日本鳗鲡 II型干扰素基因(IFN-γ和 IFN-γrel)的原核表达与纯化研究
2019-10-26黄文树徐继松翟少伟张婉婷刘海姿
李 想,黄文树,黄 贝,徐继松,翟少伟,梁 英,张婉婷,刘海姿
(1.集美大学水产学院,福建厦门 361021;2.福建省特种水产配合饲料重点实验室,福建福清 350308;3.鳗鲡现代产业技术教育部工程研究中心,福建厦门 361021)
Ⅱ型干扰素(interferon,IFN),即 IFN-γ,是一类二聚化的可溶性细胞因子,主要由T淋巴细胞和NK细胞分泌,具有抗病毒和免疫调节作用。IFN-γ还是一种多效能的细胞因子,能促使巨噬细胞活化、增加抗原递呈作用、激活先天性免疫系统以及调控细胞增殖和凋亡[1]。
在哺乳动物中,Ⅱ型干扰素只有一种类型,即 IFN-γ,由一个基因编码[1]。与哺乳动物不同的是,鱼类的Ⅱ型干扰素存在2个IFN-γ基因,即IFN-γ1和 IFN-γ2,与哺乳动物同源的 IFN-γ命名为IFN-γ2,而IFN-γ1与高等脊椎动物同源性很低,所以又被命名为干扰素相关因子(IFN-γ related molecule,IFN-γrel)[2]。IFN-γrel首次在斑马鱼 (Danio rerio)和绿河豚 (Tetraodon nigirovirdis)中 被 发 现[3]。随 后 在 大 西 洋 鲑(Salmo salar)[4]、虹鳟(Oncorhynchus mykiss)[5]、斑点叉尾鮰(Ictalurus punctatus)[6]、鲤(Cyprinus carpio)[7]、金 鱼 (Carassius auratus)[8]、日 本 鲫(Carassius auratus langsdorfii)[9]、 草 鱼(Ctenopharyngodon idella)[10]以 及 日 本 鳗 鲡(Anguilla japonica)[11]等鱼类中均鉴定出两个IFN-γ基因。与IFN-γ基因结构不同的是 IFN-γrel基因的C端缺乏核定位信号(NLS)结构域,这使其丧失了诱导趋化因子的能力[4]。在免疫原刺激下,斑马鱼IFN-γ和IFN-γrel基因在各组织中的表达水平有所不同,说明其功能可能存在差异[7,12]。有研究显示硬骨鱼类的两个 IFN-γ基因分别有其特异性的受体。在斑马鱼及金鱼中发现 了 2个 IFN-γR1基 因,即 IFN-γR1-1/CRFB17和 IFN-γR1-2/CRFB13,它们可以分别和不同的Ⅱ型干扰素配体结合。金鱼的IFN-γR1-1特异性结合 IFN-γrel,IFN-γR1-2特异性结合IFN-γ[13],说明 IFN-γrel的细胞信号途径可能不同于 IFN-γ。已有的这些研究表明,IFN-γ和IFN-γrel的结构、功能以及信号转导存在差异。
