硫化物对黑臭河道底泥反硝化潜势的影响作用研究
2019-10-28汪珊朱瑾何岩黄民生周运昌
汪珊 朱瑾 何岩 黄民生 周运昌

摘要:通过探究不同浓度硫化物对黑臭河道底泥反硝化过程的影响,同时分析底泥细菌、反硝化菌和硫酸盐还原菌的响应变化,为强化底泥反硝化脱氮提供理论依据与技术支撑.研究结果表明:较低浓度的硫化物(8 mg.L-1)对底泥反硝化潜势无明显影响;适宜浓度的硫化物(40和64 mg.L-1)对底泥反硝化有明显的促进作用,且浓度越高促进作用越明显;当硫化物浓度升高到96 mg.L-1及以上时,还原态硫对反硝化过程起抑制作用,浓度越高抑制作用越明显.底泥经过一段时间的反硝化培养,细菌多样性以变形菌门、绿弯菌门、拟杆菌门为主;同时,底泥细菌总数明显增加,代谢菌群的nirS丰度比、dsrB丰度比分别为1.42%和0.05%,相较原始底泥(0.15%,0.19%),反硝化细菌增值明显,但硫酸盐还原菌数量有所下降.
关键词:黑臭河道;还原态硫;反硝化;底泥
中图分类号:X522
文献标志码:A
DOI: 10.3969/j.issn.1000-5641.2019.04.015
0 引言
隨着黑臭河道整治工作的相继开展,河道黑臭现象得到了有效控制,但总氮去除率偏低问题逐渐凸显,氮营养盐难以去除成为黑臭河道治理过程中亟待解决的突出问题.硫元素是河道生态系统的重要元素,由于水体中硫化物的高度不稳定性,其对氮素削减过程的影响作用往往被屏蔽.此外,现阶段关于硫对反硝化作用的研究还存在分歧.一方面,硫化物可能耦合硝酸盐还原过程,促进硫自养反硝化的发生[1-2]:另一方面,高浓度的硫化物会通过抑制一氧化氮(NO)和一氧化二氮(N20)还原酶的活性来阻碍反硝化过程的进行[3-4],但尚不明确还原态硫对反硝化影响作用的浓度限值.因此,探究硫化物对黑臭河道底泥反硝化潜势的影响机制可为强化底泥反硝化脱氮提供理论依据.
1 材料与方法
1.1 研究对象
本实验所需的河道底泥和上覆水均采集于上海市桃浦区里店浦,该河为上海市工业河的一条支流,受沿岸排污口影响不断纳入污水,污染情况严重,河水的水动力条件较差,自净能力较弱,是典型的城市黑臭河道.采样河道的位置如下图1所示,
里店浦底泥的基本理化性质:底泥酸可挥发硫化物(AVS)含量为122.7 μmol.g-1,底泥总铁含量为19.7 9.kg-1.
1.2 实验装置
将取回的底泥置于500 mL血清瓶中,加入基础培养液和微量元素溶液[11],河道底泥与液体培养基比例为1:10,添加相应浓度的硫化物(Na2S.9H20,分析纯)和硝酸盐氮(KN03,分析纯),密封遮光处理后置于30。C恒温箱中培养.
为探究不同硫化物浓度对自养反硝化的影响,设置实验组1 5(见表1),分别设置硫化物溶液为0、8、40、64和96 mg.L.1进行反硝化培养实验(此前预实验已知8和40mg.L.1对反硝化有促进作用,96 mg.L-1起抑制作用).
在培养期间,定期取样测定NH4-N、N02 -N、N03-N、S2-和S02-的浓度.每当测定的S2-和N02 -N浓度低于检测限时,更换培养液,同时向血清瓶中添加相应基质.
1.3 高通量测序及荧光定量PCR的测定
硫自养反硝化实验各组趋于稳定时,将根据理化指标分析,选取关键实验组中的底泥进行细菌通用引物和基于功能基因16S rDNA、nirS、dsrB的高通量测序分析及荧光定量PCR测定分析,以解析底泥细菌、反硝化和硫酸盐还原菌群丰度及结构的响应变化.高通量测序与荧光定量测定所用引物如下表2所示.
