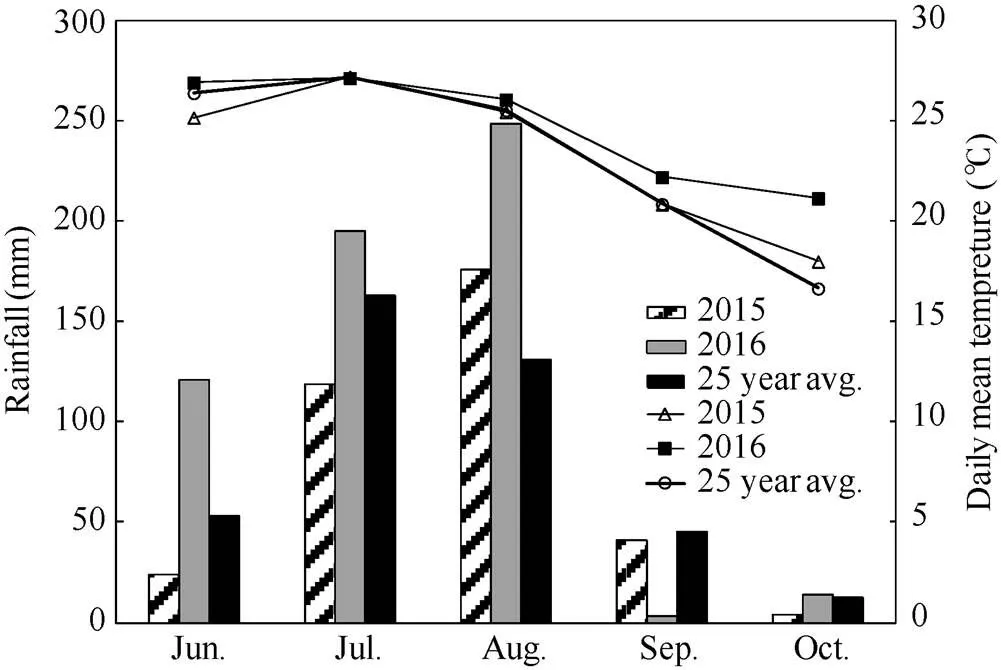
图1 2015年、2016年和25年夏大豆生长季吴桥月平均降雨量(柱)和平均气温(线)
在正常田间灌溉条件下, COR对大豆单株分枝数、株高和节数影响较小, 单株生物量处理间差异也不显著(表2)。但2015年生长季单株分枝数与节数、株高和生物量均高于2016年生长季, 这主要由年度间降雨量差异大造成。在生长季田间无灌溉条件下, COR处理对单株分枝数与节数影响较小, 但提高了株高和单株生物量, 且2个生长季均表现出相同调控趋势。
2.2 不同田间灌溉条件下COR对大豆叶片光合和叶绿素荧光特性的调控
在正常田间灌溉条件下, COR处理对叶片叶素含量和光合速率影响较小, 与对照比较差异不显著, 且年际间均表现出类似的COR调控效应(图2)。与之相比, 生长季田间无灌溉显著降低了叶片叶绿素含量和光合速率, 且2个生长季均表现出相同变化趋势。而COR处理可以缓解田间无灌溉对植株生长的抑制效应, 显著提高田间无灌溉条件下叶片叶绿素含量和光合速率。
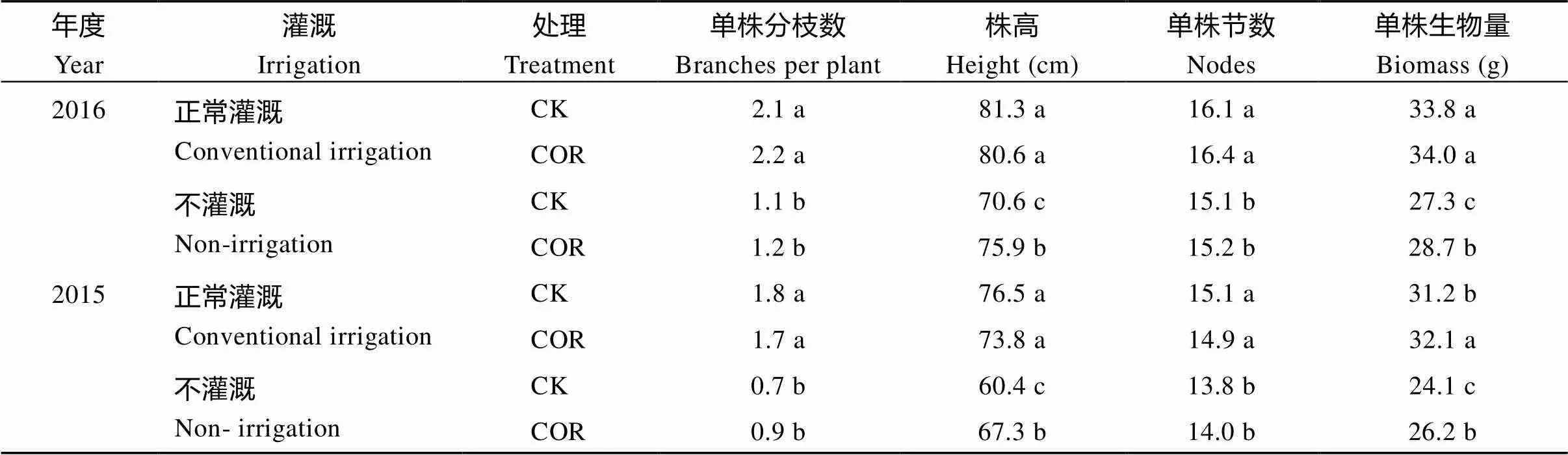
表2 不同田间灌溉条件下COR对大豆植株农艺性状与生物量的影响
同一列中同一年份间不同小写字母表示在0.05水平差异显著。
Means within a column and same year followed by different letters are significantly different at< 0.05.