有研究表明,鱼类干扰素是通过类似哺乳动物的JAK-STAT信号通路诱导ISGs的表达从而建立宿主抗病毒防御体系[14]。因此,鱼类的Ⅱ型干扰素也能够参与机体的抗病毒免疫。这些发现为探讨鱼类Ⅱ型干扰素的功能奠定了一定的基础。为了进一步了解鱼类Ⅱ型干扰素的功能特性,还需要在基因水平研究的基础上完善蛋白质水平的研究。但是目前对于鱼类干扰素的应用研究并不多见,且主要集中于I型干扰素,对鱼类IFN-γ的研究较少,特别是对鱼类中特有的Ⅱ型干扰素IFN-γrel的应用研究几乎没有,导致对鱼类两种Ⅱ型干扰素基因的认识非常有限。
日本鳗鲡(Anguilla japonica)是我国重要的名优养殖水产品种类,随着鳗鲡养殖业的发展,各种病害也接踵而来。目前,对鳗鲡免疫系统的作用机制研究较少,对鳗鲡免疫相关基因也所知甚少。之前,笔者首次在日本鳗鲡中发现2个Ⅱ型干扰素基因的存在,本研究在此基础上建立了日本鳗鲡IFN-γ和IFN-γrel基因的原核表达系统,旨在进一步对日本鳗鲡干扰素系统的功能及调控机制进行研究,也为深入研究低等脊椎动物Ⅱ型干扰素的生物学功能奠定基础。
1 材料与方法
1.1 材料与试剂
菌株与质粒包括:pMD19-T载体购自TaKaRa公司(日本);pQE30载体、pET32载体、Escherichia coli DH5α、Escherichia coli M15以及Escherichia coli BL21(DE3)均由本实验室保存。
主要试剂及耗材包括:Green Master Mix购自Promega公司(美国);Gel Extraction Kit试剂盒、Plasmid Mini Kit I(200)试剂盒购自OMEGA公司(美国);Sac I、BamH I和HindⅢ限制性内切酶、T4 DNA Ligase、DL 2000 Marker、DL 5000 Marker以及 Premixed Protein Marker(Low)购自 TaKaRa公司(日本);预染蛋白 Marker和 HisPurTM Ni-NTA Spin Columns购自Thermo公司(美国);histag Mouse Mcab和Goat anti-Mouse IgG-HRP购自Proteintech公司(美国);PVDF膜购自Bio-rad公司(美国);Ecl曝光液购自advansta公司(美国);Temed、吐温 20以及甲叉双丙烯酰胺购自GENERAY公司(中国);过硫酸铵购自aladdin公司(中国);丙烯酰胺购自biosharp公司(中国);考马斯亮蓝G-250购自MP公司(美国);咪唑购自MACKLIN公司(中国);透析袋购自 Solarbio公司(中国);胰蛋白胨、酵母粉、琼脂粉、Trisbase、SDS、甲醇、无水乙醇、异丙醇、冰醋酸、氯化钠、磷酸二氢钠、磷酸氢二钠以及氯化钾等其它常用试剂均为国药分析纯。
1.2 原核表达载体的构建
