‘西伯利亚’百合丙二烯氧化物环化酶基因LsAOC的克隆及其表达分析
2019-11-04付宇辰闫子飞胡增辉冷平生
付宇辰,吴 琦,闫子飞,胡增辉,冷平生
(北京农学院 园林学院//北京林果业生态环境功能提升协同创新中心,北京 102206)
茉莉酸类物质(Jasmonates,JAs)在植物体内发挥着各项生理和信号分子的作用[1]。茉莉酸(JA)在植物体内是由亚麻酸氧化合成的[2]。亚麻酸被细胞膜释放起始,茉莉酸的形成还需要经过脂氧合酶(Lipoxygenase,LOX)、丙二烯氧化合酶(Allen oxide synthase,AOS)、丙二烯氧化物环化酶(Allene oxide cyclase,AOC)、12-氧代植二烯酸还原酶(12-oxophytodienoate reductase,OPR)等关键酶催化与一系列的β氧化过程。AOC基因是茉莉酸合成途径中的关键基因,其表达量直接影响植物体内合成JA的含量。AOC可以将AOS的产物丙二烯氧化物(12,13(s)-epoxy-(9Z,11E,15Z)-octadecatrienoic acid, 12,13-EOT)特异性催化并生成1,2-氧-植物二烯酸(cis-(+)-12-oxo-phytodienoic acid,OPDA)产物,在α-亚麻酸代谢通路中起到了关键作用[3]。同时AOC基因也能参与脱落酸(Abscisic acid,ABA)信号途径,通过增加其活性使植物的耐盐性增强[4]。
百合(Liliumspp.)是著名的五大切花之一,‘西伯利亚’百合是市面上最常见的百合品种,也东方百合种系中的经典代表。‘西伯利亚’百合花朵全为白色,花被片硕大,花香浓郁,有很高的观赏价值,是日常生活中常见的百合切花品种之一[5]。本研究拟通过分子生物学手段,使用RACE技术在 ‘西伯利亚’百合中克隆得到LsAOC基因,并进行生物信息学分析与时空表达分析,初步探究在花香的形成及释放过程中茉莉酸信号途径所起的作用,也为百合花香育种及应用提供理论基础。
1 材料和方法
1.1 材料与采样方法
东方百合‘西伯利亚’种球,用等比例草炭与蛭石为基质种植在北京农学院园林植物实践基地日光温室内。温室温度25~28 ℃,相对湿度55%~65%,通风良好,肥料充足。采集花蕾期、半开期、盛开期、衰败期4个不同时期的花被片,内瓣、外瓣各取1片为1组用铝箔纸包裹,暂存于液氮中。此外以相同方法另取盛开期百合的根、茎、叶、内花被片、外花被片、子房、花柱、花丝、花药共9组不同组织样品,液氮暂存后在-80 ℃冰箱内储存。
1.2 试验方法
1.2.1 总RNA提取与反转录 使用北京全式金生物技术有限公司的RNA提取试剂盒提取‘西伯利亚’百合总RNA,通过琼脂糖电泳检测及估算样品RNA浓度,再使用TARAKA公司的反转录试剂盒将样品总RNA反转录成cDNA备用。方法参照2试剂盒说明书。