2 结果与讨论
2.1 硫化物浓度对底泥反硝化作用的影响
不同硫化物浓度对反硝化过程的影响不同.图2是比较各实验组硝酸盐的浓度变化,从图上直线的下降水平可以看出,硫化物浓度为8 mg.L-1、40 mg.L-1和64 mg.L-1的实验组硝酸盐还原速率高于不加硫化物的空白对照组,并且硫化物浓度为64 mg.L-1的实验组硝酸盐还原速率最快,其次是40 mg.L-1和8 mg.L-1,即硫化物浓度越高,硝酸盐还原速率越快,且底泥培养体系在10~14 d内硝酸盐的还原率均可达到98%以上.相比不添加硫化物的培养组(对照组),8 mg.L-1的硫化物对反硝化促进作用不明显;40 mg.L-1的硫化物对反硝化有促进作用但在第3个培养周期后才显现出,平均每个培养周期的第6天硝酸盐还原率即可达到98%以上;64 mg.L-1的硫化物对反硝化有明显的促进作用且平均每个培养周期的第3天硝酸盐还原率即可达到98%以上(见表3).实验组5硫化物浓度为96 mg.L-1,硝酸盐的还原速率明显低于空白组,反硝化过程一直被抑制,一个培养周期结束时反应体系内硝酸盐的浓度约为(6.62±0.03)mg.L-1,抑制率约为23.64%;实验组2后期硫化物浓度升高至160 mg.L-1(从第6个培养周期开始),硝酸盐的还原速率明显降低,同时一个培养周期结束时反应体系内硝酸盐的浓度约为(9.52±0.22)mg.L-1,第6~8个培养周期硝酸盐还原抑制率约为34%.
由此可见,较低浓度的硫化物(S2-浓度不高于64 mg.L-1)对反硝化过程起一定的促进作用,且浓度越高促进作用越明显,推测可能出现了硫自养反硝化的发生[7] S2-作为电子供体加速硝酸盐的还原过程:而高浓度硫化物对底泥的反硝化过程有抑制作用,其原因主要是硫化物对一氧化氮(NO)和一氧化二氮(N20)还原酶的活性有抑制从而阻碍的反硝化进程[6].
此外,高浓度硫化物实验组后期出现较明显的氨氮积累(见图3).S2-浓度为96 mg.L-1和160 mg.L-1的底泥培养组氨氮浓度明显高于其他实验组,氨氮积累浓度分别为(3.94±0.16)mg.L-1和(4.46±0.14)mg.L-1,同时亚硝氮积累浓度分别为(4.02±0.15)mg.L-1和(2.82±0.13)mg.L-1,但积累的亚硝氮会在每个培养周期结束时消耗完毕,推测实验中可能存在异化硝酸盐还原过程(Dissimilatory nitrate reduction to ammoumum,DNRA),硫化物浓度较高会抑制反硝化并作为电子供体参与呼吸型DNRA过程[8].
不同硫化物浓度下各实验组硫化物和硫酸盐浓度变化有明显差异(见图3).当S2-浓度小于或等于64 mg.L-1时,一个培养周期结束时硫酸盐SO4一的平均浓度随所添加硫化物浓度的增加而升高.S2-浓度为40 mg.L-1和64 mg.L-1時,每个培养周期结束时SO4一浓度分别为(241.5±2.6)mg.L-1和(306.5土4.4)mg.L-1.S2一浓度升高至96 mg.L-1和160 mg.L-1,硫酸盐浓度却没有升高,推测其原因可能是高浓度硫化物未能促使硫自养反硝化的发生,部分还原态的硫未能通过该反应生成硫酸盐并积累.
2.2 硫影响底泥反硝化过程的关键菌群群落结构及丰度响应
本实验选取实验组3(硫化物浓度为40 mg.L-1,硝酸盐28 mg.L-1)、原始底泥、对照组(未添加硝酸盐,硫化物浓度为40 mg.L-1)来进行高通量及荧光定量PCR分析,实验组3是按硫自养反硝化计量比添加的硫化物浓度,上覆水实验也测得该组反硝化作用得到显著促进,而对照组是为了排除硫化物本身对微生物的影响,对比可得硫自养反硝化作用过程中微生物群落特点.
借助于高通量及荧光定量PCR探究底泥培养实验组3的底泥中与氮硫代谢相关微生物的丰度与结构响应变化,以期望进一步解析硫对底泥反硝化潜力的影响.
2.2.1 底泥氮硫代谢相关菌群结构的响应变化
培养组底泥细菌多样性与对照组有明显差异(见表4).实验组3(硫化物40 mg.L-1)细菌的OTU数、Chaol、Shannon和Simpson指数均低于对照培养组(硫化物40 mg.L-1,未添加硝酸盐).原因可能是实验组发生反硝化作用,微生物以自养细菌为主,而自养细菌世代周期较长、增殖较慢,因此丰富度偏低.