根据已获得的日本鳗鲡IFN-γ和IFN-γrel基因的cDNA序列以及pQE30和pET32a多克隆位点,利用生物信息学软件Primer Premier 6.0分别设计扩增IFN-γ的引物AJIFNg-F/AJIFNg-R和扩增 IFN-γrel的引物 AJIFNgrel-F/AJIFNgrel-R,并分别在上游及下游引物中引入酶切位点。AJIFNg-F:CGAGCTCGACACCATATCTCAAAAAA T(下划线部分为 Sac I酶切位点),AJIFNg-R:CCCAAGCTTGACCCTGCCTC(下划线部分为HindⅢ酶切位点);AJIFNgrel-F:CGGATCCAACTCCCC GCTCGTCCTGG(下划线部分为 BamH I酶切位点),AJIFNgrel-R:CAAGCTTGTCGGACAGTGAAT CAGTCAGCC(下划线部分为HindⅢ酶切位点)。
分别以日本鳗鲡IFN-γ和IFN-γrel的重组质粒 pMD19-T-IFN-γ和 pMD19-T-IFN-γrel为模板扩 增,以 AJIFNg-F/AJIFNg-R 和 AJIFNgrel-F/AJIFNgrel-R为上下游引物。IFN-γ扩增条件为:94℃预变性5 min;94℃变性30 s,60℃退火30 s,72℃延伸 30 s,35个循环;72℃延伸 10 min。IFN-γrel扩增程序为:94℃预变性5 min;94℃变性30 s,58℃退火 30 s,72℃延伸 30 s,35个循环;72℃延伸10 min。产物经1%琼脂糖凝胶电泳检测后进行胶回收。将IFN-γPCR扩增产物和表达载体pQE30分别于37℃进行SacⅠ和HindⅢ水浴双酶切,IFN-γrel PCR产物和表达载体pET32a分别于37℃水浴进行BamHⅠ和HindⅢ双酶切,酶切产物用T4 DNA连接酶连接过夜。连接产物转化至E.coli DH5α,利用氨苄青霉素和卡那霉素双抗性平板筛选IFN-γ重组质粒阳性克隆,利用氨苄青霉素抗性平板筛选IFN-γrel重组质粒阳性克隆。IFN-γ及 IFN-γrel重组质粒阳性克隆分别用pQE30通用引物和pET32a通用引物经PCR鉴定后送通用生物技术有限公司测序,其中pQE30与pET32a通用引物序列分别为pQE30-F:TGAGCGGATAACAATTTC AC, pQE30-R: GTTCTGAGGTCATTACTGG;pET32a-F:TAATACGACTCACTATAGG,pET32a-R:TGCTAGTTATTGCTCAGCGG。测序正确的阳性重组质粒分别命名为pQE30-IFN-γ和pET32a-IFN-γrel。
1.3 原核表达载体诱导表达条件的优化与表达形式的鉴定
将重组质粒 pQE30-IFN-γ转化至 E.coli M15,重组质粒 pET32a-IFN-γrel转化至 E.coli BL21(DE3),37℃培养过夜,挑取重组质粒pQE30-IFN-γ阳性菌落转接至40 mL含氨苄青霉素和卡那霉素的LB培养基中,挑取重组质粒pET32a-IFN-γrel阳性菌落转接至40 mL含氨苄青霉素的LB培养基中,37℃、180 r·min-1震荡培养过夜。