图2 不同田间灌溉条件下COR对大豆叶片叶绿素含量和光合速率的影响
A和B分别表示2015年和2016年生长季大豆叶片叶绿素含量; C和D分别表示2015年和2016年生长季大豆叶片光合速率。
A and B indicate the chlorophyll content of soybean leaves during 2015 and 2016 growing seasons, respectively. C and D indicate the photosynthetic rate in soybean leaves during 2015 and 2016 growing seasons, respectively.
在正常田间灌溉条件下, COR处理对叶片叶绿素荧光参数PSⅡ最大量子产量(v/m)和PSII实际量子产量(Yield)影响较小, 且处理与对照间无显著差异, 2个生长季均表现出类似的调控效应(图3)。与之相比, 生长季田间无灌溉显著降低了叶片v/m和Yield值, 且2个生长季均表现出相同变化趋势。但是, 在生长季田间无灌溉条件下, COR处理显著提高了叶片v/m和Yield值, 且2个生长季均表现出相同的COR调控效应。
2.3 不同田间灌溉条件下COR对叶片RuBP羧化酶和蔗糖磷酸化合成酶活性的调控
在正常田间灌溉条件下, COR处理对叶片光合关键酶RuBP羧化酶活性的影响与对照没有显著差异, 且2个生长季均表现相同的变化趋势(图4)。与之相比, 生长季田间无灌溉显著降低了RuBP羧化酶活性, 且2个生长季均表现出相同变化趋势。但COR处理显著提高了生长季田间无灌溉条件下RuBP羧化酶活性, 且2个生长季均表现出相同的COR调控效应。
在正常田间灌溉条件下, COR处理对叶片蔗糖磷酸合成酶(SPS)活性的影响与对照没有显著差异, 且2个生长季均表现相同的调控趋势(图5)。与之相比, 生长季田间无灌溉显著降低了叶片SPS酶活性, 但COR处理显著提高了该条件下SPS酶活性, 且2个生长季均表现出相同的COR调控效应。
2.4 不同田间灌溉条件下COR对叶片水势的调控
在正常田间灌溉条件下, COR处理对叶片水势的影响与对照间没有显著性差异, 且2个生长季均表现相同的调控趋势(图6)。与之相比, 生长季田间无灌溉显著降低了叶片水势, 但COR处理显著提高了该条件下叶片水势, 且2个生长季均表现出相同的COR调控效应。
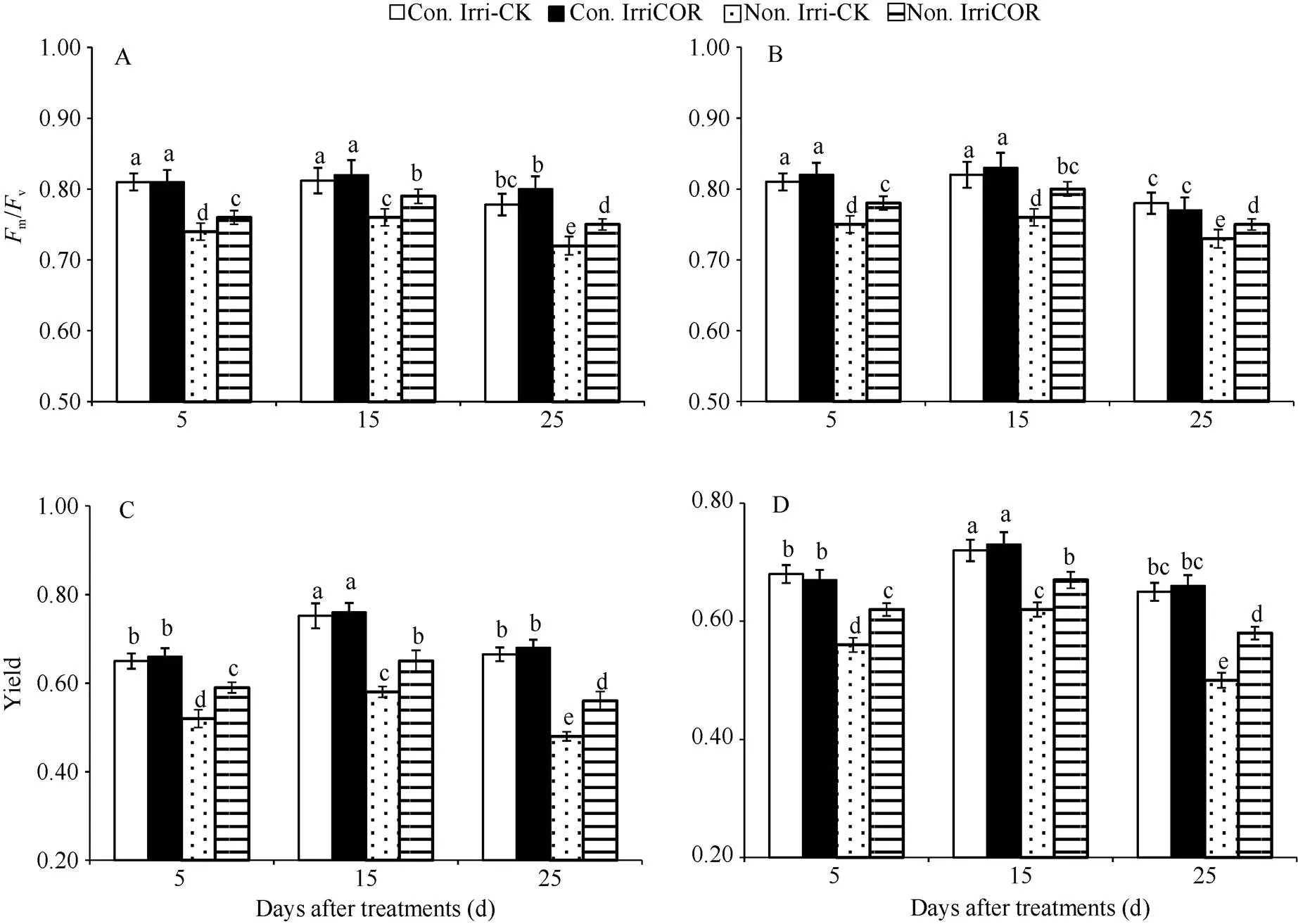
图3 不同田间灌溉条件下COR对大豆叶片荧光参数的影响
A和B分别表示2015年和2016年生长季大豆叶片PSII最大量子产量(v/m); C和D分别表示2015年和2016年生长季大豆叶片PSII实际量子产量(Yield)。
A and B indicate the maximal quantum yield of PSII (v/m) of soybean leaves during 2015 and 2016 growing seasons respectively. C and D indicate the actual photochemical efficiency of PSII (Yield) in soybean leaves during 2015 and 2016 growing seasons respectively.