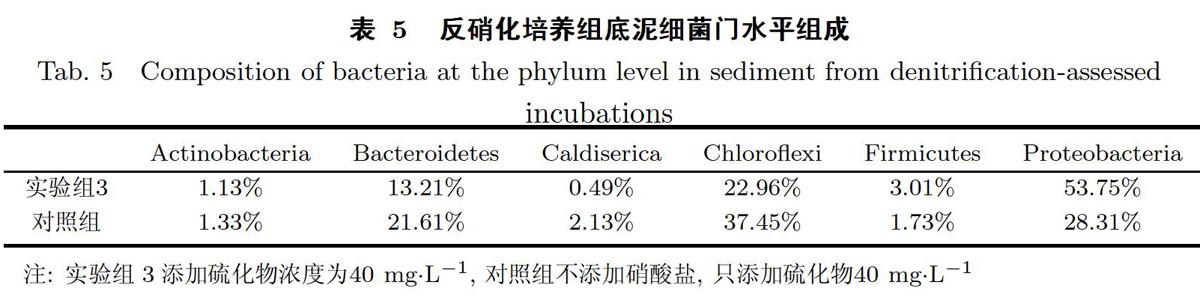
根据底泥结构检测结果(见图4),反硝化培养组底泥细菌多样性以变形菌门(Proteobac-teria)、绿弯菌门(Chloroflexi)、拟杆菌门(Bacteroidetes)、厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)和嗜热丝菌门(Caldiserica)为主;对照组底泥细菌以变形菌门(Proteobacte-ria)、厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)、绿弯菌门(Chloroflexi)和拟杆菌门(Bacteroidetes)为主.变形菌门(Proteobacteria)在实验组3和对照培养组底泥中的相对丰度分别为53.75%和28.31%;绿弯菌门(Chloroflexi)在各培养组底泥中的相对丰度分别为22.96%和37.45%;拟杆菌门(Bacteroidetes)在各培养组底泥中的相对丰度为13.21%和21.61%(见表5).在为期近100 d的培养后,实验组3底泥的变形菌门(Proteobacteria)远远高于对照培养组底泥,分析可能是长时间反硝化潜力的培养使得底泥反硝化细菌(隶属于变形菌门)活性提高且数量增加,而对照培养组培养过程中只添加了硫化物,没有硝酸盐作为电子受体供反硝化细菌利用,因此其反硝化细菌的活性相对较低.
基于功能基因nirS检测,实验组3底泥的优势硫自养反硝化菌为硫杆菌属(Thiobacillus)与硫针菌属(Sulfuritalea)分别占21.5%与4.85%.对照培养组优势硫自养反硝化菌为硫杆菌属(Thiobacillus)与硫针菌属(Sulfuritalea),分别占2.43%和0.805.硫杆菌属(Thiobacillus)是最常见且研究最为广泛的硫自养反硝化菌[27].实验组3底泥中的硫杆菌属( Thiobacillus)的相对丰度(21.5%)远高于对照培养组,表明该体系中硫自养反硝化活性很高,与促进反硝化作用分析一致.另外,帕氏氢噬胞菌(llydrogenophaga)和苯基杆菌属(Phenylobacterium)在该培养组中相对丰度较高,上述菌属的主要功能分别是多环芳烃的降解和石油烃及一些混合烃的降解,推测是培养组中反硝化菌为自养细菌,为其他降解有机物的细菌提供了更适宜的生成空间.对于反硝化细菌,实验组3底泥中主要的反硝化细菌为假单胞菌属(Pseudomonas)与陶厄氏菌属(Thauera)分别占1.78%与14.3%;在只添加硫化物的对照培养组底泥中分别占2.38%和14.6%.实验组底泥的核心反硝化菌属为陶厄氏菌属(Thauera)、假单胞菌(Pseudomonas)、贪铜菌属(Cupriavidus)、丛毛单胞菌属(Comamonas)和副球菌(Paracoccus).
基于功能基因dsrB分析底泥硫酸盐还原菌可知:实验组3底泥中的硫酸盐还原菌主要为脱硫微菌属(Desulfomicrobium)与脱硫肠状菌属(Desulfotomaculum),分别占0.56%与15.3%;在只添加硫化物的对照培养组底泥中分别占1.18%和5.65%.可以看出实验组脱硫肠状菌属含量明显小于对照培养组,推测可能实验组反硝化作用得到促进,反硝化细菌与硫酸盐还原菌竞争激烈,不利于硫酸盐还原,导致该菌属含量明显下降.