当菌液 OD200达到0.6左右时,于37℃诱导温度下,测定以不同诱导剂浓度(0.2、0.5、1.0、2.0 mmol·L-1IPTG)、不同诱导时间(2、4、6 h)诱导 IFN-γ蛋白表达工程菌表达重组蛋白的表达量。测定以1.0 mmol·L-1IPTG分别诱导 IFN-γrel蛋白表达工程菌2、4、6 h时表达重组蛋白的表达量。收集菌液并用PBS重悬,SDS-PAGE电泳鉴定重组蛋白表达量。同时设置空载体即未加IPTG诱导的重组质粒作为对照。
在最佳诱导条件下收集表达菌液,10 000×g离心5 min,弃上清液,用30 mL PBS重悬菌体,于4℃下进行超声波裂解(工作6 s间隔9 s),13 000×g离心5 min。分别收集上清液和沉淀,取少量样品进行SDS-PAGE电泳,鉴定重组蛋白的表达形式。
1.4 IFN-γrel目的蛋白的变复性处理
选择最佳条件诱导IFN-γrel表达工程菌,收集沉淀;用事先预冷的PBS重悬沉淀,加入适当的蛋白酶抑制剂PMSF,4℃超声波裂解(工作6 s间隔9 s),13 000×g离心 5 min,收集沉淀;加入10 mL含4 mol·L-1尿素的变性液洗涤沉淀2次,再加入10 mL含8 mol·L-1尿素的蛋白变性液,4℃震荡过夜,彻底溶解包涵体;次日将收集的蛋白变性液放入处理好的透析袋中,移至含4 mol·L-1尿素的透析液中,期间更换透析液,使变性液中8 mol·L-1的尿素逐步降低至4、2、1 mol·L-1,最后用pH8.0的PBS透析2次,将溶液中的尿素全部去除完成复性。
1.5 目的蛋白的分离纯化及Western blot鉴定
将IFN-γ表达工程菌上清液和复性好的IFN-γrel重组蛋白溶液分别过 HisPurTMNi-NTA Spin Columns纯化目的蛋白。IFN-γ目的蛋白纯化步骤:首先加入 6 mL平衡缓冲液(20 mmol·L-1磷酸钠、500 mmol·L-1氯化钠、10 mmol·L-1咪唑各 2 mL,pH8.0)平衡柱子,重复操作1次;向柱子内加入IFN-γ表达工程菌上清液,4℃结合30 min,使样品在重力的作用下缓慢流出,收集流出液;将收集的流出液再加入柱子内,重复过柱1次;加入6 mL洗涤缓冲液(20 mmol·L-1磷酸钠、500 mmol·L-1氯化钠、25 mmol·L-1咪唑各 2 mL,pH8.0)洗涤柱子,重复洗柱 2次;而后加入 3 mL洗脱缓冲液(20 mmol·L-1磷酸钠、500 mmol·L-1氯化钠、250 mmol·L-1咪唑各 1 mL,pH 8.0),收集洗脱液,并进行SDS-PAGE凝胶电泳检测。纯化IFNγ-rel目的蛋白使用的缓冲液为将上述缓冲液中的氯化钠浓度置换为300 mmol·L-1,其余试剂及其浓度不改变,纯化步骤同IFN-γ。