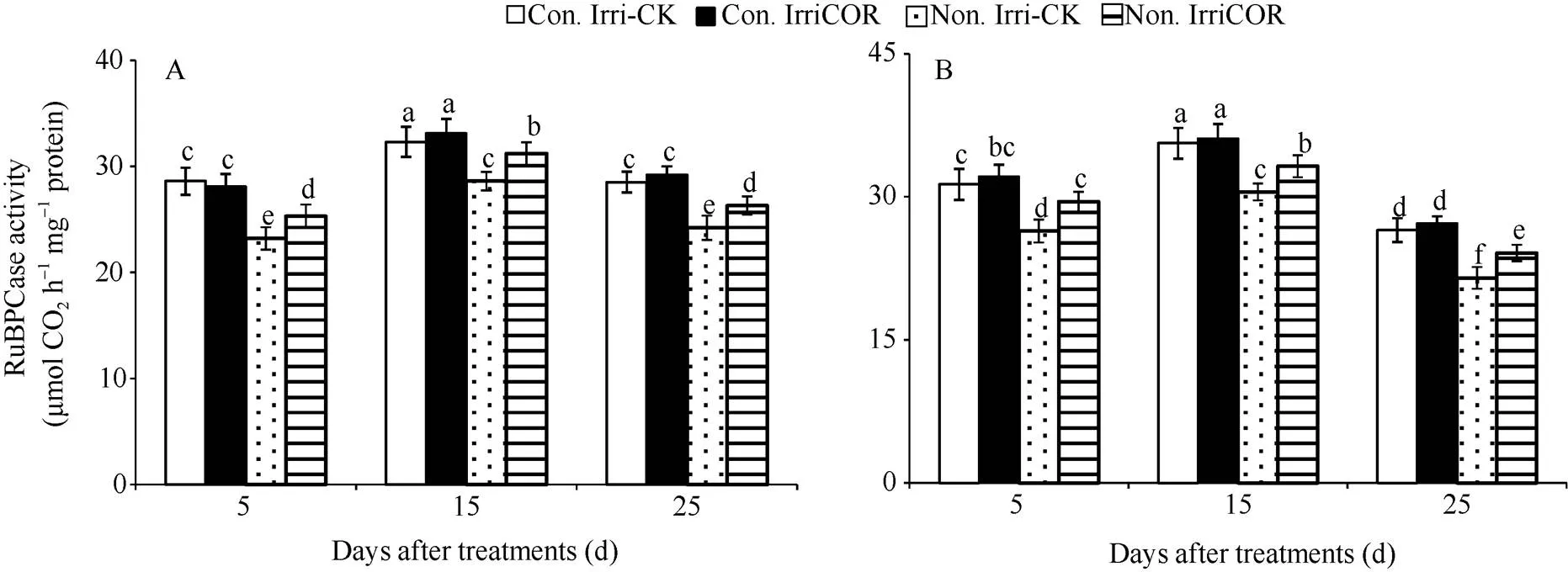
图4 不同田间灌溉条件下COR对叶片RuBP羧化酶活性的影响
A和B分别表示2015年和2016年生长季大豆叶片RuBP羧化酶活性的影响。
A and B indicate the activities of RuBPcase of soybean leaves during 2015 and 2016 growing seasons respectively.
3 讨论
大豆对水分胁迫较为敏感, 如果苗期发生干旱会导致减产20%, 而开花期发生干旱, 产量会减少40%以上[1]。由于我国大豆种植区存在水资源不足或季节降雨量分布不均的问题, 季节性干旱和间歇式干旱频繁发生, 限制了大豆产量。2年的试验结果表明, 与正常灌溉相比, 雨养条件下大豆的单株荚数、粒数和百粒重显著下降, 导致产量显著降低。