1.2.2LsAOC基因全长克隆 通过查找转录组数据库中的资料得到基因序列片段,根据此片段在Primer3网页(http://primer3.ut.ee)设计引物LsAOC-ZJF和LsAOC-ZJR。以反转录得到的cDNA为模板将此引物进行PCR扩增,产物经过纯化回收、连接转化、鉴定测序和比对分析得到了LsAOC基因中间片段,长度约为350 bp。后根据中间片段测序结果使用Primer3网页设计5’端引物LsAOC-5和3’端引物LsAOC-3-1和LsAOC-3-2,分别进行5’端与3’端RACE克隆,得到约700 bp的5’端片段和约350bp的3’端片段,测序并使用DNAMAN软件将测序结果拼接,即得到LsAOC基因全长序列。根据LsAOC基因全长序列设计全长引物LsAOC-QCF和LsAOC-QCR,PCR扩增后产物长度与拼接结果相符,测序验证为全长序列。
1.2.3LsAOC基因相对表达量分析 提取‘西伯利亚’百合4个开花时期和9个不同组织RNA,根据LsAOC基因全长序列设计荧光定量引物LsAOC-YGF和LsAOC-YGR,选取β-Actin-F和β-Actin-R为内参引物。反转录得到各组cDNA并以其为模板进行荧光定量PCR,测定和记录Ct值。使用WPS表格统计并分析LsAOC基因相对表达量。

表1 引物序列Tab.1 Primer sequences
2 结果与分析
2.1 LsAOC基因生物信息学分析
2.1.1LsAOC基因序列及蛋白同源性分析LsAOC基因CDS全长741bp,用DNAMAN软件将其翻译成氨基酸序列,该序列编码246个氨基酸残基(图1),输入至NCBI网站(https://www.ncbi.nlm.nih.gov/)发现此序列属于Allene_ox_cyc 超基因家族,其中第22~246号氨基酸为PLN02343特异性位点。
使用NCBI网站的BLAST功能对LsAOC基因编码的蛋白序列进行在线比对,下载比对结果中与此序列相似性较高的10种其他植物的AOC氨基酸序列。用DNAMAN软件的多序列比对功能比对后发现,‘西伯利亚’百合AOC蛋白序列与麝香百合(Liliumlongiflorum, AET50928.1)和岷江百合(Liliumregale, ART33463.1)相似性很高,可达93%;与赤豆(Vignaangularis, XP_017427281.1)的相似性稍次之,为77%;大多数植物如:木豆(Cajanuscajan, XP_020226154.1)、蔓花生(Arachisduranensis, XP_015967605.1)、牵牛花(Ipomoeanil, XP_019173088.1)和胡杨(Populuseuphratica, XP_011004427.1)等与‘西伯利亚’百合的相似性均为76%。与凤梨(Ananascomosus, XP_020106214.1)、豌豆(Pisumsativum, BAE45342.1)和大豆(Glycinemax, NP_001304386.1)的相似度在74%以上(图2)。