将纯化的目的蛋白进行SDS-PAGE电泳检测。电泳后,以300 mA 70 min转印至PVDF膜上,然后将PVDF膜置于封闭液中浸泡1 h,清洗后加入 his-tag Mouse Mcab(稀释浓度1∶1 000),4℃孵育过夜,加入Goat anti-Mouse IgG-HRP(稀释浓度1∶10 000)室温孵育1 h。各步骤之间用TBST(Washing Buffer)清洗 3次,每次各 20 min。最后将PVDF膜放入避光处加Ecl显色液,2 min后置凝胶成像系统观察结果并拍照。
2 结果与分析
2.1 原核表达载体的鉴定
以重组质粒 pMD19-T-IFN-γ作为模板,以AJIFNg-F和AJIFNg-R为上下游引物,PCR扩增IFN-γ基因成熟肽编码序列,结果如图1-A所示,在约464 bp处有一条特异性条带,与预期大小基本一致。以重组质粒 pMD19-T-IFN-γrel作为模板,以AJIFNgrel-F和AJIFNgrel-R为上下游引物,PCR扩增IFN-γrel基因成熟肽编码序列,结果如图1-B所示,在约440 bp处有一条特异性条带,与预期大小符合。表明已分别扩增出IFN-γ和IFN-γrel基因的成熟肽编码序列。
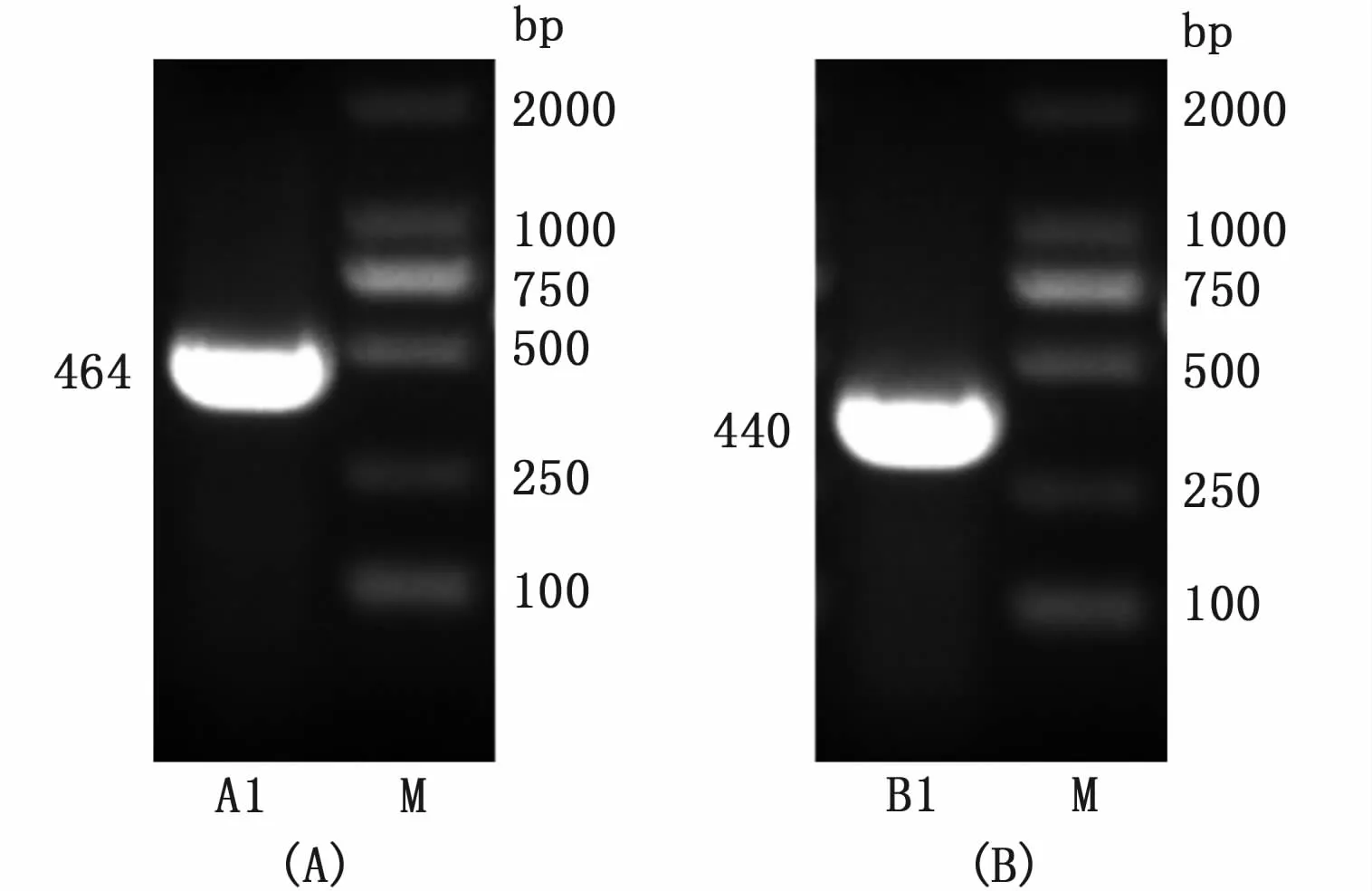
图1 IFN-γ和IFN-γrel基因PCR扩增结果Fig.1 PCR amplification results of IFN-γand IFN-γrel
挑取单克隆做阳性克隆鉴定,结果如图2所示,回收片段比成熟肽编码序列多100 bp左右,初步判断IFN-γ和IFNγ-rel成熟肽编码序列分别与载体连接成功。将重组质粒测序,结果表明重组表达载体 pQE30-IFN-γ和 pET32a-IFN-γrel构建成功。
2.2 重组表达蛋白诱导表达条件的优化
将重组表达载体 pQE30-IFN-γ转化至 E.coli M15,经IPTG诱导表达后进行SDS-PAGE分析。结果如图3所示,与pQE30空载体相比,重组质粒pQE30-IFN-γ在M15中有明显的诱导表达蛋白条带,大小在21 kD左右,与预计算的理论分子量(20.5 kD)相似,初步判断已诱导表达出IFN-γ目的蛋白。分别测定4个IPTG诱导浓度以及不同诱导时间对重组蛋白表达量的影响,发现pQE30-IFN-γ在IPTG浓度为1.0 mmol·L-1时表达量最高,并且表达量随着诱导时间的增加而增加,即1.0 mmol·L-1IPTG诱导6 h时表达量最高。

图2 p QE30-IFN-γ和 pET32a-IFN-γrel重组载体的阳性克隆鉴定Fig.2 Identification of positive clones of pQE30-IFN-γand p ET32a-IFN-γrel recombinant vectors
将重组表达载体pET32a-IFN-γrel转化至E.coli BL21,经1 mmol·L-1IPTG诱导表达后进行SDS-PAGE分析。结果如图4所示,与pET32a空载体相比,重组质粒pET32a-IFN-γrel在BL21中有明显的诱导表达蛋白条带,大小在31 kD左右,与预计算的理论分子量(34 kD)相似,故初步判断已诱导表达出IFN-γrel目的蛋白。与未诱导的IFN-γrel蛋白表达工程菌相比,诱导0~6 h内,随着诱导时间的延长,蛋白表达量越高,即诱导6 h为最优时间。