2.2.2 底泥氮硫代謝相关菌群丰度的响应变化
在为期近100 d的培养后,底泥细菌总数明显增加.实验组3中的细菌数量由初始的1.06x109 copies/g dry sediment增加至2.6lx109copies/g dry sediment,底泥细菌数量较培养之前的细菌数增加了约1.5倍.表明对河道底泥较长时间反硝化潜力的培养,有利于培养体系底泥中细菌的增殖,细菌数量有大幅度增加,活性也响应会有大幅度增加.
培养后的底泥反硝化细菌数量明显增加,而硫酸盐还原菌数量有所下降,实验组3中的反硝化细菌数量由初始的1.58x106copies/g dry sediment增加至3.7lx107copies/g dry sediment,底泥反硝化细菌数量较培养之前的反硝化细菌数增加了约22倍.硫酸盐还原细菌数量由初始的1.99x106copies/g dry sediment降至1.32x106copies/g dry sediment.根据实验结果(见表6)也可得实验组3底泥代谢菌群与原始底泥的nirS丰度比、dsrB丰度比,相比之下可以看出,底泥在40 mg.L-1硫化物的条件下培养一段时间,nirS丰度比明显上升,而dsrB丰度比则有一定程度下降.上述现象均表明添加40 mg.L-1硫化物培养,有利于硝化细菌的增值,而硫酸盐还原菌的增值受到抑制.硫酸盐还原菌与硝酸盐还原菌共同竞争为电子供体,存在竞争关系,硝酸盐还原菌的增值可能一定程度上抑制硫酸盐还原菌的生长,导致数量下降.周期性地向培养体系内添加硫化物与硝酸盐,有利于硫自养反硝化细菌的生长与富集.
3 结论与展望
本实验探究了硫化物对黑臭河道底泥反硝化作用潜势.当硫化物浓度控制在64 mg.L-1以下时,还原态的硫会对反硝化过程起促进作用,硝酸盐还原率可达98%以上,且S2-浓度越高硝酸盐还原速率越快,反硝化过程越短.当还原硫浓度升高到96 mg.L-1及以上时,还原态硫对反硝化过程起抑制作用,浓度越高抑制作用越明显.同时,添加适宜浓度的硫化物(40 mg.L-1)培养一段时间,细菌总数明显增加,尤其是反硝化细菌的增加量达到原来的22倍,同时一定程度上抑制了硫酸盐还原菌的生长,总体上细菌总数增加,对反硝化过程起促进作用.
实验结果表明当硫化物浓度控制在64 mg.L-1以下会对反硝化作用起促进作用,浓度高于96 mg.L-1对反硝化起抑制作用,但是64~96 mg.L-1之间相应情况未知,需要进一步设置浓度梯度,探究最佳促进浓度和抑制限值,从而更有效地进行硫自养反硝化,提高脱氮除硫效率,为黑臭河道治理工程提供理论依据与技术支撑,缓解城市黑臭河道现象.
[参考文献]
[1] 朱艺双,朱瑾,何岩城市黑臭河道内源硫与硝酸盐异化还原过程的耦合机制研究[C]//2017中国环境科学学会科学与技术年会论文集(第二卷),2017.
[2] sHAo M F,zHANG T,FANG H H.sulfur—driven autotrophic denitrification:Diversity,biochemistry,andenginee“ng applications[J].Applied Microbiology and Biotechnology,2010,88(5):1027一1042.
[3] sENGA Y,MocHIDA K,FuKuMoRI R,et al.N20 accumulation in estua“ne and coa8tal sediments:Theinfluence of H2S on dissimilatory nitrate reduction[J].E8tuarine,Coastal and Shelf Science,2006,67(1/2):231—238.
[4] BowLEs M w,NIGRo L M,TEsKE A P,et al.Denitrification and environmental factors influencing nitrateremoval in Guaymas Basin hydrothermally altered sediments[J].nontiers in Microbiology,2012,3(377):1—11.
[5] 汪建华城市黑臭河道氮转化途径分型表征及微生物作用机理研究[D]上海:华东师范大学,2017.
[6] PLuMMER P,ToBIAs c,cADY D.Nitrogen reduction pathways in estuarine sediment8:Infiuences of organiccarbon and sulfide IJ] Journal of Geophysical Research:Biogeosciences,2015,120(10):1958—1972.