图1 LsAOC基因序列及其编码的蛋白序列Fig.1 The gene sequence and deduced amino acid sequence of LsAOC
2.1.2 ‘西伯利亚’百合LsAOC编码蛋白的理化性质分析及结构预测 通过ExPASy网站ProtParam和Compute pI/Mw工具预测得到,LsAOC蛋白呈碱性,共有21个负电荷氨基酸残基和23个正电荷氨基酸残基,以亮氨酸、脯氨酸、丝氨酸、苯丙氨酸为氨基酸主要组成成分。其理论等电点为8.44,脂肪系数为82.80,相对分子质量27.52 kD。LsAOC蛋白属于不稳定蛋白,不稳定系数为59.57;其亦为亲水性蛋白,平均亲水指数为-0.217。通过 ExPASy 网站的GOR4工具预测得到,LsAOC蛋白二级结构,其中Cc包含178个氨基酸残基,占氨基酸残基总数的72.36%;Ee包含46个氨基酸残基,占18.70%;Hh包含22个氨基酸残基,占总氨基酸残基8.94%。利用Swiss-model 程序构建了LsAOC蛋白的三维立体结构模型。以ALLENE OXIDE CYCLASE 2(4cq6.1.A)为模板,相似性达68.79%(图3)。
2.2 LsAOC基因表达分析结果
在‘西伯利亚’百合4个不同花期中,LsAOC基因的相对表达量变化呈现出一定的规律。LsAOC基因在花蕾期、半开期和衰败期表达量基本一致,而在盛开期有明显的下降,约为其他3个时期表达量的3/5(图4)。在‘西伯利亚’百合的9个不同组织器官中,表达情况也存在较大差异。内瓣和叶片中LsAOC基因的表达量均高于其他组织,子房和外瓣中的表达量基本一致,约为内瓣的2/3。花柱、花丝和花药的表达处于同一水平上,根中的表达量最低,约为内瓣的17%(图4)。

图2 ‘西伯利亚’百合LsAOC基因编码的氨基酸序列与其他植物AOC氨基酸序列的同源性比对Fig.2 The homology of LsAOC coding protein between Lilium ‘Siberia’ and other plants
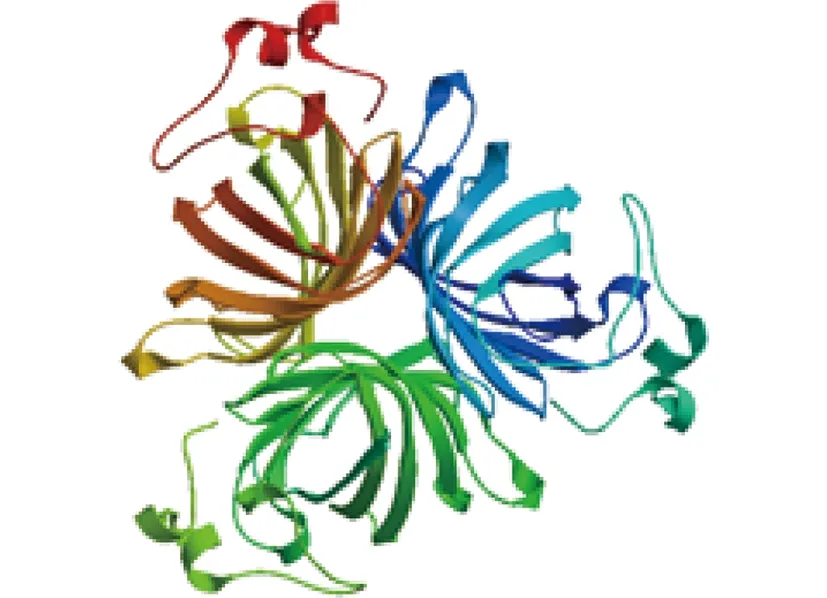
图3 LsAOC基因编码蛋白三级结构预测Fig.3 Prediction of tertiary structure of LsAOC coding protein
3 讨 论
丙二烯氧化物环化酶可以将经丙二烯氧化合酶催化得到的不稳定12,13-EOT重新排列,形成外消旋的12-OPDA。因为只有顺式异构体cis-(+)-12-O-OPDA才能进一步转化形成天然的茉莉酸,所以AOC基因的作用至关作用。目前AOC基因已在拟南芥 (Arabidopsisthaliana,AtAOC)、白木香(Aquilariasinensis,AsAOC)、蕙兰(CymbidiumfaberiRolfe,CfAOC)、大豆(Glycinemax,GmAOC)、金鱼草(Antirrhinummajus,AmAOC)等多种植物中克隆获得。