2.3 重组蛋白表达形式的鉴定
取表达菌体超声波破碎裂解后,分离上清液和沉淀进行SDS-PAGE电泳分析。结果显示,重组表达pQE30-IFN-γ蛋白大部分以可溶性蛋白的形式存在于破碎菌体的上清液中(图5-A),而重组表达pET32a-IFN-γrel蛋白大部分以包涵体蛋白的形式存在于破碎菌体的沉淀中(图5-B)。
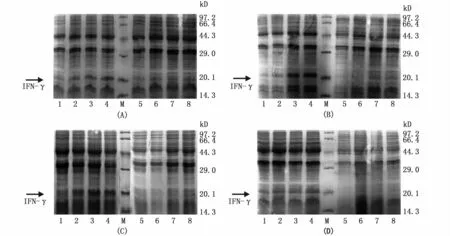
图3 不同诱导条件对pQE30-IFN-γ在E.coli M15中表达的影响Fig.3 Influence of different induction conditions on expression of pQE30-IFN-γin E.coli M15
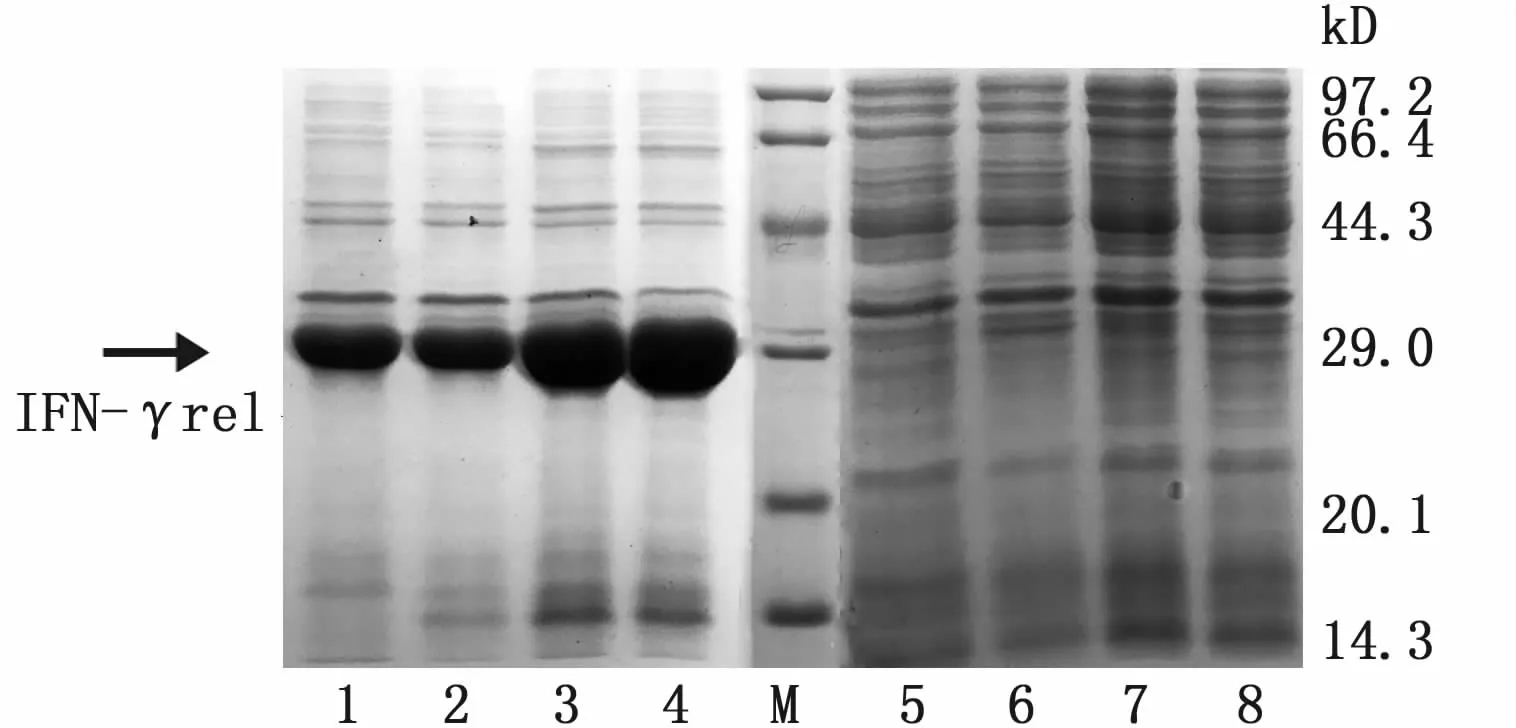
图4 不同诱导时间对pET32a-IFN-γrel在E.coli BL21中表达的影响Fig.4 Influence of different induction time on expression of pET32a-IFN-γrel in E.coli BL21
2.4 重组蛋白的纯化和Western blot分析结果
采用镍离子亲和层析柱纯化目的蛋白。收集洗脱液进行SDS-PAGE凝胶电泳检查,结果显示,在IFN-γ表达工程菌破碎后的上清液中纯化得到IFN-γ融合蛋白,大小约为21 kD(图6-A),在复性好的IFN-γrel表达工程菌破碎后的沉淀中纯化得到 IFNγ-rel融合蛋白,大小约为31 kD(图6-B)。两者均为单一条带,说明经过纯化步骤获得的重组蛋白纯度较高。
将纯化得到的目的蛋白进行Western blot鉴定,结果表明,与空载体相比,表达产物泳道显示为阳性条带,即带有His标签的重组蛋白IFN-γ和 IFN-γrel得到表达(图7)。
3 讨论
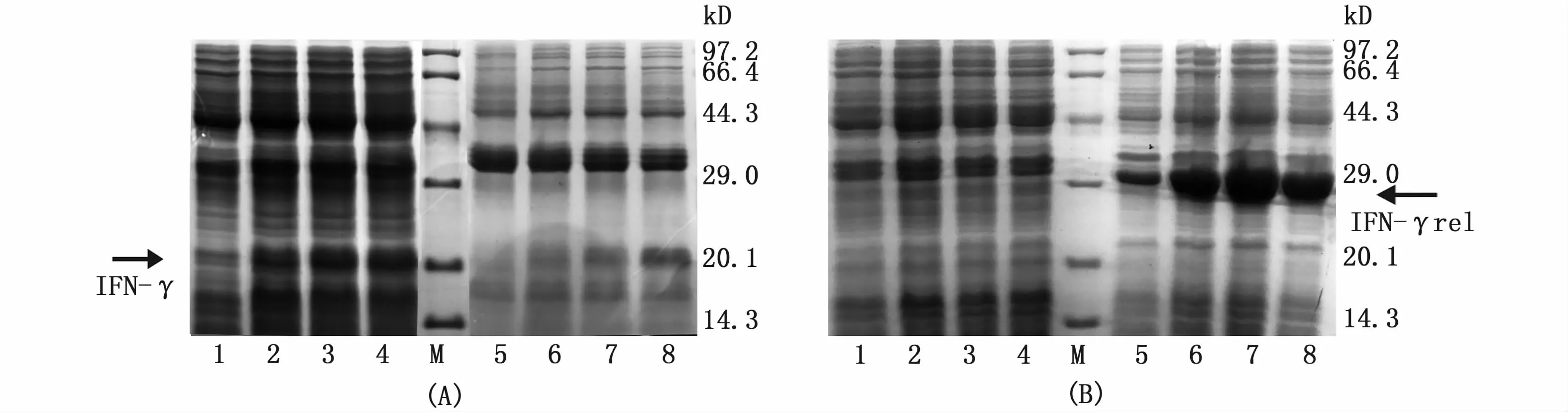
图5 pQE30-IFN-γ和pET32a-IFN-γrel重组蛋白表达形式鉴定Fig.5 Identification of protein expression patterns of pQE30-IFN-γ and pET32a-IFN-γrel recombinant proteins
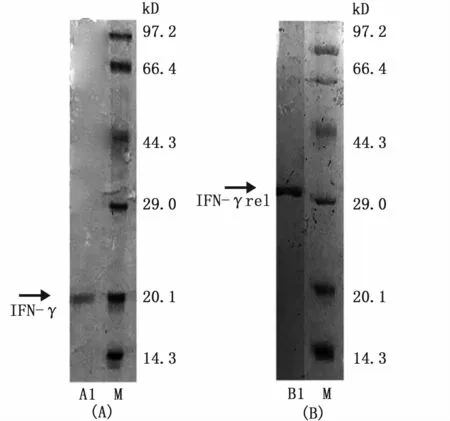
图6 纯化的 IFN-γ和 IFN-γrel重组蛋白SDS-PAGE分析Fig.6 SDS-PAGE analysis of purified IFN-γ and IFN-γrel recombinant proteins