[7] MAHMooD Q,zHENG P,cAI J,et al. Anoxic sulfide biooxidation using nitrite as electron acceptor[J].Journal of Hazardous Materials,2007,147(1/2):249—256.
[8] AN s,GARDNER w s Dis8imilatory nitrate reduction to ammonium(DNRA)as a nitrogen link,versusdenitrification as a sink in a shal10w e8tuary(Laguna Madre/Baffin Bay,Texas[J]Marine Ecology Progres8Series.2002.237:41—50.
[9] JuNcHER J c,JAcoBsEN o s,ELBERLING B,et al. Microbial oxidation of pyrite coupled to nitratereduction in anoXic groundwater sediment[J].Environmental Science&Technology,2009,43(13):4851一4857.
[10] sHAo M,zHANG T,FANG H H.sulfur—driven autotrophic denitrifica村on:Diversity biochemistry,and engi—nee ring applications[J].Applied Microbiology and Biotechnology,2010,88(5):1027—1042.
[11] DAPENA—MoRA A,FERNANDEz I,cAMPos J L,et al. Evaluation of activity and inhibition effects onAnammox process by batch te8ts based on the nitrogen gas production[J].Enzyme and Microbial Tbchnology,2007,40(4):859_865。
[12] 鄧旭亮,王爱杰,荣丽丽,等.硫自养反硝化技术研究现状与发展趋势[J].工业水处理,2008(3):13—16.
[13] YANG x,HuANG s,wu Q Nitrate reduction coupled with microbial oXidation of sulflde in river sediment[J].Journal of Soils and Sediments,2012,12(9):1435—1444.
[14] HAAIJER s c M,LAMERs L P M,sMoLDERs A J P,et al.Iron sulfide and pyrite as potential electrondonors for microbial nitrate reduction in freshwater wetlands[J].Geomicrobiology Journal,2007,24(5):391—401
[15] HAYAKAwA A,HATAKEYAMA M,AsANo R,et al Nitrate reduction coupled with pyrite oXidation in the8urface sediment8 0f a sulfide—rich ecosystem[J].Journal of Geophysical Research Biogeosciences,2013,118(2):639—649.
[16] 张梦绯生态修复技术在治理城市黑臭河流的应用[J]污染防治术,2017,30(5):59—61
[17] 李文超,王琦,石寒松,等,城市黑臭河道中硫、铁自养反硝化微生物研究[J].广东化工,2017,44(4):77—78.
[18] 程庆霖,何岩,黄民生,等.城市黑臭河道治理方法的研究进展[J].上海化工,2011,36(2):25—31
[19] 李艳梅硫自养反硝化细菌脱氮除硫性能研究[D].辽宁大连:大连理工大学,2012
[20] sAss H,RAMAMooRTHY s,YARwooD c,et al Desulfouibrio idahonensis sp.nov.,,sulfate—reducingbacteria i80lated from a metal(loid)一contaminated freshwater sediment[J] International Journal of Sy8tematicand Evolutionary Microbiology,2009,59(9):2208一2214.
[21] HALLER L,ToNoLLA M,zoPFI J,et al.comp08ition of bacterial and archaeal communitie8 in freshwatersediments with different contamination levels(Lake Geneva,Switzerland)[J].Water Re8earch,2011,45(3):1213—1228
[22] MEYER B,KuEVER J.Molecular analysis of the distribution and phylogeny of di8similatory aden08ine一5一phosphosulfate reducta8e—encoding gene8(aprBA)among sulfur—oXidizing prokaryotes[J].Microbiology,2007,153f10):3478—3498
[23]MEYER B, KUEVER J. Molecular Analysis of the diversity of sulfate-reducing and sulfur-oxidizing prokaryotesin the environment, using aprA as functional marker gene [J]. Applied and Environmental Microbiology, 2007,73(23): 7664-7679.
[24] MEYER B, KUEVER J. Phylogeny of the alpha and ~oeta subunits of the dissimilatory adenosine-5'-phosphosulfate (APS) reductase from sulfate-reducing prokaryotes - origin and evolution of the dissimilatorysulfate-reduction pathway [J]. Microbiology, 2007, 153(7): 2026-2044.
[25] PHILIPPOT L. Denitrifying genes in bacterial and Archaeal genomes [J]. B Biochimica et Biophysica Acta,2002, 1577(3): 355-376.
[26] 孫韶玲.水体黑臭演过挥发性硫化物的产生机制初步研究 [D].山东烟台:中国科学院烟台海岸带F研究所, 2017.