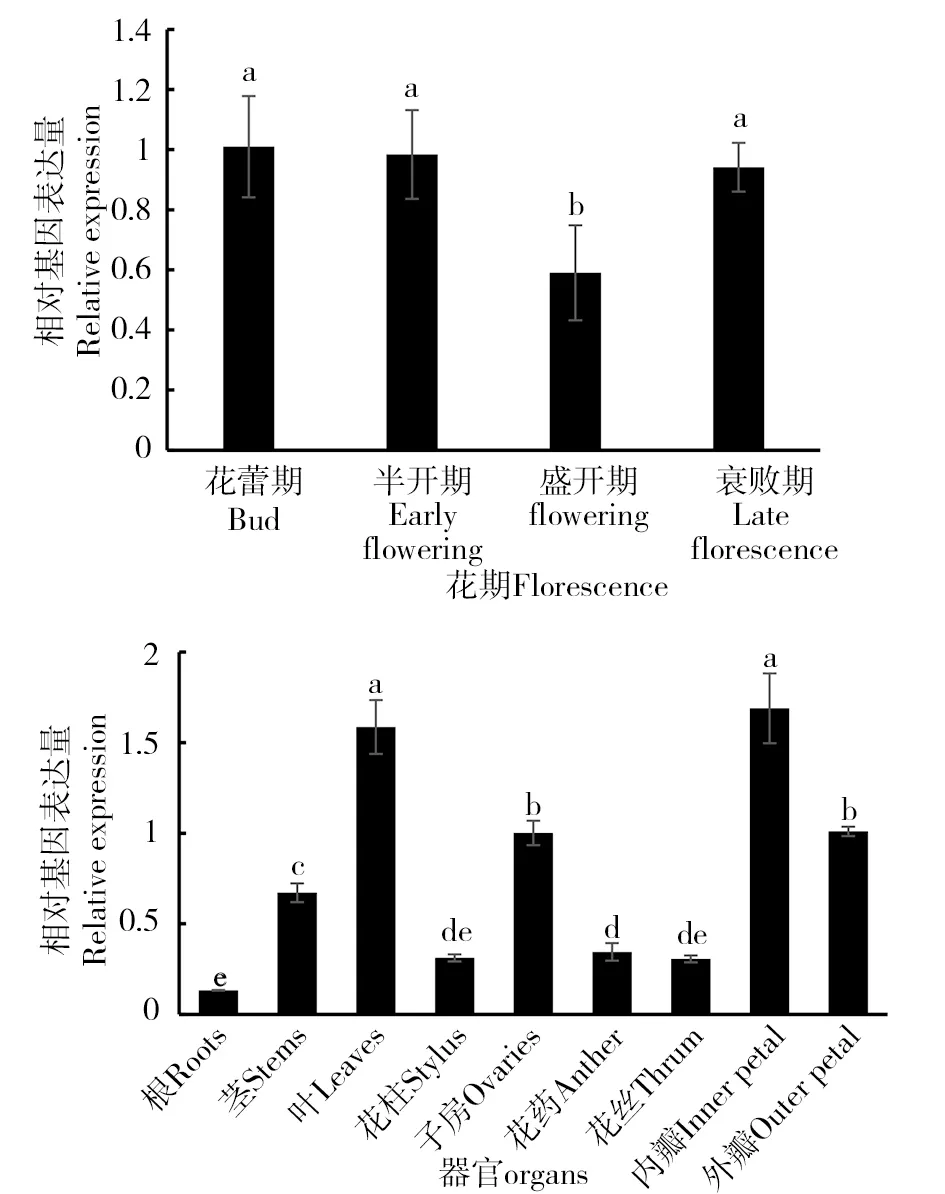
图4 不同花期和不同组织中的LsAOC基因的表达水平Fig.4 The LsAOC expression levelsat different flowering stages,in different tissues
本研究使用RACE法在‘西伯利亚’百合中克隆得到了LsAOC基因全长,得知LsAOC基因CDS全长741 bp,编码246个氨基酸残基,相对分子质量Mw=27.52 kD,呈碱性且不稳定,亲水性好,从属Allene_ox_cyc超基因家族。其蛋白序列与其他百合品种麝香百合和岷江百合极为相似。
‘西伯利亚’百合LsAOC基因在不同花期和不同组织部位中的表达呈现出一定的规律。首先,LsAOC基因在花蕾期、半开期和衰败期表达量基本一致,高于盛开期。在金鱼草中,AmAOC基因与本研究结果相似,相对表达量亦在盛花期最低,败花期又升高[6]。由此分析,AOC基因可能在花朵发育前期发挥作用,通过调节JA合成调控植物生长。而AOC基因作为JA合成途径关键基因,衰败期LsAOC基因相对表达量的升高可能是与衰败期植物生长类激素的累积有关,因为JAs信号参与了植物的发育与代谢过程。在九个不同组织器官中,表达情况也存在较大差异。内瓣和叶片相对表达量最高,其次是子房和外瓣,根的相对表达量则最低。内瓣和叶片的相对表达量显著高于其他组织,与周银在蕙兰不同组织中测定CfAOC基因相对表达量结果一致[7]。此外AOC基因在不同的物种、不同的AOC基因间存在一定的表达差异。枸杞AOC基因在花和茎中相对表达量较高,在叶和根中相对低[8]。在白香木中,AOC基因在茎中表达量最高, 根与茎尖次之, 叶中表达量最低[9]。在大麦中AOC表达在叶基,盾片节点和根尖相对较高[10]。吴倩在大豆AOC家族中对6个基因进行了组织特异性检测,发现GmAOC2和GmAOC5分别在根和茎中表达量较高,而GmAOC3则在叶片中表现突出,还有两个基因在所有组织中表达均很微弱[11]。不同植物AOC基因在不同组织中相对表达量的高低存在差异,可能是由于JAs的合成调控方式在不同物种中各不相同,尤其在单子叶植物和双子叶植物之间,这种差异存在着明显的不同[10]。
丙二烯氧化物环化酶是JA合成代谢途径中的重要限速酶,调控着植物的生命活动进程,具有研究价值和应用前景,但AOC基因有关花香释放的研究成果并不充分,缺少更多试验数据支撑,因此还有需要进一步深入探究。本研究结果对LsAOC基因在‘西伯利亚’百合中的表达提供了数据及理论基础,具体应用方向还需要进一步探索。