图7 IFN-γ和 IFN-γrel重组蛋白Western blotting鉴定结果Fig.7 Identification results of IFN-γand IFN-γrel recombinant proteins by Western blotting
目前,哺乳动物和禽类的重组IFN-γ蛋白已得到广泛研究和利用。崔然[15]构建了猪 IFN-γ基因的原核表达载体,发现重组蛋白具有抵抗猪繁殖与呼吸综合征病毒和水泡性口炎病毒的能力。王立红等[16]成功利用E.coli M15感受态细胞表达出具有生物活性的鸡IFN-γ。然而,有关鱼类基因工程干扰素制剂的研究并不多见。ZOU等[17]将重组虹鳟IFN-γ蛋白对巨噬细胞进行刺激,重组蛋白能刺激IFN-γ信号通路下游基因的表达。GRAYFER等[8]发现金鱼重组IFN-γ蛋白刺激后的巨噬细胞能增强吞噬作用。此外,重组IFN-γ蛋白还能刺激一些促炎症因子(如:TNFα-1、TNFα-2等)的表达。IFN-γrel是硬骨鱼特有的基因,对于此基因的报道很少,而对其应用研究几乎没有。目前仅见鳜(Siniperca chuatsi)IFN-γrel重组蛋白应用的报道,其能够诱导免疫相关基因Mx、IRF1、STAT1等的表达上调,可作为鱼类免疫增强剂或者免疫佐剂[18]。为了深入研究干扰素基因的功能,在基因水平研究的基础上通过基因重组技术进一步完善蛋白质水平的研究是有必要的。
融合表达载体是目前原核表达系统中常用的基因工程载体。由于pET表达载体系统含有的his-tag是一个广泛应用于亲和层析的标签,并根据已有的关于鱼类IFN-γ重组蛋白表达的研究报道,在选择载体时,笔者首先使用高效融合表达载体pET32a来构建2个Ⅱ型干扰素重组质粒。将IFN-γrel成熟肽编码序列克隆至原核表达载体 pET32a上,成功构建了重组表达载体pET32a-IFN-γrel,并在 E.coli BL21中得到表达,说明所选pET32a载体和IFN-γrel宿主表达菌是合适的。但在实验中,重组载体pET32a-IFN-γ表达的蛋白与预测不符,未能正确表达,后尝试优化培养条件,经多次实验证明重组pET32a质粒无法表达出IFN-γ重组蛋白,所以改为选用带有6个his标签的pQE30质粒作为表达载体,以E.coli M15作为宿主菌,最终成功表达出IFN-γ蛋白。
对动物干扰素进行原核诱导表达时发现,除了基因本身的序列影响表达量外,诱导表达条件(如诱导时间、诱导温度和诱导剂浓度等)同样重要[19]。因此,有必要对原核表达条件进行摸索。蔡梅红等[20]对鸡IFN-γ阳性大肠杆菌宿主菌的不同诱导时间进行了对比研究,确定了4 h为最佳的诱导时间。马普等[21]对重组载体的最佳表达条件进行了探索,最终确定了重组红鳍东方鲀(Takifugu rubripes)IFN-γ蛋白在 IPTG为 1.0 mmol·L-1、37℃条件下诱导6 h时达到最大表达量。在本研究中发现通过调整不同的IPTG诱导浓度和诱导时间,日本鳗鲡IFN-γ和IFN-γrel在1.0 mmol·L-1IPTG诱导6 h时表达量最高。