大豆生长季降雨量会显著调节产量水平, 如2016年大豆生育期降雨量显著高于2015年, 其产量也存在显著差异。这表明水分供给不足或干旱是大豆产量增加的主要限制因素之一[15-16]。同时发现, 在正常灌溉条件下, COR处理对植株性状、产量构成因素与产量均未表现出显著调控效应, 但在生长季无灌溉的条件下, COR会显著提高单株生物量, 改善单株荚数、粒数和百粒重, 增加产量。COR在2种条件下表现出的调控效应差异表明, COR的生物学功能主要是作为抗逆诱导剂, 对作物抗逆具有很好的调控效应。植物通过渗透调节抵御逆境。在植物遭遇干旱时, 代谢活动增强使细胞内溶质浓度提高, 从而降低其渗透势, 进一步降低水势, 保持一定的膨压, 维持正常的生理生化代谢过程[17]。COR能够提高大豆叶片中丙二醛和脯氨酸的含量, 降低叶片水势, 以此适应水分胁迫。此外, COR能够提高干旱条件下大豆叶片中抗氧化物酶SOD、POD、CAT和APX的活性, 这与在花椰菜、水稻和大豆等作物[5,9-10]上的研究结果一致。
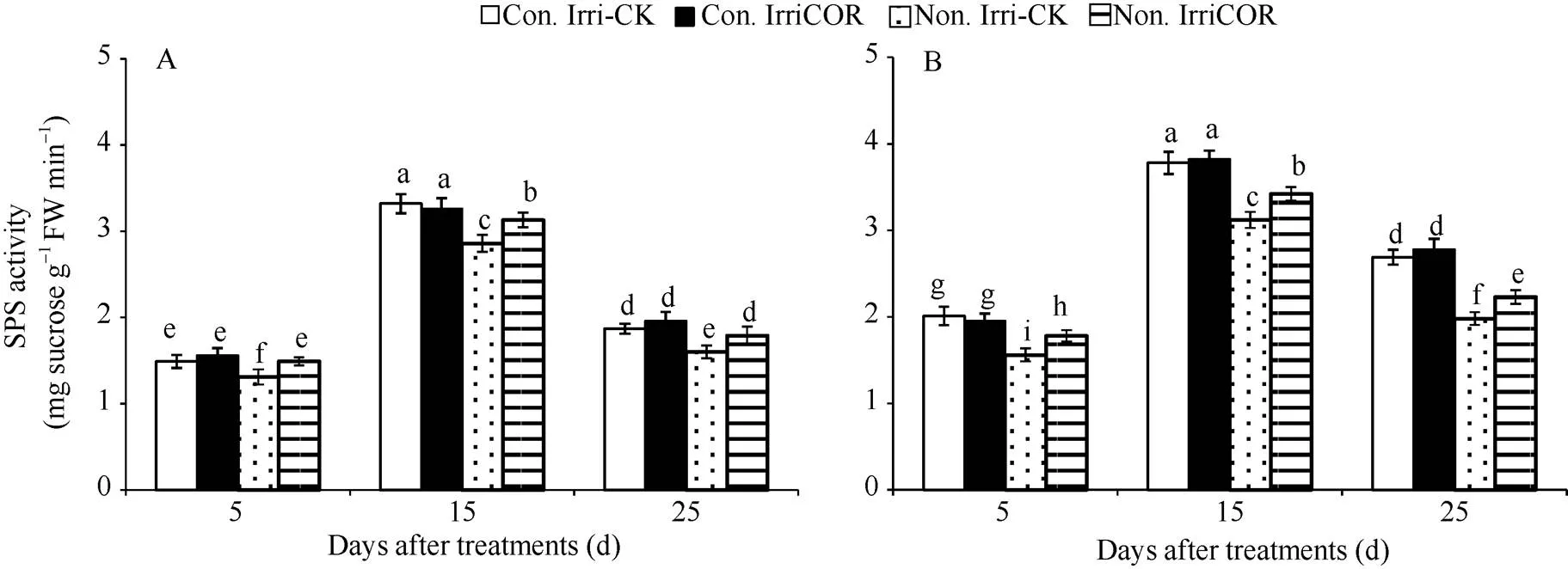
图5 不同田间灌溉条件下COR对叶片蔗糖磷酸合成酶活性的影响
A和B分别表示2015年和2016年生长季大豆叶片SPS蔗糖磷酸化合成酶活性的影响。
A and B indicate the activities of SPS of soybean leaves during 2015 and 2016 growing seasons, respectively.
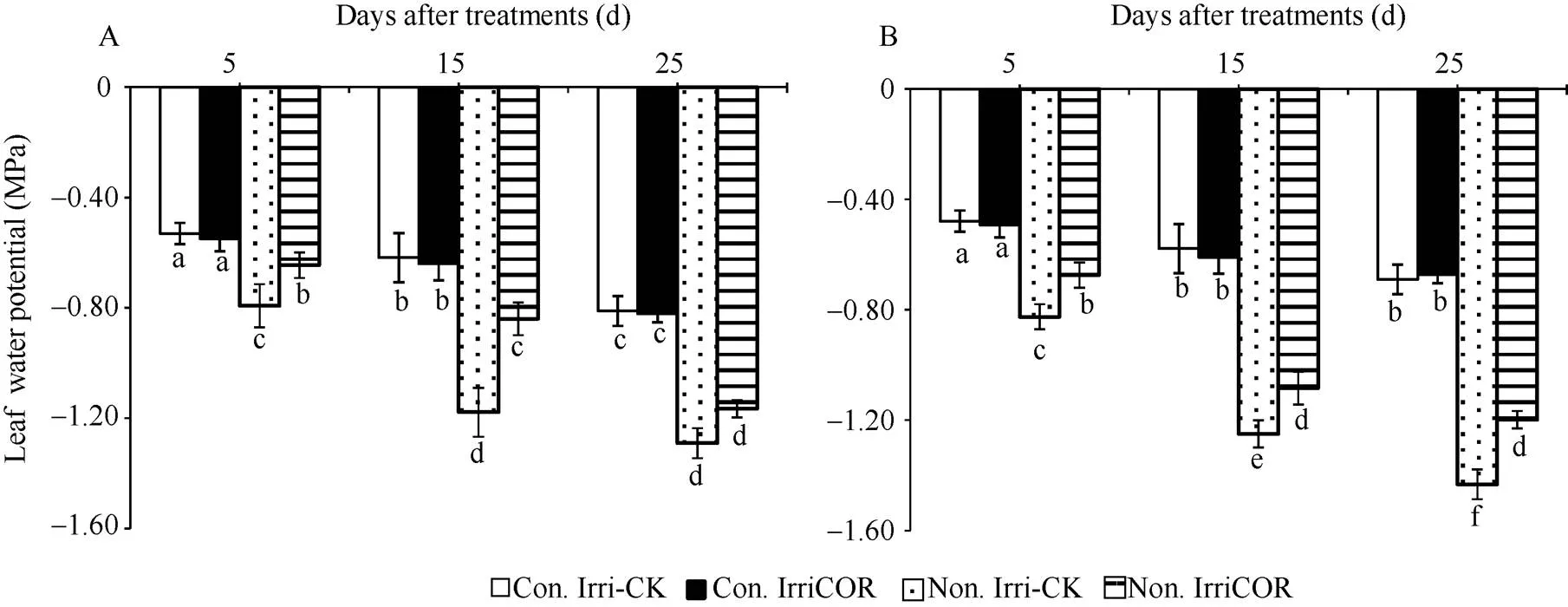
图6 不同田间灌溉条件下COR对叶片水势的影响
A和B分别表示2015年和2016年生长季大豆叶片水势的影响。
A and B indicate the leaf water potential during 2015 and 2016 growing seasons, respectively.