[27]YE W J. LIU X L, LIN S Q, et al. The vertical distribution of bacterial andarchaeal communities in the waterand sediment of LakeTaihu [J]. FEMS Microbiology Ecology, 2009, 70: 263-276.
[28]PURCELL A M, MIKUCKI J A, ACHBERGER A M, et al. Microbial sulfur transformations in sediments fromSubglacial Lake Whillans [Jl Frontiers in Microbiology, 2014, 5(594): 1-16.
[29] CARDOSO R B, SIERRA-ALVAREZ R, ROWLETTE P, et al. Sulfide oxidation under chemolithoautotrophicdenitrifying conditions [J]. Biotechnology and Bioengineering, 2006, 95(6): 1148-1157.
[30] FLANAGAN D A. GREGORY L G, CARTER J P, et al. Detection of genes for periplasmic nitrate reductasein nitrate respiring bacteria and in community DNA [Jl FEMS Microbiology Letter, 1999, 177(2): 263-270.
[31] ANGELONI N L, JANKOWSKI K J, TUCHMAN N C, et al. Effects of an invasive cattail species (Typha xglauca) on sediment nitrogen and microbial community composition in a freshwater wetland [Jl FEMS Micro-biology Letters, 2006, 263(1): 86-92.
[32]BRAKER G, TIEDJE J M. Nitric oxide reductase (norB) genes from pure cultures and environmental samples[J]. Applied and Environmental Microbiology, 2003, 69(6): 3476-3483.
[33] ANGELONI N L, JANKOWSKI K J, TUCHMAN N C, et al. Effects of an invasive cattail species (Typha xglauca) on sediment nitrogen and microbial community composition in a freshwater wetland [Jl FEMS Micro-biology Letters, 2006, 263(1): 86-92.
[34]BIDERRE-PETIT C, BOUCHER D, KUEVER J, et al. Identification of sulfur-cycle prokaryotes in a low-sulfatelake (Lake Pavin) using aprA and 16S rRNA gene markers [J]. Microbial Ecology, 2011, 61(2): 313-327.
[35]WATANABE T, KOJIMA H, FUKUI M. Identity of major sulfur-cycle prokaryotes in freshwater lake ecosystemsrevealed by a comprehensive phylogenetic study of the dissimilatory adenylylsulfate reductase [J]. ScientificReports, 2016, 6(1): 1-9.
[36] WATANABE T, KOJIMA H, TAKANO Y, et al. Diversity of sulfur-cycle prokaryotes in freshwater lake sed-iments investigated using aprA as the functional marker gene [J]. Systematic and Applied Microbiology, 2013,36(6): 436-443.
[37] KOJIMA H. FUKUI M. Sulfuricella denitrificans gen. nov., sp. nov., a sulfur-oxidizing autotroph isolatedfrom a freshwater lake [J]. I International Journal of Systematic and Evolutionary Microbiology, 2010, 60(12):2862-2866.
[38] SHAO M. ZHANG T. FANG H H. Sulfur-driven autotrophic denitrification: diversity, biochemistry, and engi-neering applications [Jl Applied Microbiology and Biotechnology, 2010, 88(5): 1027-1042.
[39] KOJIMA H. WATANABE T. IWATA T, et al. Identification of major planktonic sulfur oxidizers in stratifiedfreshwater Lake [J]. Plos One, 2014, 9(4): 1-7.
[40]BISOGNI J J, DRISCOLL C T. Denitrification Using Thiosulfate and Sulfide [J]. J Env Eng Div, 1997, 103(4):593-604
[41] RIOS-DEL TORO E E, CERVANTES F J. Coupling between anammox and autotrophic denitrification forsimultaneous removal of ammonium and sulfide by enriched marine sediments [J] . Biodegradation, 2016, 27(2/3):107-118.
[42]REYES-AVILLA J. RAZO-FLORE E, GOMEZ J. Simultaneous biological removal of nitrogen, carbon and sulfurby denitrification [J]. Water Research, 2004, 38(14/15): 3313-3321.
[43] 王愛杰,杜大仲,任南琪.脱氮硫杆菌在废水脱硫、脱氮处理工艺中的应用 [J]哈尔滨i业大学学报, 2004, 36(4): 423-425.
[44]ZHANG S Y, ZHOU Q H, XU D, et al. Effects of sediment dredging on water quality and zooplankton communitystructure in a shallow of eutrophic lake [J]. Journal of Environmental Sciences, 2010, 22(2): 218-224.2015: 8-14.