外源基因表达量低以及易形成包涵体是外源基因在大肠杆菌中表达时常见的难题。据估计,有15%~20%是可溶性蛋白,大约20%~40%是包涵体蛋白,剩余的可能在表达过程中有降解或者不表达。获得可溶性蛋白可以避免蛋白质的变性和复性,便于今后对蛋白活性的研究。因此,在大肠杆菌中进行外源基因表达时首先要考虑改善和优化影响外源基因可溶性表达的因素,如表达菌株、表达载体、培养基的组成及培养条件等[22]。有研究表明,在高水平表达时,新生肽链聚集速率一旦超过蛋白折叠的速率就会导致包涵体的形成,而表达量低则利于产生可溶性表达[23]。在本研究中,笔者发现IFN-γ蛋白呈现可溶性表达,但蛋白表达量却不高。而IFN-γrel蛋白则以包涵体形式表达,且获得了高效表达,这也是包涵体存在的一个优势,即高效的表达易于从宿主细胞中纯化[24]。
HisPurTMNi-NTA Spin Columns是以预挂Ni2+电荷的形式提供的。蛋白和金属离子之间的结合强度受几种因素影响,包括长度、位置、亲和标记在蛋白中的暴露程度、所用离子的类型以及缓冲液的pH等。在纯化IFN-γ蛋白的过程中,根据蛋白的等电点,笔者最初选择pH值为7.4的平衡、洗涤及洗脱缓冲液,但在收集的平衡和洗涤穿透液中检测到大量目的蛋白,说明目的蛋白和Ni2+未能有效结合,纯化效果不理想。所以笔者通过改变缓冲液的pH值(比较了7.4、8.0和8.5等3个pH值)来优化纯化效果。结果发现,在缓冲液pH值为8.0的条件下,获得了较高纯度和较高产量的目的蛋白。咪唑的浓度在重组蛋白纯化过程中起着十分重要的作用[25]。低浓度咪唑能够减少宿主细胞中其它蛋白质与Ni2+的非特异性结合,所以,通常在平衡和洗涤缓冲液中采用低浓度的咪唑。笔者在实验中选择了10~25 mmol·L-1的咪唑配制平衡洗涤缓冲溶液。高浓度咪唑则有利于洗脱目的蛋白。本实验采用250 mmol·L-1咪唑进行洗脱达到了较好的纯化效果。而参照纯化IFN-γ蛋白的条件对IFN-γrel蛋白进行纯化时发现在洗脱液中仅收集到非常少的目的蛋白,因此改变了缓冲液中氯化钠的浓度,将氯化钠浓度从500 mmol·L-1降低至300 mmol·L-1,结果回收到的目的蛋白量明显增加。这是因为洗脱液的pH值和盐度是影响蛋白质和金属螯合配体间作用力的主要因素[26]。在蛋白纯化过程中加入中性盐(如氯化钠)能影响蛋白质的溶解度。一般蛋白质溶解度在一定范围内随着盐度的升高而增加,当盐度继续升高时,蛋白质的溶解度出现不同程度下降并先后析出,而且不同的蛋白质纯化时其洗脱体系存在不同的合适盐度条件。故推测IFN-γrel蛋白在500 mmol·L-1氯化钠的缓冲液体系中发生了一定的盐析作用,导致蛋白的损耗。
采用His标签抗体对诱导表达的蛋白进行的Western blot检测,结果表明重组载体按照正确的翻译框架翻译并成功表达。