大豆花荚期植株营养生长与生殖生长并进, 对水分与养分供给最为敏感。水分供给不足导致叶片水势、叶绿素含量和光合速率等下降, 促进了花荚脱落, 导致产量降低[18-19]。在生长季无灌溉条件下, COR处理有助于维持大豆叶片较高的水势, 提高叶绿素含量和光合速率, 这与在水稻和花椰菜等的研究结果相一致[5,9]。一般情况下, 水分胁迫会降低大豆植株叶绿素含量, 影响PSII, 导致电子传递速率显著降低[20]。叶绿素荧光可以反映类囊体膜结构与功能变化, 通常被用来表征干旱胁迫下植物叶片光合性能[21]。同时, 叶绿素荧光可以表示PSII使用叶绿素吸收能量和过度的光能量对PSII损坏[22]。正如预期的那样, COR处理显著提高了生长季无灌溉条件下大豆叶片v/m和Yield, 从而提高电子传递速率, 促进物质同化。v/m被用于检测逆境对光合器官的损坏程度[21], 较高的v/m表明COR改善水分胁迫下光合器官功能和维持较高PSⅡ光化学最大量子效率。
RuBP羧化酶是C3植物光合作用过程的关键酶, 而且RuBP羧化酶活性与大豆籽粒产量呈正相关[23], 但干旱胁迫显著降低叶片RuBP羧化酶活性[24]。王磊等[25]研究表明, 在大豆不同生育时期进行干旱处理均降低叶片中RuBP羧化酶活性, 其中开花期干旱降低幅度最大, 其次是鼓粒期和分枝期。本研究发现, COR处理显著提高生长季无灌溉条件下大豆花后叶片RuBP羧化酶活性, 这一结果与COR提高叶片光合速率和叶绿素荧光参数相一致。同时, COR 处理也显著提高生长季无灌溉条件下大豆花后叶片SPS活性, 而叶片SPS活性高低能反映籽粒对同化产物的需求程度[26]。此外, COR处理显著提高干旱胁迫下14C同化产物从叶片中输出, 提高单株生物量与产量[10], 这一研究结果也为本研究提供了旁证。
4 结论
COR对雨养大豆的产量形成具有显著调控效应。在正常灌溉条件下, COR处理对大豆植株性状、产量及其构成因素、水分状况、光合特征等影响较小。但在雨养条件下, COR处理会显著提高大豆开花后叶片水势、叶绿素含量、光合速率和叶绿素荧光参数, 增加叶片RuBP羧化酶和SPS活性, 从而改善大豆产量构成因素, 促进籽粒产量增加。
[1] Manavalan L P, Guttikonda S K, Tran L P, Nguyen H T. Physiological and molecular approaches to improve drought resistance in soybean., 2009, 50: 1260–1276.
[2] Anjum S A, Wang L, Farooq M, Khan I, Xue L. Methyl jasmonate-induced alteration in lipid peroxidation, antioxidative defence system and yield in soybean under drought., 2011, 197: 296–301.
[3] Zhang M C, Duan L S, Tian X L, Wang B, Li Z. Uniconazole-induced tolerance of soybean to water deficit stress in relation to changes in photosynthesis, hormones and antioxidant system., 2007, 164: 709–717.
[4] Zhang M C, Zhai Z X, Tian X L, Duan L S, Li Z H. Brassinolide alleviated the adverse effect of water deficits on photosynthesis and the antioxidant of soybean (L.)., 2008, 56: 257–264.
[5] Ai L, Li Z H, Xie Z X, Tian X L, Eneji A E, Duan L S. Coronatine alleviates polyethylene glycol-induced water stress in two rice (L.) cultivars., 2008, 194: 360–368.
[6] Zhou Y Y, Zhang M C, Li J M, Li Z H, Tian X L, Duan L S. Phytotoxin coronatine enhances heat tolerance via maintaining photosynthetic performance in wheat based on Electrophoresis and TOF-MS analysis., 2015, 5: 1–13.
[7] Wang B M, Li Z H, Eneji A E, Tian X L, Zhai Z X, Li J M, Duan L S. Effects of coronatine on growth, gas exchange traits, chlorophyll content, antioxidant enzymes and lipid peroxidation in maize (L.) seedlings under simulated drought stress., 2008, 11: 283–290.
[8] Xie Z X, Duan L S, Tian X L, Wang B M, Eneji A E, Li Z H. Coronatine alleviates salinity stress in cotton by improving the antioxidative defense system and radical scavenging activity., 2008, 165: 375–384.
[9] Wu H L, Wu X L, Li Z H, Duan L S, Zhang M C. Physiological evaluation of drought stress tolerance and recovery in Cauliflower (L.) seedlings treated with methyl jasmonate and coronatine., 2012, 31: 113–123.
[10] Hao L, Wang Y Q, Zhang J C, Xie Y, Zhang M C, Duan L S, Li Z H. Coronatine enhances drought tolerance via improving antioxidative capacity to maintaining higher photosynthetic performance in soybean., 2013, 210: 1–9.
[11] 王玉琼. 冠菌素提高大豆抗旱性的生理机制研究. 中国农业大学硕士学位论文, 北京, 2015. Wang Y Q. Physiological Mechanism on the Coronatine Improved Drought Tolerance in Soybean. Master’s Degree Thesis of China Agricultural University, Beijing, China, 2015.
[12] 郑伟, 谢甫绨, 郭泰, 王志新, 李灿东, 张振宇, 吴秀红, 张茂明, 王庆胜. 密度对不同类型大豆叶部性状的影响. 中国油料作物学报, 2014, 36: 66–70. Zheng W, Xie F T, Guo T, Wang Z X, Li C D, Zhang Z Y, Wu X H, Zhang M M, Wang Q S. Effect of density for different types of leaf traits on soybean., 2014, 36: 66–70 (in Chinese with English abstract).
[13] 顾万荣, 李召虎, 翟志席, 段留生, 张明才. DCPTA和DTA-6对大豆叶片光合及叶绿素荧光特性的调控. 大豆科学, 2008, 27: 777–782. Gu W R, Li Z H, Zhai Z X, Duan L S, Zhang M C. Regulation of DCPTA and DTA-6 on photosynthesis and chlorophyll fluorescence parameters of soybean leaves., 2008, 27: 777–782 (in Chinese with English abstract).
[14] Huber S C. Role of sucrose phosphate synthetase in partition of carbon in levels., 1983, 71: 818–821.
[15] Cooper R L, Fausey N R, Streeter J G. Yield potential of soybean grown under a subirrigation/drainage water management system., 1991, 83: 884–887.
[16] 霍建玲, 邢雪莹, 杨雪, 杨艳玲, 周静文, 魏彬, 喻金秋, 王昕奕, 柏锡. 干旱对黑龙江省大豆品种农艺性状的影响. 分子植物育种, 2018, 16: 1668–1676. Huo J L, Xing X Y, Yang X, Yang Y L, Zhou J W, Wei B, Yu J Q, Wang X Y, Bai X. Effects of drought stress on agronomic traits of soybean cultivars from Heilongjiang province., 2018, 16: 1668–1676 (in Chinese with English abstract).
[17] 王邦锡, 黄久常, 王辉. 不同植物在水分胁迫条件下脯氨酸的累积与抗旱性的关系, 植物生理学报, 1989, 15: 46–51. Wang B X, Huang J C, Wang H. Relationship between accumulation of proline and drought resistance of different plants under water stress conditions., 1989, 15: 46–51.
[18] Liu F L, Christian R J, Mathias N A. Drought stress effect on carbohydrate concentration in soybean leaves and pods during early reproductive development: its implication in altering pod set., 2004, 86: 1–13.
[19] Liu F L, Andersen M N, Jacobsen S E, Jensen C R. Stomatal control and water use efficiency of soybean (L. Merr.) during progressive soil drying., 2005, 54: 33–40.
[20] 高小丽, 孙健敏, 高金锋, 冯佰利, 柴岩, 贾志宽. 不同基因型绿豆叶片光合性能研究. 作物学报, 2007, 33: 1154–1161. Gao X L, Sun J M, Gao J F, Feng B L, Chai Y, Jia Z K. Photosynthetic performance in the leaves of different mung bean genotypes., 2007, 33: 1154–1161 (in Chinese with English abstract).
[21] Baker N R, Rosenqvis E. Applications of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities., 2004, 55: 1607–1621.
[22] Maxwell K, Johnson G N. Chlorophyll fluorescence: a practical guide., 2000, 345: 659–668.
[23] 郝乃斌, 戈巧英, 张玉竹, 徐继, 张国铮, 谭克辉, 杜维广, 张桂茹, 栾晓燕, 黄承运. 高光效大豆光合特征的研究. 大豆科学, 1989, 8: 283–287. Hao N B, Ge Q Y, Zhang Y Z, Xu J, Zhang G Z, Tan K, Du W G, Zhang G R, Luan X Y, Huang C Y. Studies on the photosynthetic properties of soybean with photosynthetic efficiency., 1989, 8: 283–287 (in Chinese with English abstract).
[24] 张明才, 何钟佩, 田晓莉, 段留生, 王保民, 翟志席, 董学会, 李召虎. SHK-6对干旱胁迫下大豆叶片生理功能的作用. 作物学报, 2005, 31: 1215–1220.Zhang M C, He Z P, Tian X L, Duan L S, Wang B M, Zhai Z X, Dong X H, Li Z H. Effects of plant growth regulator SHK-6 on physiological function of soybean leaves under water deficiency., 2005, 31: 1215–1220 (in Chinese with English abstract).
[25] 王磊, 王鹏程, 张彤, 张恒月, 丁圣彦. 结荚期短期干旱和复水对大豆叶片光合和产量的影响. 生态学报, 2009, 29: 3328–3332. Wang L, Wang P C, Zhang T, Zhang H Y, Ding S Y. Effect of short-term drought and rewatering during the pod-setting stage on leaf photosynthesis and yield of the soybean., 2009, 29: 3328–3332 (in Chinese with English abstract).
[26] Sarquı́s J I, Gonzalez H, de Jiménez E S, Dunlap J R. Physiological traits associated with mass selection for improved yield in a maize population., 1998, 56: 239–246.
Regulatory effects of coronatine on photosynthetic characteristics and yield of soybean under different irrigation conditions
YU Hai-Yue, YAN Yan, ZHANG Yu-Shi, ZHANG Ming-Cai*, and LI Zhao-Hu
Engineering Research Center of Plant Growth Regulator, Ministry of Education / College of Agronomy and Biotechnology, China Agricultural University, Beijing 100093, China
Drought is one of the important factors limiting the high and stable yield of soybean, and an important problem restricting the sustainable production of soybean in China. Using biological regulators to improve the drought tolerance of soybean in production is a new feasible biological water-saving mode. In this study, the field experiments were conducted to investigate the effects of coronatine (COR) on leaf water potential, photosynthetic characteristics, plant traits, yield and its components of soybean under conventional irrigation and no irrigation conditions from 2015 to 2016. Under conventional irrigation conditions, COR could not change the values of leaf water potential, chlorophyll content, photosynthetic rate, chlorophyll fluorescence parameters, and activities of RuBP carboxylase and SPS in soybean leaves compared to control. And there was no significant difference in yield and biomass between COR treatment and control. However, under no irrigation conditions, COR significantly increased the leaf water potential, chlorophyll content, photosynthetic rate and chlorophyll fluorescence parameters, as well as the activities of RuBP carboxylase and SPS of soybean leaves, thus improved the yield and yield components. These results suggest that COR has a positive regulatory effect on photosynthetic characteristics and yield formation of soybean under no irrigation conditions, and can be used as a new technical means for cultivation preventing drought resistance in soybean production.
coronatine; irrigation; soybean; photosynthetic characteristics; yield
本研究由国家自然科学基金项目(31471420)资助。
This study was supported by the National Natural Science Foundation of China (31471420).
张明才, E-mail: zmc1214@163.com, Tel: 010-62733049
E-mail: yuhaiyueo@126.com
2019-03-05;
2019-06-12;
2019-07-08.
URL:http://kns.cnki.net/kcms/detail/11.1809.s.20190705.1204.002.html
10.3724/SP.J.1006.2019.94035
